第15章 蛋白质翻译
合集下载
第15章蛋白质翻译ppt课件

原核的mRNA 是多顺反子;真核的mRNA 是单顺反子。
1.2 转运RNA (tRNA)
原核tRNA有30-40种,真核有50-60种,含70-90个核苷酸, 并 有多种稀有碱基。
tRNA是最小的RNA, 占细胞总RNA 的15%左右,其功能 是搬运氨基酸和解读密码子。
tRNA 具有“四环一臂”和“三叶草” 形的典型结构。
(3)氨基酸侧链的修饰 氨基酸侧链的修饰包括羟化、羧化、甲基化 及二硫链的形成等。
(4)糖基化修饰 糖蛋白是细胞蛋白质组成的重要成分。它是在翻译 后的肽链上以共 价键与单糖或寡聚糖连接而成。糖基化是在酶催化 下进行的。
分泌型蛋白质在翻译过程中通过信号肽协助转入内质网的机制
信号肽(signal peptide)是在新生的多肽链中,可被细胞识别系统识别的 特征性氨基酸序列,在蛋白质翻译过程中或翻译后的定位发挥引导的作用。
第15章 蛋白质翻译
Protein Biosynthesis (Translation)
本章主要内容
翻译系统 蛋白质生物合成的过程 多肽链翻译后的修饰 蛋白质的到位
Байду номын сангаас
原核
真核
基因在原核和真核细胞中最终得到表达
1 蛋白质的翻译系统
原料氨基酸,20种 mRNA是合成蛋白质的“蓝图(或模板)” tRNA是原料氨基酸的“搬运工” rRNA与多种蛋白质结合成核糖体,作为合成多肽链的 “装配机(操作台)”
多个核糖体结合在同一条mRNA上,由5’——3’进行翻译,形成多 核糖体(polyribosome),以提高翻译的效率
3 多肽链的修饰和加工
(1)N—端修饰 原核生物修饰时是由肽甲酰基酶除去甲酰基,多数 情况甲 硫氨酸也被氨肽酶除去,真核生物中甲硫氨酸则全部被切除。
1.2 转运RNA (tRNA)
原核tRNA有30-40种,真核有50-60种,含70-90个核苷酸, 并 有多种稀有碱基。
tRNA是最小的RNA, 占细胞总RNA 的15%左右,其功能 是搬运氨基酸和解读密码子。
tRNA 具有“四环一臂”和“三叶草” 形的典型结构。
(3)氨基酸侧链的修饰 氨基酸侧链的修饰包括羟化、羧化、甲基化 及二硫链的形成等。
(4)糖基化修饰 糖蛋白是细胞蛋白质组成的重要成分。它是在翻译 后的肽链上以共 价键与单糖或寡聚糖连接而成。糖基化是在酶催化 下进行的。
分泌型蛋白质在翻译过程中通过信号肽协助转入内质网的机制
信号肽(signal peptide)是在新生的多肽链中,可被细胞识别系统识别的 特征性氨基酸序列,在蛋白质翻译过程中或翻译后的定位发挥引导的作用。
第15章 蛋白质翻译
Protein Biosynthesis (Translation)
本章主要内容
翻译系统 蛋白质生物合成的过程 多肽链翻译后的修饰 蛋白质的到位
Байду номын сангаас
原核
真核
基因在原核和真核细胞中最终得到表达
1 蛋白质的翻译系统
原料氨基酸,20种 mRNA是合成蛋白质的“蓝图(或模板)” tRNA是原料氨基酸的“搬运工” rRNA与多种蛋白质结合成核糖体,作为合成多肽链的 “装配机(操作台)”
多个核糖体结合在同一条mRNA上,由5’——3’进行翻译,形成多 核糖体(polyribosome),以提高翻译的效率
3 多肽链的修饰和加工
(1)N—端修饰 原核生物修饰时是由肽甲酰基酶除去甲酰基,多数 情况甲 硫氨酸也被氨肽酶除去,真核生物中甲硫氨酸则全部被切除。
蛋白质的生物合成翻译3(共51张PPT)

AGGA或GAGG(Shine-Dalgarno顺序)互补。正是由这样的
配对将AUG(或GUG,UUG)密码子带到核糖体的起始位置 上 。 fMet-tRNA 与小亚基上的A位点结合。
2. 70S起始复合物的形成
30S起始复合物一旦完全形成后,IF3即释放出来。50S大亚 基参加进来,并引起GTP水解和释放其它两个起始因子,最后
