腈水合酶制备关键技术研究
腈水合酶研究进展

腈水合酶研究进展摘要从腈水合酶的结构、分布、酶学性质及其基因克隆和生产中的利用形式等方面简要阐述了腈水合酶的研究进展。
关键词腈水合酶;酶学性质;基因克隆腈水合酶(Nitrile hydratase,EC4.2.1.84)是一类可以催化腈类物质转化成相应酰胺类物质的酶[1],其最初是由日本学者Asano发现并命名的。
微生物的腈水合酶作为生物催化剂应用在有机物合成的工艺上具有巨大的潜力,它的反应条件温和、产率高、副产物少、产物的自聚损失少,具有区域选择性、立体选择性和光激活性,可广泛应用于氨基酸、酰胺、羧酸及其衍生物的合成。
更重要的是,它环境污染小、成本低、符合绿色化工发展理念,有着化学方法无法替代的优越性,从而促进了精细化工产品、可降解塑料和手性药物以及维生素等方面的研制和开发。
因此,20多年来一直受国内外研究者的广泛关注。
1腈水合酶的产生菌及其结构无论是腈类物质还是酰胺类物质对人体来说都是极其有毒的[2],然而有些微生物却能利用它们作为自身生长的碳源和氮源。
原因就在于这些微生物能够自身合成降解腈类物质的酶—腈水合酶(NHase),腈水合酶是某些微生物在进行肟或尿代谢过程中所产生的一种蛋白[3]。
目前已知可产腈水合酶的菌株有红球菌、诺卡氏菌、假诺卡氏菌、棒状杆菌、假单胞菌、产碱杆菌、短杆菌等。
可见产腈水合酶的菌的分布相当广泛[4]。
不同菌株的腈水合酶在结构上存在着一定的差异,但是也存在来自不同菌株的腈水合酶在结构上的相同。
尽管不同菌株的腈水合酶在氨基酸序列上存在差异,但是它们仍然存在着一个共同点,那就是在酶的活性部位都含有螯合的金属离子作为辅助因子。
根据所含辅助因子的不同,腈水合酶可分为2类,一类为高分子量腈水合酶(H-NHase),其所含金属离子为钴;另一类为低分子量腈水合酶(L-NHase),其所含金属离子为铁。
尽管铁型和钴型腈水合酶在氨基酸序列上具有极大的同源性,但这2种酶在生物转化活性和底物特异性上还是有着很大的差异。
新型耐热腈水合酶的异源表达及酶学性质研究

新型耐热腈水合酶的异源表达及酶学
性质研究
新型耐热腈水合酶是一种具有高耐热性的腈水合酶。
腈水合酶是一种重要的酶,可以在高温条件下进行水合反应,广泛应用于食品、医药和化工等领域。
然而,目前已知的腈水合酶在高温条件下的耐受性较低,因此开发新型耐热腈水合酶具有重要的意义。
异源表达是指将外源基因转移到另一个生物体中,使得该生物体能够表达出外源基因的蛋白质。
新型耐热腈水合酶的异源表达通常采用转基因技术,将耐热腈水合酶基因转移到可以表达的生物体中,以获得新型耐热腈水合酶的蛋白质。
在异源表达完成后,可以进行酶学性质研究。
酶学性质研究是指对酶进行物理化学测定,了解酶的活性、热稳定性、pH稳定性、选择性等性质。
这些性质对于评价新型耐热腈水合酶的高效性和适用性具有重要意义。
新型耐热腈水合酶的异源表达和酶学性质研究是提高腈水合酶应用效率的重要手段。
通过异源表达获得的新型耐热腈水合酶具有更高的耐热性和高效性,可以在高温条件下进行水合反应,为食品、医药和化工等领域的应用提供新的选择。
产腈水合酶优良菌种的选育及其在生产中的应用研究

生 提 2 0 生 物 催 化 珐 生 产 丙 烯 酰 胺 (c ya d , a r l mie AM ) 量 , 产 中得 到 的 结果 表 明 , 高 丙 烯酰 胺 琼 脂 , 0. 。
所 的催化剂 , 能催化 丙烯腈 (cyo i i , N) 溶 液 百 分 比 浓 度 可 减 少 蒸 气 用 量 。 以 提 arlnt l A re
摘 要 : 文通 过 出发 茵株 N c ri.p 1 3 瓶 发 酵 9 h 本 oada s . 摇 6 6 后分 别 向 发 酵 液 中 间歇 加 入 丙 烯腈 和 催 化 生 成 高 浓 度 丙 烯 酰 胺 分 别 对 茸 兰 的毒 害和耐受作 用以及发酵液 中残存 茵丝涂布平板 置于1 8℃低 温培 养成单茵落 , 筛选到一株比 出发 茵株N cr i.p 1 3 oadas .6 相对腈水合 酶酶活提 高 了7 % 2 号优 良 茵株 。 茵株 在 催 化 腈 水 合 生 成 丙烯 酰 胺 的 反 应 中 , 酰 胺 酶 酶 活 性 此 出发 茵株 降低 了近 5 % 6的 4 该 产 0 丙 蚌 蔬 胺 水 溶 液 的 电 导 率 比 出发 茵株 降 低 了4 %, 应 次 数 提 高 了3 3 次 , 烯 酰 胺 放 料 浓 度 提 高 了5 9 %。 5 反 .3 丙 .3 关 键 词 : 卡 氏 茵 腈 水合 晦 丙烯 酰 胺 耐 受 菌种 选 育 诺 中图分 类 号 : Q9 5 T 2 文 献标 识码 : A 文 章编 号 : 6 4 9 X( 0 1O () 0 6 3 l 7 -0 8 Z 1 ) 4b一0 0 -0
No a d a.p.6 为 本 实验 室保藏 。 c r i s 1 3, 1 1 2培 养基 ..
1 I 2 I斜 面 和 平 板 培 养 基 ( L) ... g/
腈水合酶催化反应条件的研究

腈水合酶催化反应条件的研究刘舒;沈旭丰;孙先锋【摘要】Taking the enzyme of free cell as the catalyst, the effects of dilution factor, concentration of acrylonitrile, temperature, pH value, concentration of cells and concentration of acrylmide on the activity of nitrile hydratase were studied. The result shows that the temperature, the concentration of acrylonitrile and the concentration of gcrylamide are the most important among these factors. The activity of nitrile hydratase is 4 901U/mL(broth) at the reaction temperature 28℃ t he concentration of acrylonitrile is 40g/L;the activity of nitrile hydratase in the solution of 40% acrylamide is just 40% of that in solution of 5% acrylamide. It is found that the activation energy of the nitrile hydratase is 40. 291kJ · mot1 through the research at the different reaction temperature.%以自由细胞的酶作为催化剂,研究了菌体稀释倍数、丙烯腈浓度、温度、pH值、菌悬液用量和丙烯酰胺浓度对腈水合酶活性的影响,结果表明,温度、丙烯腈浓度和丙烯酰胺浓度是影响酶活性的主要因素.反应温度在28℃时酶活性达到最大为4091U/mL菌液,丙烯腈浓度采用40g/L.当反应体系中丙烯酰胺浓度为40%时,其酶活只有丙烯酰胺浓度为5%时酶活的39.2%.对不同反应温度下酶活的研究,得出腈水合酶催化反应的活化能为40.291kJ · mol-1.【期刊名称】《纺织高校基础科学学报》【年(卷),期】2012(025)001【总页数】6页(P83-87,106)【关键词】丙烯酰胺;腈水合酶;丙烯腈;催化反应【作者】刘舒;沈旭丰;孙先锋【作者单位】西安工程大学环境与化学工程学院,陕西西安710048;西安工程大学环境与化学工程学院,陕西西安710048;西安工程大学环境与化学工程学院,陕西西安710048【正文语种】中文【中图分类】Q559在微生物转化生产丙烯酰胺的过程中,酶催化反应是关键环节,因此研究酶催化反应的条件成为必然.腈水合酶(Nitrile hydratase,EC4.2.1.84)是以硫原子和半胱氨酸-亚磺酸残基(Cysl12)为绝对中心的一类可催化腈水解的酶.随着科研人员对腈水合酶结构、催化机理、生物合成调节以及酶学性质的深入研究,腈水合酶已被成功应用于工业生产丙烯酰胺上.最早提出用微生物法生产酰胺的是法国研究者Galzy及其研究组,他们发现了一种能催化腈水解的微生物Brevbacteriu m R312,可催化合成丙烯酰胺(A M),但是酶活很低[1].1982年,京都大学山田秀明研究组发现了Pseudomonas Chlor aphis B32,经过驯化其酶活达到1 260个单位[2].1988年,山田秀明等发现了一种性能更好的腈水合酶产生菌Rhodococcus r hodococcus J1,其酶活达到了1 630个单位,一度成为日本酰胺生产的工程菌.高活力菌种的成功应用,使日本日东公司年产量从最初的0.4万t/a迅速提升至3万t/a[3].采用酶生物催化剂代替化学催化剂已成为绿色化学发展的方向之一[4].与化学法相比,微生物法制备丙烯酰胺具有反应在常温常压下进行、选择性高、活性高、能耗低、环境污染小、成本低、产物光学纯度高等优点[5].目前,国内用生物法生产丙烯酰胺多采用海藻酸盐包埋法固定化细胞的工艺,而该工艺本身存在通透性差、固定化细胞强度小以及引入离子等问题,另外在固定化阶段会损失相当一部分酶活.正由于酶活的损失,一批固定化的细胞往往水合反应3批左右就需要更新[6].利用自有细胞直接进行水合反应并实现连续化生产能较好地解决以上问题.本文对诺卡氏菌Nocar dia sp.ZT-2自由细胞的腈水合酶反应条件进行了研究,以期优化酶反应条件,提高酶活性.1 实验1.1 原料丙烯腈(上海三浦化工有限公司,分子量53.06,化学纯)、丙烯酰胺(北京化学试剂公司,分子量71.08,分析纯).1.2 实验菌株诺卡氏菌Nocar dia sp.ZT-2(从咸阳第二印染厂污水处理厂的活性污泥中筛选,由西安工程大学生物工程实验室保藏并鉴定.)1.3 培养基1.3.1 斜面培养基(g/L)牛肉膏5,蛋白胨1,Na Cl 5,琼脂4,p H 7.2.1.3.2 发酵培养基(g/L)葡萄糖20,酵母膏5,尿素6,味精(谷氨酸钠)1,KH 2 PO4 0.5,K 2 HPO4 0.5,M gSO4·7 H 2 O 0.5,Co Cl2·6 H 2 O 0.08 mmol/L,吐温80 1,p H 7.2.1.4 自由细胞的培养方法将菌株从斜面接种于250 mL发酵摇瓶中,装液量为25 mL发酵培养基,在28℃下培养96h,摇床转速为180r/min.1.5 生物量的测定将发酵液稀释一定倍数后,在460n m波长下测量吸光度值.