ATP与GTP提供。
蛋白质的生物合成过程主要包括:合成的起始;肽链的延伸;合 成的终止与多肽链释放。
一、氨基酸的激活与氨酰基-tRNA的合成
1. 氨基酸的激活与氨酰基-tRNA的合成过程
氨基酸不能直接与模板相结合,必须首先与相应的tRNA结合, 形成氨酰基‐tRNA。这一过程就是氨基酸的激活。
将氨基酸接合于tRNA以形成氨酰基‐tRNA的激活反应是在氨酰 基‐tRNA合成酶的催化作用下进行的,需要ATP提供能量 。这个反 应是不可逆转的 。
2. 真核生物mRNA为单顺反子;原核生物mRNA为多顺反子。
3. 原核生物的mRNA的5’-端有SD序列。许多真核mRNA的AUG上游 存在Kozak(CCACC)序列。
4. 真核生物的成熟mRNA需要从细胞核中进入细胞质,才能参与蛋 白质的翻译;而原核生物的转录与翻译偶联在一起。
(二)遗传密码
Together with models in the corresponding orientation. The complete 70S ribosome has an asymmetric construction. The partition between the head and body of the small subunit is aligned with the notch of the large subunit, so that the platform of the small subunit fits into the large subunit. There is a cavity between the subunits which contains some of the important sites.
配对将AUG(或GUG,UUG)密码子带到核糖体的起始位置 上 。 fMet-tRNA 与小亚基上的A位点结合。
2. 70S起始复合物的形成
30S起始复合物一旦完全形成后,IF3即释放出来。50S大亚 基参加进来,并引起GTP水解和释放其它两个起始因子,最后
ATP与GTP提供。
蛋白质的生物合成过程主要包括:合成的起始;肽链的延伸;合 成的终止与多肽链释放。
一、氨基酸的激活与氨酰基-tRNA的合成
1. 氨基酸的激活与氨酰基-tRNA的合成过程
氨基酸不能直接与模板相结合,必须首先与相应的tRNA结合, 形成氨酰基‐tRNA。这一过程就是氨基酸的激活。
将氨基酸接合于tRNA以形成氨酰基‐tRNA的激活反应是在氨酰 基‐tRNA合成酶的催化作用下进行的,需要ATP提供能量 。这个反 应是不可逆转的 。
2. 真核生物mRNA为单顺反子;原核生物mRNA为多顺反子。
3. 原核生物的mRNA的5’-端有SD序列。许多真核mRNA的AUG上游 存在Kozak(CCACC)序列。
4. 真核生物的成熟mRNA需要从细胞核中进入细胞质,才能参与蛋 白质的翻译;而原核生物的转录与翻译偶联在一起。
(二)遗传密码
Together with models in the corresponding orientation. The complete 70S ribosome has an asymmetric construction. The partition between the head and body of the small subunit is aligned with the notch of the large subunit, so that the platform of the small subunit fits into the large subunit. There is a cavity between the subunits which contains some of the important sites.