根据测定的菌体干重和吸光度标准曲线,一个OD值相当于每L发酵液有0.45g干菌重量.1.6 操作过程1.6.1 菌悬液的制备取一定量的发酵液,在6 500r/min条件下离心8 min,弃上层清液,用p H=7.0的磷酸缓冲液洗涤菌体.在同样条件下重复离心分离和洗涤两次,制成与原发酵液菌体浓度相等的菌悬液,冰箱保存备用.1.6.2 催化水合反应取19 mL 50 mmo L p H为7的磷酸缓冲溶液和1 mL纯丙烯腈,放入250 mL具有磨口玻璃塞的三角瓶中,恒温至28℃.取一定量的菌悬液根据具体实验要求稀释适当数倍,取1 mL于三角瓶中,准确反应5 min,然后加入1 mL 4 mol/L的盐酸终止反应[7].1.7 酶活测定方法除在丙烯酰胺浓度对产物影响中丙烯酰胺生成量用液相色谱测定外,其余丙烯酰胺生成量及所有酶活的测定采用KBr O3-KBr法.液相色谱操作条件如下:流动相为甲醇∶水=15∶85,流速1 mL/min,温度25℃,检测波长205n m.产物计算方法为:采用外标法以峰面积为标准配置各种浓度梯度的丙烯酰胺标准样品,拟合出丙烯酰胺标准校正曲线.KBr O3-KBr 法测定丙烯酰胺生成量及酶活:丙烯酰胺的生成量(mg)=0.5×71.0×C(V 0-V)×V 2/V 1,酶活=C(V 0-V)×(1/2)×V 2/V 1×1 000/t.C为Na2 S2 O3 溶液的摩尔浓度,mol/L;V 0 为空白消耗Na2S2 O3溶液的体积,mL;V为滴定样品消耗Na2S2 O3溶液的体积,mL;1/2为反应所生成Br 2与Na2 S2 O3溶液的化学计量比;V 1为反应所取上清液的体积,mL;V 2为上清液总体积,mL;1 000为mmol转换成μmol;t为反应时间,min;0.5为生成丙烯酰胺的摩尔数;71.0为丙烯酰胺的分子量.2 结果与讨论表1 菌液稀释倍数与酶活稀释倍数4 8 12 16 20 30 40酶活/U·mL-1 2 853 2974 2 910 2 737 2 658 2 808 2 5302.1 稀释倍数对酶活的影响因为培养所获得的菌液酶活都比较高,而在后面的研究中,为了更好地反映各种因素对催化反应的影响,是不期望反应过快的,所以需要对菌液进行稀释.因此,分别将菌液稀释4,8,12,16,20,30,40倍,水合反应后分别测量此时的酶活,然后将测得的酶活乘以稀释倍数,换算得到原始的酶活.换算后结果如表1所示.以稀释16倍的酶活为1,求出不同稀释倍数下的相对酶活U,然后作图,结果如图1所示.从图1可以看到,菌液经稀释后,对折算的原始酶活,在稀释4~12倍之间,酶活相对偏高但在1.1以内;而在16~30倍之间折算的酶活相对偏差较小,误差保持在±10%以内.可以认为,在研究的范围内,稀释倍数对于酶活可以用折算的办法来补偿.2.2 丙烯腈浓度对酶活的影响在催化水合反应中,丙烯腈是一种底物,其浓度对酶活可产生两种影响.一方面,由于底物与中间络合物(ES)生成的三元复合物(SES)不可分解成产物,且具有生理毒性,浓度过高对菌体有害,会造成酶活的快速下降;另一方面,从反应动力学的角度来讲,底物浓度的提高又有利于提高反应速率[8-10].恰是这两种影响正好相反,所以研究丙烯腈浓度对酶活的影响十分必要.在总体积为25 mL,反应温度为28℃,p H 为7.0情况下,分别考察丙烯腈浓度为10g/L,20g/L,30g/L,40g/L,50g/L,60g/L,80g/L,100g/L,120g/L,140g/L,160 g/L时对水合反应体系中反应速率的影响,结果如图2所示.图1 稀释倍数对酶活的影响图2 丙烯腈浓度对酶活的影响如图2所示,当丙烯腈浓度小于40g/L时,酶活随浓度的增加而快速上升;而在丙烯腈浓度40g/L至120g/L之间,随着浓度的增加,其酶活增加比较平缓,并且在丙烯腈浓度120g/L时,酶活达到最大,为4 125 U/mL,然后酶活随丙烯腈浓度的增加而缓慢降低.可以看出,在丙烯腈浓度大于60g/L以后,酶活还能随着浓度的增加而上升,但是上升幅度比较小,说明高浓度底物已开始对酶活产生抑制作用了.本实验中,丙烯腈浓度最大为160g/L,但此时还观察不到对酶活明显的抑制作用.这说明在本研究的范围内,菌株ZT-2所产的酶对底物的耐受性是比较强的.在后续的实验中,采用丙烯腈浓度为40g/L,虽然丙烯腈浓度在120g/L时所产酶活最高,但相比40g/L时的酶活仅提高了31%,而底物用量却是前者的3倍之多,也就是说丙烯腈转化率降低了.可见采用高的底物浓度并不是最佳选择.根据丙烯腈浓度与酶活的关系,可求得腈水合酶催化反应的米氏常数.求得的米氏常数K m=14.048g/L,表示的是反应速率达到最大速率一半时的底物浓度.2.3 温度对酶活的影响由图3可见,从18℃到28℃之间,酶活随着反应温度的升高而加快,在28℃时达到最大,测定的酶活为4 091 U/mL菌液;而当反应温度大于30℃后,酶活下降非常快,到40℃时酶活只有28℃时的44.83%,不及其一半,表观酶活仅为1 834 U/mL菌液.分析其原因,可能是酶在高温下失活所造成的.由阿累尼乌斯公式可知,反应速率常数k的对数应该与1/T成正比,反应速率常数k与表观酶活r 成正比.因而,在低于301 K时,以ln(r/U·mol-1)对1/T 作图,可得到一条直线,如图4.直线拟合结果由此可以算出反应的活化能为E a=40.291kJ·mol-1.2.4 p H对酶活的影响目前从文献报道来看,水合反应大都在中性环境下进行,在此环境下反应速率达到最大值,多数研究者得到的结果也多在7~7.5之间达到最大反应速率[13-14].为此,本实验在总体积为25 mL,反应温度为28℃,丙烯腈浓度为40g/L的情况下考察了p H 为5,6,6.5,7.0,7.5,8.0,9.0时对酶活的影响情况.结果如图5所示.由图5可知,p H在低于6.5时,酶活随p H的升高而逐渐增大,而在6.5~7.0时,酶活快速上升;当体系p H超过7.5时酶活迅速下降,在7.0~7.5这个区间内酶活维持在较高水平,说明反应体系的p H处在中性范围有利于酶的高效表达,从而获得较高的酶活.因此在以后的研究中水合反应体系的p H为7.3.图3 温度对酶活的影响图4 温度与酶活的线性拟合注:粗线为拟合曲线,细线为数据点连线.2.5 菌悬液用量对反应速率酶活的影响酶作为生物催化剂,其催化反应的速度直接取决于酶浓度[15].本实验考察在总体积为25 mL,反应温度为28℃,丙烯腈浓度为40g/L,p H为7.3的情况下菌体加入量对酶活影响.实验中发现当菌悬液加入量大于20 mL时,酶活开始下降,估计是受到底物限制的原因.所以只研究了菌悬液加入量为2 mL,5 mL,8 mL,11 mL,14 mL,17 mL,20 mL时对产酶的影响.图5 p H对腈水合酶活性的影响图6 菌悬液用量对反应速率的影响由图6可知,酶活随着菌悬液用量的增加而逐渐增大,在菌悬液加入量为20 mL 时,酶活达到了最高值.这符合酶促反应动力学的基本原理,也就是当底物过量时,酶活随着菌液量的增加而增大,当底物浓度限制时,酶活有最大值.当菌悬液加入量V cell大于20 mL时,酶活开始减少.当V cell=20 mL时,其最大酶活达5 308 U/mL.以菌悬液V cell加入量对酶活作图,并进行拟合,得到直线方程通过拟合方程可以知道一定体积下菌体加入量的酶活,因而在实际生产中,在保证后期分离及生产成本的前提下,采用合理的菌悬液用量可以保持相对较高的酶活性.2.6 丙烯酰胺浓度对产物的影响产物丙烯酰胺(A M)是一种高毒性的物质,对细胞有较大的生理毒性,能够破坏酶的三维结构并在产物释放前与酶形成中间产物EP的复合物,从而使酶不能与底物接触,降低了酶与底物接触的机会,抑制了酶的活性,也就降低了整个反应的速率[16-17].在不同丙烯酰胺浓度的水合反应体系中得到丙烯酰胺浓度结果如表2所示.表2 丙烯酰胺浓度对酶活的影响反应前A M浓度/*************反应后AM浓度/% 2.04 6.94 11.73 16.65 21.42 31.18 40.76由表2可知,丙烯酰胺作为反应的产物,其浓度的增加能够极大地抑制酶的活性.当其在体系中的浓度低于20%时,对酶活的影响并不是特别明显,若以添加丙烯酰胺为5%的体系作为标准,其相对酶活都在70%以上.而当其浓度大于30%时,酶活急剧下降,到浓度为40%时,相对酶活已不足40%,说明丙烯酰胺浓度的增加能够极大地抑制体系的酶活,降低反应速率.因此,在实际生产中,及时分离出反应体系中生成的丙烯酰胺是非常必需的,也是目前亟待解决的问题.3 结论底物和产物浓度、催化反应温度、反应p H、菌悬液加入量是影响丙烯腈水合酶活性的主要因素,其中反应温度、底物和产物浓度是影响酶活的关键性因素.对于温度的选择,需要从两个影响方面权衡优化得出,一是温度升高有利于获得较高酶活,二是较高的温度会使酶失活.对于底物浓度,也应从两方面加以考虑,一方面底物浓度的增加有利于反应速率的提高,另一方面底物浓度的增加会抑制酶活性.产物浓度在水合反应中应保持在较低浓度范围,其在体系中的浓度低于20%,相对酶活能保持在70%以上,但是当体系中丙烯酰胺浓度大于20%时,酶活下降非常迅速.本文所得到的对酶活影响的参数,对工业用微生物法生产丙烯酰胺的操作条件优化提供了依据.【相关文献】[1] OHTSUKA Y,HASGA WA J,NAKATSUJI H et a1.Density functional st udy on geo metry and electronic str ucture of nitrile hydratase active site model[J].J Inter Quan Chem,2002,90(3):1 174-1 187.[2]刘金元,王慧琴,徐淑霞,等.腈水合酶研究进展[J].安徽农学通报,2008,14(4):111-113.[3]罗积杏,薛建萍,沈寅初.我国生物法生产丙烯酰胺的现状及其研发概况[J].上海化工,2007,32(2):17-21.[4]王浩.腈水合酶产生菌的分离筛选及其特性研究[J].微生物学杂志,1996,16(4):12-17.[5]胡常伟,李贤均.绿色化学原理和应用[M].北京:中国石化出版社,2002:98-99.[6]陈跖,孙旭东,史悦,等.微生物法生产丙烯酰胺的研究(Ⅱ)[J].生物工程学报,2002,18(2):225-230.[7]邓林,刘延岭,王忠彦,等.产腈水合酶菌株的驯化选育及其产酶条件的优化[J].食品与发酵工业,2005,31(1):53-56.