9. 蛋白质翻译(1)

摇摆的原因(摇摆假说):
一般地,同义密码子的第1、2位是保守的,而第3位 则是可变的,意味着该可变位点的配对具有一定的灵活 性。
tRNA的反密码子在反密码环上呈弧状排列,与密码子 不能保持完全的平行排列;另外,反密码子的第1个核 苷酸位于非双链结构的松弛环内,摇摆的自由度较大, 从而导致密码子的第3位核苷酸和反密码子的第1位核苷 酸之间形成非标准的碱基配对。(反密码子的这个位点 称为摇摆位点) 如果tRNA的摇摆位点是被修饰的碱基,就可能出现更 多的选择配对关系。
上次讲解内容
一、顺式作用元件与反式作用因子(重点) 二、真核生物RNA的转录过程 三、真核生物RNA转录后加工(重点) 1. 5’加帽; 2. 3’加尾; 3. 选择性剪接; 4. RNA编辑 四、RNA编辑
碱基的突变
C变为U
ApoB 基因有 29 个外显子
CAA
第 2153 个密码子编码 Glu 编辑
T-loop(TψC环)
• 这个环中始终含有胸 腺嘧啶-假尿嘧啶-胞嘧 啶的序列。 • 它与核糖体大亚基的 5S rRNA结合,稳定 蛋白质的结构
D-loop (DHU环)
直接与氨基酰tRNA合成酶
结合,使氨基酸连接到 tRNA的受体位点上。
tRNA与氨基酰tRNA合成酶的结合
氨基酸连接到受体位点上的过程:
UAA
3’UTR
AAA
Open reading frame(开放阅读框), ORF (3’非翻译区)
Stop codon(终止密码) UAG UGA UAA
开放阅读框(open reading frame, ORF): mRNA中从起始密码子(AUG)到终止密 码子(UAA、UAG或UGA)的核酸序列, 它可以编码一条完整的多肽链。
分子生物学第十五章 蛋白质的生物合成

8
1.遗传密码种类:
• mRNA 分子有4种碱基:A、G、C、U,可组合成64个密 码子,其中61个分别代表20种不同氨基酸
• 遗传密码共有43=64种, 64: UAA、UAG、UGA 终止密码 61: AUG 起始密码 代表Met 60: 代表19种氨基酸
9
遗传密码表 第一碱基
(5/-端)
第二碱基
43
一、起始阶段
(一)原核生物翻译起始复合物的形成
参与的物质: 核糖体50S和30S大小两类亚基 mRNA 起始作用的fMet-tRNAfMet GTP供能 起始因子(IF1、IF2、IF3)
44
起始过程:
• 核糖体大小亚基解离 • 小亚基与mRNA结合:
16sRNA识别mRNA的SD序列
• fMet-tRNAfMet与mRNA起始密码子AUG结合 • 核糖体大小亚基形成起始复合物
tRNAphe Phe-tRNAphe
• 肽链起始和延长的甲硫氨酰-tRNA:
真核生物:起始: Met-tRNAiMet 延长: Met-tRNAMet
原核生物: 起始:fMet-tRNAfMet 延长:Met-tRNAMet
39
40
第三节 肽链的合成过程
41
整个翻译过程可分为 :
• 翻译的起始 • 翻译的延长 • 翻译的终止
tRNA反密码子 第1位碱基
mRNA密码子 第3位碱基
I
U
G AC
U, C, A A, G U, C U G
17
摆 动 配 对
32 1
U
123
18
• ⑤遗传密码的通用性
从原核生物到人类都共用同一套遗传密码,被称为遗传密码的通用性。 密码的通用性进一步证明各种生物进化自同一祖先。
1.遗传密码种类:
• mRNA 分子有4种碱基:A、G、C、U,可组合成64个密 码子,其中61个分别代表20种不同氨基酸
• 遗传密码共有43=64种, 64: UAA、UAG、UGA 终止密码 61: AUG 起始密码 代表Met 60: 代表19种氨基酸
9
遗传密码表 第一碱基
(5/-端)
第二碱基
43
一、起始阶段
(一)原核生物翻译起始复合物的形成
参与的物质: 核糖体50S和30S大小两类亚基 mRNA 起始作用的fMet-tRNAfMet GTP供能 起始因子(IF1、IF2、IF3)
44
起始过程:
• 核糖体大小亚基解离 • 小亚基与mRNA结合:
16sRNA识别mRNA的SD序列
• fMet-tRNAfMet与mRNA起始密码子AUG结合 • 核糖体大小亚基形成起始复合物
tRNAphe Phe-tRNAphe
• 肽链起始和延长的甲硫氨酰-tRNA:
真核生物:起始: Met-tRNAiMet 延长: Met-tRNAMet
原核生物: 起始:fMet-tRNAfMet 延长:Met-tRNAMet
39
40
第三节 肽链的合成过程
41
整个翻译过程可分为 :
• 翻译的起始 • 翻译的延长 • 翻译的终止
tRNA反密码子 第1位碱基
mRNA密码子 第3位碱基
I
U
G AC
U, C, A A, G U, C U G
17
摆 动 配 对
32 1
U
123
18
• ⑤遗传密码的通用性
从原核生物到人类都共用同一套遗传密码,被称为遗传密码的通用性。 密码的通用性进一步证明各种生物进化自同一祖先。
蛋白质的翻译过程
起始密码
➢肽链合成的起始
❖30s起始复合物形成
1.核糖体亚基的拆离
2.mRNA在小亚基上就位 3.fmet-tRNAfmet的结合
起始序列(SD 序列)
30S小亚基与mRNA识别、结 合
IF1、IF3协助 fmet-tRNAfmet -IF2-GTP 通 过
其反密码与mRNA上的起始密
码
AUG相配对
蛋白质的生物合成-翻译
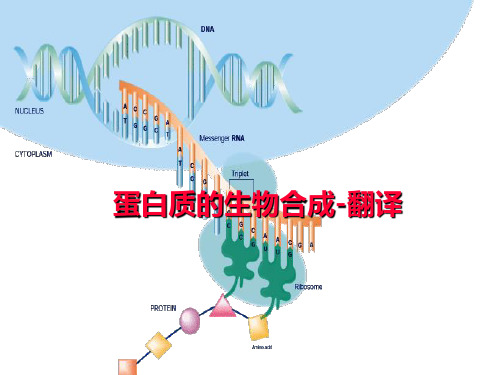
分子生物学的中心法则(central dogma)
复制 DNA
RNA复制
转录
RNA
翻译
蛋白质
逆转录
2
翻译(蛋白质的生物合成)
蛋白质生物合成体系
➢以氨基酸为原料
➢以mRNA为模板 ➢以tRNA为运载工具 ➢以核糖体为合成场所
➢起始、延长、终止各阶段蛋白因子 参与合成后加工成为有活性蛋白质
❖氨基酰tRNA合成 酶
❖催化反应
❖氨基酰tRNA
氨基酰tRNA合成酶
A.A+特异tRNA
氨基酰tRNA
ATP AMP+PPi
氨基酸 பைடு நூலகம் ATP-E 氨基酰-AMP-E + PPi
氨基酰-AMP-E+tRNA 氨基酰tRNA+AMP+E (-COOH) (3’-CCA-OH)
16
2 、氨基酰tRNA合成酶 的高度专一性
➢核糖体与特异蛋白质、mRNA、tRNA的反应 部位
➢新技术 低温电子显微镜技术 中子散射技术
14
第二节 蛋白质合成的过程
原核生物 氨基酸的活化与转运 肽链合成的起始 肽链的延长 “核糖体循环” 肽链合成的终止 蛋白质的加工、修饰
分子生物学-蛋白质的翻译课件

详细描述
核糖体通过识别mRNA上的起始密码子与mRNA结合,形成 翻译起始复合物。这个过程需要消耗能量,以确保核糖体正 确地定位在起始密码子上。
起始复合物的形成
总结词
起始复合物的形成是翻译过程的重要步骤,它涉及到多个蛋白质和RNA分子的相互作 用。
详细描述
起始复合物的形成涉及多个步骤。首先,核糖体与mRNA结合后,需要招募翻译起始 因子,如IF3和IF2。这些因子帮助核糖体正确地定位在起始密码子上,并确保翻译的准 确性。随后,氨酰-tRNA结合到核糖体的A位点上,准备开始多肽链的合成。至此,起
肽链的延长
01
02
03
肽键的形成
氨基酸在加入到肽链中后, 通过肽键的形成相互连接, 形成多肽链。
转肽酶的作用
转肽酶在肽键形成过程中 起催化作用,促进氨基酸 之间的连接。
核糖体的移动