[8] Y Reddy,S Kalju Kahn,ZHENG Y J,et al.Protein engineering of nitrile hydratase activity of papain:molecular dynamics study of a mutant and wild-type enzy me [J].Journal of the American Chemical Society,2002,124(44):12 979-12 990.[9] CHEN Zh,SUN X D,SHI Y,et al.Study on production of acrylamide by microbial method(11)-enzy me catalytic kinetics and de-active dynamics of nitrile hydratase [J].生物工程学院,2002,18(2):225-230.[10] ALIKIN V N,ALIAGA A,TERESA M,et al.Detoxification and recycling of wastes from biotechnological manufactuer of acrylamide[J].Ekologiyai Pro myshlennost Rossii,2005,12:14-16.[11] RUI A Pereira,GRAHA M Dan,RAINEY Fred A,et al.A novel ther mostablenitrile hydratase[J].Extremophiles,1998,2(2):347-357.[12] PADMAKUMAR R,ORIEL P.Bioconversion of acr ylonitrile to acr ylamide using a ther mostable nitrile hydratase[J].Applied Biochemistry and Biotechnology,1999,77(9):671-679.[13] YASUO Kato,TAKASHI Tsuda.Yasuhisa asano nitrile hydratase involved in aldoxi me metabolis m fro m Rhodococcus sp.Stran YH3-3,purification and characterization [J].European Journal of Biochemistry,1999,263:662-670.[14] NAGASA WA Tor u,NANBA Hir onori,RYUNO Koichiro,et al.Nitrile hydratase of pseudomonas chloror aphis B23,purification and characterization[J].European Jour nal of Biochemistr y,1987,162:691-698.[15] JUNA C,OL MSTEAD M M,MASCHARAK P K.A synthetic analogue of the active site of Fe-containing nitrile hydratase with car boxa mido N and t hiolato S as donors:synthesis,str uct ure,and reactivities[J].Jour nal of the American Chemical Society,2001,123(14):3 247-3 259.[16] LIU M,LI Ch,HUANG Y.et al.Impr oving the acrylamide-tolerance of nitrile hydratase in Nocar dia sp.by extreme cultivation[J].过程工程学报,2004,4(3):250-255.[17] ASTAUROVA O B,LEONOVA T E,POLIAKOVA I N,et al.Adaptation of r hodococcus r hodochrous M8,a pr oducer of acrylamide,to changes in ammoniu m concentration in the growt h mediu m[J].Prikladnaya Biokhi miya Mikrobiologiya,2000,36(1):21-25.。
腈水合酶研究进展

i( man t f( Re iDaa—m.o io ) 4 P st n <2 5 i
{
抽象 ,便 于普 通用户或者管 理决 策者使用 ,提 高了管理效 率。随着数字 灌区行业 的不 断发 展 ,将会更切 实的感受到
二者结 合 的重要 性 。
参 考 文 献
f1 1 肖乐斌 ,钟 耳顺 ,刘纪元等 . 三维 GI S的基本问题探讨 … . 中国
摘
要:该 文简要阐述 了腈水合酶的结构 、分布 、酶学性质及其基 因克隆和生产 中的利用形式 。
文献 标 识码 A 文 章编 号 1 0- 7 1( 0 8 4 1 卜O 07 7 3 2 0 )0 — 1 3
关键词:腈 水合酶;酶学性质 ;基 因克隆
中图 分类 号 Q 1. 849
学者 A a o sn 发现并命名的。微生物的腈水合酶作为生物催 化 剂应用在有机物合成的工艺上具有 巨大的潜力 。它 的反应条