随着肽链的延长,核糖体 沿着mRNA移动,确保下 一个密码子被正确识别和 翻译。
终止密码子的识别
终止密码子的种类
终止密码子有UAA、UAG和UGA三种,它们作为翻译终止的信号 被核糖体识别。
翻译的起始
02
起始密码子
总结词
起始密码子是mRNA上的一个特定 序列,用于标记蛋白质合成的起始位 置。
详细描述
起始密码子是mRNA上的三个连续的 核苷酸,通常为AUG。它不仅标记了 翻译开始的位点,还决定了从这里开 始合成多肽链的方向。
核糖体与mRNA的结合
总结词
核糖体是负责蛋白质合成的细胞器,它通过与mRNA的结合 开始翻译过程。
无意义校正是指当mRNA上的终止密码子提前出现时,核 糖体会提前终止多肽链的合成。这种机制有助于减少多肽 链的错误合成。
分子生物学—蛋白质翻译PPT课件

• 反密码子臂:位于套索中央有三联反密码子。
• D臂:含有二氢尿嘧啶。
各种tRNA均有74-95个核苷酸,其中 有22个核苷酸是恒定不变的。
• 受体臂:链两端碱基序列互补形成 7bp的茎;3’端有未配对的3~4个碱基; 3’端的CCA,最后一个碱基2'和3’烃基 可被氨酰化。
• TψC臂:常由5bp的茎和7nt的环组成。 负责核糖体的识别。
• tRNA的性质是由反密码子而不是它所携带的氨基酸所决定的:
试验:14C-Cys-tRNAcys ---Ni---> 14C-Ala-tRNAcys
4.2.3 tRNA的种类
1. 起始tRNA和延伸tRNA。
• 一类能特异地识别mRNA模板上起始密码子的tRNA叫起始tRNA,其他tRNA统称为 延伸tRNA。 起始tRNA具有独特的结构特征。
• 反密码子臂:常由5bp的茎和7nt的环 组成。
• D臂:含有二氢尿嘧啶。茎的长度常 为4bp。
• 额外臂:4-21nt不等。
tRNA上碱基的修饰
tRNA的稀有碱基非常丰富,约有 70余种。每个tRNA分子至少有2 个稀有碱基,最多有19个。
多数分布在非配对区,尤其反 密码子3‘端附近部位,且大多 为嘌呤核苷酸。
4 生物信息的传递 (下)
基因的遗传信息在转录过程中从 DNA转移到mRNA;再由mRNA将 遗传信息表达为蛋白质中氨基酸顺 序的过程称翻译,即蛋白质的生物 合成过程。
• 蛋白质是生物信息通路上的终产物,一个活细胞在任何发育阶段都需要 数千种不同的蛋白质。因此,活细胞内时刻进行着各种蛋白质的合成、 修饰、运转和降解反应。
用其它2个碱基的共聚物进行类似的实验,也可以推断出其他密码子的碱 基组成,但不能确定密码子中碱基排列。
• D臂:含有二氢尿嘧啶。
各种tRNA均有74-95个核苷酸,其中 有22个核苷酸是恒定不变的。
• 受体臂:链两端碱基序列互补形成 7bp的茎;3’端有未配对的3~4个碱基; 3’端的CCA,最后一个碱基2'和3’烃基 可被氨酰化。
• TψC臂:常由5bp的茎和7nt的环组成。 负责核糖体的识别。
• tRNA的性质是由反密码子而不是它所携带的氨基酸所决定的:
试验:14C-Cys-tRNAcys ---Ni---> 14C-Ala-tRNAcys
4.2.3 tRNA的种类
1. 起始tRNA和延伸tRNA。
• 一类能特异地识别mRNA模板上起始密码子的tRNA叫起始tRNA,其他tRNA统称为 延伸tRNA。 起始tRNA具有独特的结构特征。
• 反密码子臂:常由5bp的茎和7nt的环 组成。
• D臂:含有二氢尿嘧啶。茎的长度常 为4bp。
• 额外臂:4-21nt不等。
tRNA上碱基的修饰
tRNA的稀有碱基非常丰富,约有 70余种。每个tRNA分子至少有2 个稀有碱基,最多有19个。
多数分布在非配对区,尤其反 密码子3‘端附近部位,且大多 为嘌呤核苷酸。
4 生物信息的传递 (下)
基因的遗传信息在转录过程中从 DNA转移到mRNA;再由mRNA将 遗传信息表达为蛋白质中氨基酸顺 序的过程称翻译,即蛋白质的生物 合成过程。
• 蛋白质是生物信息通路上的终产物,一个活细胞在任何发育阶段都需要 数千种不同的蛋白质。因此,活细胞内时刻进行着各种蛋白质的合成、 修饰、运转和降解反应。
用其它2个碱基的共聚物进行类似的实验,也可以推断出其他密码子的碱 基组成,但不能确定密码子中碱基排列。
蛋白质翻译

翻译调控因子(延伸因子)的作用确保只有负载 正确的氨酰-tRNA才能进入核糖体的A位,形成稳 定的结合。
如果错误的氨酰-tRNA进入核糖体的A位,那么由 于错误配对,其缔合能偏低,仅为正确配对的 1/3000,延伸因子会将错误进入的aa-tRNA清除。
原核生物:EF-Tu
真核生物:eEF-1
合,再结合mRNA。
(一)真核生物蛋白质翻译起始
(1)40S核糖体小亚基与起始因子eIF-1和eIF-3结 合,使核糖体大小亚基分离;
(2)形成eIF-2-Met-tRNAMet-GTP三联体复合物; 它们与40S小亚基(包括eIF-1和eIF-3)P位点结 合,形成43S前起始复合物。
(3)在帽子结合复合物起始因子eIF-4F的帮助下, 前起始复合物与mRNA的5’端结合,形成起始复 合物。 eIF-4F复合物包括: eIF-4E(结合到mRNA的5’帽 子结构上)、eIF-4A(解旋酶活性)和eIF-4G(连接 eIF-4E与eIF-3)。
原核生物:EF-T (EF-Tu, EF-Ts)和EF-G
1、AA-tRNA与核糖体A位点的结合
需要消耗GTP,并需EF-Tu、EF-Ts两种延伸因子来进 行能量的再利用。
能量的再生: EF-Tu-GDP+ EF-Ts
EF-Tu-Ts + GDP
EF-Tu-Ts + GTP EF-Tu-GTP + EF-Ts 重新参与下一轮循环
二、真核生物蛋白质翻译(起始) 三、保证蛋白质翻译准确起始的机制(翻译 保真性) 四、复习题
一、原核生物翻译的起始
1. 蛋白质合成装备的组装 2. 模板mRNA在核糖体上的准确定位 3. 起始氨基酸的插入
1. 氨基酸的活化
如果错误的氨酰-tRNA进入核糖体的A位,那么由 于错误配对,其缔合能偏低,仅为正确配对的 1/3000,延伸因子会将错误进入的aa-tRNA清除。
原核生物:EF-Tu
真核生物:eEF-1
合,再结合mRNA。
(一)真核生物蛋白质翻译起始
(1)40S核糖体小亚基与起始因子eIF-1和eIF-3结 合,使核糖体大小亚基分离;
(2)形成eIF-2-Met-tRNAMet-GTP三联体复合物; 它们与40S小亚基(包括eIF-1和eIF-3)P位点结 合,形成43S前起始复合物。
(3)在帽子结合复合物起始因子eIF-4F的帮助下, 前起始复合物与mRNA的5’端结合,形成起始复 合物。 eIF-4F复合物包括: eIF-4E(结合到mRNA的5’帽 子结构上)、eIF-4A(解旋酶活性)和eIF-4G(连接 eIF-4E与eIF-3)。
原核生物:EF-T (EF-Tu, EF-Ts)和EF-G
1、AA-tRNA与核糖体A位点的结合
需要消耗GTP,并需EF-Tu、EF-Ts两种延伸因子来进 行能量的再利用。
能量的再生: EF-Tu-GDP+ EF-Ts
EF-Tu-Ts + GDP
EF-Tu-Ts + GTP EF-Tu-GTP + EF-Ts 重新参与下一轮循环
二、真核生物蛋白质翻译(起始) 三、保证蛋白质翻译准确起始的机制(翻译 保真性) 四、复习题
一、原核生物翻译的起始
1. 蛋白质合成装备的组装 2. 模板mRNA在核糖体上的准确定位 3. 起始氨基酸的插入
1. 氨基酸的活化