酸 、酰 胺 、羧酸 及其 衍 生物 的合 成 。更 重要 的是 ,它对
环境 污染 小 、成本低 ,符合绿色化工发 展理念 ,有着化学
基金 项 目:河 南农业 大学博 士基金 ( 0 6 3 ) 2002 1 作者 简介 :刘元 金( 9 2~ ,在读 硕 士研究 生 ,从事 微生 物酶 工程研 究 。 通 讯联 系人 。 18 )
产腈水合酶菌株的驯化及其产酶条件优化研究

c n ii so h e te ta t n r t r sf l ws l c s 0 / u e 0 g L, c y o < 4 , c y a d o d t n ft e b s x r ci a i a ea ol o o o o :g u o e2 g L, r a0. 6 / a r l n % a r l mi e< 2 % , o 0 3 / p 7g L, H 0 a d t mp r t r 8
产腈水合 酶菌株 的驯化及其 产酶条 件优化研究
邱 峰 李 志 东 , , 李 娜 张 洪林 王 战 勇 , , , 邱 江
( . 宁石 油化 工大 学环境 工程 系 , 宁抚 顺 1 3 0 ; . 顺石 油三厂 , 宁抚 顺 13 0 ) 1辽 辽 10 12 抚 辽 1 0 1
F su 1 0 1 L a nn C ia; . uh nOi P a t . , uh n 1 3 0 , io ig, hn ) uh n 13 0 , io ig, hn 2 F s u l ln No 3 F s u 1 0 1 L a nn C ia
AB T S RAC T:A a t ra sr i a i g hg e i i y r t s c i i n c y o i i o c n a in d t l r b c e il tan h v n i h rn t l h d a a e a t t a d a r ln t l c n e t r t e o e — re v y re o a c , u e e c r i p L n e n mb r d No a d a s NS 6, e eo e y r p a e u c lu i g o ti h r t o t i i g a r — Y0 i d v l p d b e e t d s b u t rn fi n t e b o h c n an n c y s
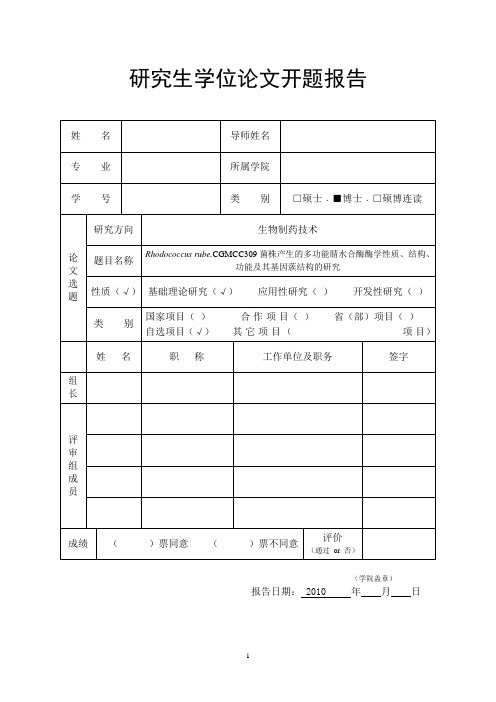
研究生学位论文开题报告-腈水合酶酶学性质、结构、功能及其基因蔟结构的研究
2、本课题研究领域在国内外的研究动态及发展趋势:
2.1 腈水合酶理论的研究动态及发展趋势 腈水合酶(NHase; E.C.4.2.1.84)催化腈的水合作用转变成相应的酰胺。 酰胺然后经酰胺酶 (E.C.3.5.1.4).转化成羟基酸。这组酶之所以受到广泛的研究主要是因为它们作为催化剂成功 地应用于大规模的商业生物转化之中,由Nitto化学公司进行的大规模的工业化转化生产 (6000t/年) 这一成就的取得是由于使用了来自Pseudomonas chlororaphis B23菌株的腈水 。 合酶[6] 。使用Rhodococcus rhodochrous J1产生的腈水合酶,这一过程进一步改进,使得年 产丙烯酰胺达30000吨以上[7]。另一个方面他们对环境生物修复具有潜在的应用价值。生物 催化优于传统方法的优势[8,9]有很多,对环境更加友好,使合成产量更高,并具有化学、区 域和立体选择性,在更加的温和的反应条件下进行的特点。 2.1.1 腈代谢酶的发现、分布和分离 许多微生物菌株可以经亚乙基羟胺水化酶作用下,代谢催化亚乙基羟胺[10]。亚乙基羟 胺是合成天然产物的中间体。 亚乙基羟胺水解酶基因像腈代谢酶 (腈水合酶和酰胺酶) 一样, 具有同样基因蔟和操纵子。该途径的作用是分流亚乙基羟胺进入中心能量代谢机制。在
3
和钴型(Co-type)腈水合酶[32,33]。大多数腈水合酶的最小的功能单元由两个亚基组成,还包括 一个金属离子单位。迄今为止,至少有两个已经被描述的酶有同型二聚体功能单元,以2α 表示(Agrobacterium tumefaciens[34] (Trott et al., 2001) 和Corynebacterium sp. C5 [35] (Yamamoto et al.,1992)),一个有一个单体功能单元。不同亚基α和β的大小范围从23kDa到 30kDa不等。天然分子量在30kDa[36](Rhodococcus equi SHB-121菌)和500-530kDa[37] ( Rhodococcus rhodochrous J1之间。这样腈水合酶就分成了轻型腈水合酶(L-NHase)和重型 腈水合酶(H-NHase),特别是来自于Rhodococcus rhodochrous J1.的酶。几个腈水合酶的晶体 结构已经阐明(Rhodococcus sp. R312,程序数据库登陆号[38]为1AHJ/2AHJ;Rhodococcus sp. N-771[39],Nagashima et al.,1998;Pseudomonas thermophila,程序数据库登陆号[40]为1IRE, Miyanaga et al.,2001;Bacillus smithii SC-J05,程序数据库登陆号[41]为1V29,Hourai et al., 2003)。所有这些酶大体折叠层次是保守的。