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
氨酰基tRNA进入A位
新的氨基酸-tRNA的进位依赖Tu-Ts因子和GTP的协助
肽键的形成
肽键的形成由肽酰基转移酶催化(此酶具有核酶的活性)
原核生物肽链的延长
核糖体沿着mRNA 5’——3’方向移位,多肽链沿着N端向着C端延伸, 须有EF-G因子和GTP参与,空载的tRNA从E位点离开
2.4 肽链的终止
蛋白质翻译系统示意图
1.1 信使RNA (mRNA)
mRNA是遗传信息的载体(载有遗传密码,genetic code),
是合成蛋白质的蓝图(模板),它以一系列三联体密码子
(codon)的形式从DNA转录了遗传信息。每个密码子代表一个 氨基酸。
mRNA占细胞总RNA的5-10%,不稳定,寿命短。
原核的mRNA 是多顺反子;真核的mRNA 是单顺反子。
倒L形的三级结构
tRNA的功能是解读mRNA上的密码子和搬运氨基酸。 tRNA上至少有4 个位点与多肽链合成有关:即3’CCA氨基酸接受位 点、氨基酰-tRNA合成酶识别位点、核糖体识别位点和反密码子位点。 每一个氨基酸有其相应的tRNA携带, 氨基酸的羧基与tRNA的 3’ CCA-OH 以酯键结合。而tRNA的反密码子可以与mRNA上相应的密码 子互补结合, 以使氨基酸正确“对号入座” (因为密码子的序列对应于
多肽链上 的氨基酸序列)。
密码子与反密码子的配对方式
变偶性或摆动性——反密码子5’端的碱基与 密码子的第三位配对不严格
1.3 核糖体(ribosome)与核糖体rRNA
核糖体是rRNA 与几十种蛋白质的复合体,有大、小两个亚基构成。
含有合成蛋白质多肽链所必需的酶、起始因子(IF)、延伸因子(EF)、
多个核糖体结合在同一条mRNA上,由5’——3’进行翻译,形成 多核糖体(polyribosome),以提高翻译的效率
3 多肽链的修饰和加工
(1)N—端修饰 原核生物修饰时是由肽甲酰基酶除去甲酰基,多数 情况甲 硫氨酸也被氨肽酶除去,真核生物中甲硫氨酸则全部被切除。 (2)多肽链的水解切除 水解切除其中多余的肽段,使之折叠成为有 活性的酶或蛋白质。如酶原激活 (3)氨基酸侧链的修饰 氨基酸侧链的修饰包括羟化、羧化、甲基化 及二硫链的形成等。 (4)糖基化修饰 糖蛋白是细胞蛋白质组成的重要成分。它是在翻译 后的肽链上以共 价键与单糖或寡聚糖连接而成。糖基化是在酶催化 下进行的。
分泌型蛋白质在翻译过程中通过信号肽协助转入内质网的机制
信号肽(signal peptide)是在新生的多肽链中,可被细胞识别系统识别的 特征性氨基酸序列,在蛋白质翻译过程中或翻译后的定位发挥引导的作用。
本章结束
第15章
蛋白质翻译
Protein Biosynthesis (Translation)
本章主要内容
翻译系统 蛋白质生物合成的过程 多肽链翻译后的修饰 蛋白质的到位
原核
真核
基因在原核和真核细胞中最终得到表达
1 蛋白质的翻译系统
原料氨基酸,20种 mRNA是合成蛋白质的“蓝图(或模板)” tRNA是原料氨基酸的“搬运工” rRNA与多种蛋白质结合成核糖体,作为合成多肽链的 “装配机(操作台)”
5’AUG CCA UGC ACC CGG GUU …………..CAA UAG 3’ Pro Cys Thr Arg Val …………..Gln
密码有简并性(degeneracy),即一个氨基酸可由几个密码子表示; 通用性,大多数生物使用同样的遗传密码,但也会有所偏爱。
遗 传 密 码 表
1.2 转运RNA (tRNA)
氨基酸和选择其所对应的tRNA,使两者连接起来(利用ATP)。
反应如下:
AA tRNA ATP AA-tRNA AMP PPi