第一个细致描述的腈水合酶结构,分辨率是 2.65,报道于1997年(Huang et al., 1997)。这个酶属于铁型亚群。酶的α亚基由一个长的延 伸的N-端臂和一个球状的C-端结构域组成。这个C-端的球状结构域很不寻常,详细的有4层 结构,即α-β-β-α。β亚基以30个氨基酸残基的长环开始,在αβ双体内将α亚基包缠起来。随 着是5个螺旋结构域,跟着是一个由6个反平行链组成圆环。Rhodococcus sp. R312的腈水合 酶的α和β亚基形成了一个紧密的二聚体结构,带有一个很大的3800 2内表面积。铁中心位 于内部表面积中的一个大的开放洞穴内部,与金属离子配位的蛋白配基由α亚基提供。 cys110α, cys113α, cys115α的侧链和ser114αand cys115α残基的主链酰胺氮原子形成了配体 球。在晶体结构中观察到的配体球是这样的形式S3N2X,带有5个一致的配体,占据八面体 的5个顶点。第6个位置(由X代表)被推测是一个氢氧化合物或水分子以酶的活性形式存在 和一氧化氮以酶的非活性形式存在[42]。来自于保守残基arg141和arg56的胍基同来自于 cys115α和 cys113α的硫原子形成氢键,这样就保正了他们的正确的位置。来自Rhodococcus sp. N-771菌株的另一个铁型腈水合酶[43] (Nagashima et al., 1998)表明cys114 αand cys112α (相 当于Rhodococcus sp. R312菌株的cys115α和cys113 α)是转录后各自被修饰成半胱次磺酸和 半胱亚磺酸。在这个酶中,arg141β和 arg56β同这些修饰的半胱残基的氧原子形成氢键。现 已表明, α亚基的Val-Cys-Ser(Thr)-Leu-Cys-Ser-Cys序列形成了金属结合模型 (即爪状模型) , [44] 在所有腈水合酶中 都具有高度的保守性。钴型腈水合酶有一个苏氨酸替代了铁型腈水合 酶的丝氨酸。 这种取代方式一般认为是对于不同的辅因子有不同的偏好。 由金属离子占据的 开放洞穴接近弥散的溶胶中, 被认为是酶的可能的活性位点。 这种观点已经受到了底物类似 物(碘乙腈)进入Rhodococcus sp. R312菌株(Huang et al.,1997)晶体结构的活性位点口袋成功 模拟的支持。 根据这种模型已经提出了三种不同的催化机理。 所有机制都需要金属离子充当 路易斯酸作用激活腈的水合反应。由于不需要配体交换,一般认为,这种机制最可能是腈的 间接性激活。三价离子的配体交换十分缓慢,致使此机制很少受到欢迎。现已阐明[45]R. rhodochrous sp. J1菌株的钴型腈水合酶与Rhodococcus sp. R312菌株的铁型腈水合酶在相同 的反应条件下,水解丙腈的反应效率几乎一致。由于低自旋铁(III)和钴(III)的配体交换动力 学特征明显不同, 这两种酶类似反应性质暗示这些酶反应机制不需要配体的交换。 在由黄等 人推崇的机制中, 即所谓的外球体机制, 有一个氢氧离子充当一般的碱在活性位点激活水分 子,这样的水分子反过来攻击腈。已经报道嗜热菌株Pseudonocardia thermophila JCM 3095 的钴型腈水合酶晶体结构[46]。该结构(分辨率:1.8)显示与适温腈水合酶有很高的结构 相似性。 然而两种酶的明显区别在于P. thermophila腈水合酶的α和β亚基之间有附加的相互作 用, 这是由两个额外的螺旋引起的, 这表明这种额外的相互作用很可能有助于酶的热稳定性。 钴型腈水合酶的活性中心(金属结合位点)像铁型酶一样有相同的爪状模型,与钴离子配位 的配体是由cys108α , cys111α and cys113α残基的硫原子和ser112α and cys113 α的主链酰胺氮 原子之间形成的。电子密度峰指出cys111α and cys113α在转录后分别氧化成半胱次磺酸和半 胱亚磺酸,这已经确定无疑[47]。在Rhodococcus sp. N-771腈水合酶[48]中,由于来自这些修饰
废弃菌体水解液作氮源发酵产腈水合酶的研究

及生长因子. 目前 , 废弃菌体的水解方法多采用生物
法( 酶解法 、 自溶法 ) 、 物 理法 ( 超声 波 破碎 法 ) 、 化 学
法( 酸解 、 碱解 ) J . 然而这些方法要实现。 f 业 应 用
1 . 1 . 1 菌种
诺 卡 氏菌 ( N o c a r d i a s p . ) , 江 西 师范
第5 期
孙婉菊, 等: 废弃菌体水解液作氮源发酵产腈水合酶 的研究
5 3 1
0 . 5 0 0, 尿素 0 . 7 0 0 , 谷 氨酸 钠 0 . 1 0 0 , 磷 酸 二 氢 钾 0 . 0 5 0, 磷酸 氢二 钾 0 . 0 5 0 , 无水 硫 酸镁 0 . 0 2 0 , 氯 化
方法应 用 于工业 中 , 生 产 各 种 与 人类 生 活 息 息 相 关 的产 品. 微生物法与化学法相 比, 减 少 “ 三废 ” 的 排放 , 对环 境 的污染 小. 然而 , 随 着微 生物 法 在 上 业 上 的广泛 应用 , 发酵 生产 废弃 细胞 常常 直接 被排 放 , 造成 环境 污染及 资 源 浪 费 … , 工 业 废 弃 菌体 的处 理
或 一类 物质 , 不利 于 废 弃 菌体 的其 他 有 机 大分 子 的 水解. 因而, 无论从 经济 成 本 、 适 用 范 围和使 用 效 果 等 方 面考 虑 . 废 弃 菌 体 水 解 的 方 法 都 有 待 进 一 步
改进 .