氨基酸的羧基与tRNA 的3’端CCA-OH 以酯键相连,因此其氨基是 自由的。
氨酰基-tRNA合成酶
tRNAfmet fMet-tRNA合成酶
Met 酯酸 一般是甲硫氨酸, 其氨基要甲酰 化成为甲酰甲硫 氨酸,予以保护。
甲酰基
氨基酸与tRNA的 连接方式
fMet-tRNA
2.2 起始复合物的形成
首先IF3、IF1帮助30S小亚基
与mRNA结合,IF2和GTP帮助
甲酰甲硫氨酸-tRNA与AUG配 对,接着IF3脱离,形成30S 起始复合物。50S大亚基进入, IF1和IF2脱离,形成70S起始
遗传密码
上世纪60年代,利用均聚核苷酸实验,破译了遗传密码。 遗传密码为三联体(每三个碱基代表一个氨基酸),由5’到3’阅读, 无间断。即使在少数重叠基因(如病毒)中,其开放阅读框架(open reading frame, ORF)仍按此原则。 起始密码为AUG, 终止密码为UAA,UAG和UGA。
3. 肽酰基位点(P位点),可结合肽酰基tRNA(肽-tRNA); 4. 脱氨基酰tRNA的出口位(E位点) ;
5. 肽酰基转移酶中心,是形成肽键的位
点等。
2
蛋白质的生物合成过程
2.1 氨基酸的活化
所有参与合成多肽链的氨基酸都要激活,并由数十种高度专一的 氨基酰-tRNA合成酶催化。该酶由两个识别位点,它们能识别特定的
rRNA只有4-5种,但占细胞 RNA 的大部分 。 图为原核生物的两种RNA 16S rRNA和5S rRNA 的结构
核糖体的结构
核糖体有4个基本功能
1. 容纳mRNA,并能沿着mRNA由5’—— 3’ 移动,由tRNA解读其密码; 2. 氨基酰位点(A位点),可结合氨基酰 -tRNA(AA-tRNA);
氨基酸进位,肽链形成和延伸,核糖体沿着mRNA的5’— —3’ 方向移位,循环往复,新合成的肽链由N端向着C端不断延 长,直至mRNA上出现终止密码,相应的肽链释放因子RF1 (对应UAA、UAG),RF2(对应UAA、UGA)占据A位。肽 链的合成终止,并从核糖体上释放。
核糖体大、小亚基解聚,并进入下一轮合成。
释放因子(RF)等。
原核的核糖体(70S)= 30S小亚基 + 50S大亚基
30S小亚基: 16S rRNA + 21种蛋白质
50S大亚基: 23S,5SrRNA + 34种蛋白质
真核的核糖体(80S)= 40S小亚基 + 60S大亚基
40S 小亚基: 18S rRNA + 33种蛋白质 60S 大亚基: 28S,5.8S,5SrRNA + 45种蛋白质
复合物,需要GTP。甲酰甲硫
氨酸-tRNA处于P位。
一个嘌呤丰富区
起始密码
起始密码AUG上游的S.D序列(一个嘌呤丰富区)与其核糖 体小亚基中16S rRNA的3’端互补结合有利于30S起始复合 物的形成
2.3 肽键的形成和延伸
(注意新的AA-tRNA如何定位,第一个肽键如何形成,核糖体如何移动…)
原核tRNA有30-40种,真核有50-60种,含70-90个核苷酸, 并有多种稀有碱基。
tRNA是最小的RNA, 占细胞总RNA 的15%左右,其功能 是搬运氨基酸和解读密码子。
tRNA 具有“四环一臂”和“三叶草” 形的典型结构。
注意:3’端CCA氨基酸受位和反密码子环
tRNA的结构—“四环一臂”
沉降系数 S
生物大分子在离心场中沉降,受到三种力的影响,它们是离
心力,浮力和摩擦力。物质在单位离心力场的沉降速度是个定值, 称为沉降系数(sedimentation coefficient) 。
蛋白质、核酸等的沉降系数在1 X 10-13到 200 X 10-13秒 之间。
为方便将10-13秒作为一个单位,称Svedberg单位,用S表示。 其数值不仅与物质分子的质量有关,也与分子的形状有关。