视程度 的提高 , 微生物法越来越多地取代传统化学
壁条件 ’ F, 细胞破碎液 中氨基氮含量为 2 4 . 5 g・ L~. 并 考察 j ’ 酵母膏与水解液的不同配 比对腈水合酶 发 酵酶活 的影响 , 确定了酵母膏与废弃菌体水解液 的最佳 配比为 3 : 2 . 实验 结果表 明 : 利用热碱 法处理废 弃
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
消息热点(NEWS ANDHIGHLIGHTS)
腈水合酶制备关键技术研究
腈水合酶是一类可以催化腈类物质转化相应的酰胺类化合物的金属酶,工业上利用其催化丙烯腈转 化为丙烯酰胺。
该生物法制备丙烯酰胺工艺在1985年被开发,是生物法替代化学法的典型案例。
随着丙烯酰胺的需求不断扩大,国内外研究机构及学者对腈水合酶产生菌的分离选育、菌株特性研 究、酶的形成条件、酶的结构和性质、生物合成机制、翻译后的成熟机制等进行了深入研究。
如中国专利 CN201710456875.6公开了 一种改造腈水合酶及其应用,该发明改造腈水合酶的抗逆性、耐热性和产物耐受 性都有显著提升,并且没有导致腈水合酶活性的降低。
改造腈水合酶可催化获得高浓度丙烯酰胺产物,且 可以多批次回用。
CN201610035437.8为一种球化酶技术固定化腈水合酶的方法,该方法包括以下步骤:(1 &向腈水合酶溶液中加入保护剂PLL,混合均匀作为水相;另向正己坑中加入表面活性剂span-80,混合均匀 作为油相;再把水相和油相混合搅拌制成粗乳液;(2)将上步得到的粗乳液在5~30 k P a的跨膜压力下通过 SPG膜乳化器,待粗乳液完全通过SPG膜后,即得到W/0型(油包水)乳液,然后向该乳液中滴加交联剂戌 二醛,交联0.5~4 h,离心洗涤,即可得到球形腈水合酶粒子。
该发明的制备工艺简单,条件温和,酶活回收 率在50%以上。
重复使用10次后,酶活还可达到初始酶活的45%左右。
同游离酶相比,储藏稳定性也更好。
CN201510244547.0公开了一种高效表达高分子量型腈水合酶的基因工程菌及其应用,该发明所述的高分 子量型腈水合酶基因nhhB(rbs)A(rbs)<是基于R.rhodochrous J1来源的腈水合酶基因bag,经密码子优化及 表达元件改造之后化学合成得到,该基因全长1 645 bp,编码533个氨基酸,能够在大肠杆菌中成功表达。
该方法高效安全,在24 "诱导16 h即可获得96.7 U/mL可溶性腈水合酶,纯化后比酶活高达234 U/mg。
该 方法有利于生产过程中腈水合酶的提取纯化以及酰胺类物质的高效酶法合成。
CN201510226690.7公开了 一种比酶活和稳定性提高的融合型腈水合酶,其将来源于恶臭假单胞菌的腈水合酶在大肠杆菌中高效表达并采用亚基融合策略提高稳定性及产物耐受性。
该发明方法简单高效安全,能够在较短的表达周期里获 得稳定性高,产物耐受性强的大量可溶性腈水合酶,有利于在简单条件下进行大规模工业生产,以及进一 步对融合型腈水合酶成熟机制的理论研究。
CN201410843384.3公开了一种耐热重组腈水合酶基因、编码 酶、载体、工程菌及其在催化腈化合物生成酰胺和生物降解除草剂敌草腈中的应用,该发明实现了源自慢 生型大豆根瘤菌USDA 110中假定腈水合酶基因的功能表达,证明了一种腈水合酶的活化元件在表达过 程中的关键作用。
同时,该发明提供了一种在大肠杆菌内构建腈水合酶基因工程菌的方法,在腈水合酶基 因在活化元件的参与下获得具有高表达量和高活性的腈水合酶。
CN201410758150.9公开了 一种腈水合酶 及其应用,该腈水合酶由!亚基和"亚基组成。
该发明从博得特氏菌DSM 12804 (Bordetella petrii DSM 12804)中克隆到腈水合酶基因,该基因表达后成功获得具有高表达量和高活性而且底物谱广、具手性选择 性的腈水合酶。
CN201280028338.7提供催化活性提高了的改良型腈水合酶,并且提供编码该改良型腈水合 酶的DNA、包含该D N A的重组载体、包含该重组载体的转化体、从该转化体的培养物中提取的腈水合酶及 其制造方法。
CN201110384089.2公开了一种克隆表达腈水合酶调控蛋白的方法,是采用在腈水合酶调控蛋 白基因序列N端加His-tag,与腈水合酶成熟酶基因串联,在大肠杆菌B L21中共表达。
在现今微生物法生产丙烯酰胺的流程中,仍然普遍存在菌种的酶活稳定性差、对底物和产物的耐受性 不高以及产物丙烯酰胺会向副产物丙烯酸进一步转化等问题。
为此,利用基因工程手段研究腈水合酶在氨 基酸序列、折叠方式和活性位点等方面的特性,并通过代谢工程优化水合反应的代谢过程,将成为今后微 生物法生产丙烯酰胺的研究热点之一。
(江南大学图书馆张群)
JOURNAL OF FOOD SCIENCE AND BIOTECHNOLOGY Vol.36 No.12 2017 |。