《现代分子生物学》第五章 3 真核生物的转录后加工
分子生物学习题集答案 第五章

第五章基因表达自测题(一)选择题A型题1. 基因表达包括①复制②转录③逆转录④翻译A. ①+②B. ①+③C. ②+④D. ③+④E. ①+②+④2. 真核生物的转录不.需要A. 依赖DNA的RNA聚合酶B.三磷酸核苷C.DNAD.RNAE.蛋白质3. 关于转录和复制的区别,说法正确的是A. DNA双链均复制和转录B.复制的原料是一磷酸脱氧核苷,转录的原料是一磷酸核苷C.复制需要引物,转录不需要引物D.复制酶是依赖DNA的DNA聚合酶,转录酶是依赖RNA的DNA聚合酶E.均遵守碱基配对规律,模板中A对应的产物是T4. 关于转录和复制的相同点,说法错误..的是A. 核苷酸以3', 5'-磷酸二酯键连接B.以DNA为模板C.原料为dNTP或NTP,组成核酸链的是dNMP或NMPD.产物可与变性的模板杂交E.新生核酸链的延伸方向是3' → 5'5. 已知某基因转录产物的部分序列是5'-AUCCUGGAU-3',那么该基因中反意义链的相应序列为A.5'-ATCCAGGAT-3'B.5'-TAGGTCCTA -3'C.5'-TAGGACCTA-3'D.5'-UAGGACCUA-3'E.5'-ATCCUGGAT-3'6. E.coli的RNA聚合酶中,辨认转录起始点的组分是A. 核心酶B. σC. αD. βE. β'7. 真核生物中,RNA聚合酶II的转录产物是A. 45S rRNAB. 5S rRNAC. tRNAD. U6 snRNAE. hnRNA8. 关于真核生物的RNA聚合酶,说法正确的是A.RNA聚合酶II对鹅膏蕈碱极敏感B.RNA聚合酶I的转录产物往往是小分子RNAC.RNA聚合酶的氨基端重复序列可发生磷酸化D.不一定需要蛋白质因子辅助E.RNA聚合酶III的转录产物是45S rRNA9. 关于E.coli RNA聚合酶的σ因子的说法错误..的是A.不参与转录延长过程B.辨认–35区和–10区起始转录C.σ因子若不脱落,转录不能进入延长阶段D.σ因子的量少于核心酶,可反复利用E.转录开始后,补加σ因子可提高RNA的生成量10. 在某个体外转录体系中,若模板链含有连续的A,加入何种物质可对产物进行放射性标记A. α-32P-UMPB.γ-32P-dTTPC. β-32P-UDPD. α-32P-UTPE. γ-32P-UTP11. 下列哪项不.是ρ因子依赖的转录终止的特点A.转录终止信号存在于RNA而非DNA模板B.ρ因子有ATP酶活性和解螺旋酶活性C.ρ因子结合RNA的能力弱于对DNA的结合力D.生成的RNA链的3' 端往往富含CE.ρ因子通过与转录产物结合而发挥转录终止作用12. 真核生物II类基因的启动子核心序列通常位于A. –25区B. –10区C. –35区D. +1区E. +10区13. 真核生物中,与原核生物的Pribnow box功能相当的是A. GC boxB. CAAT boxC. CACA boxD. 八聚体(8 bp)元件E. Hogness box14. 下列物质中,能够辅助真核生物的RNA聚合酶结合启动子的是A. 起始因子B. 增强子C. 转录因子D. 延长因子E. σ因子15. 真核生物的RNA聚合酶II在进入转录延长阶段之前,其最大亚基羧基末端结构域发生磷酸化是必需的,催化这一步骤的转录因子是A. TF II BB. TF II DC. TF II HD. TF II EE. TF II F16. 关于转录因子的说法,错误..的是A. 原核细胞中转录起始复合物的形成不需要转录因子B.转录因子的化学本质是蛋白质C.转录因子通过DNA-蛋白质、蛋白质-蛋白质相互作用控制转录起始D.转录因子的转录激活域包括锌指、螺旋-转角-螺旋等结构域E.少量转录因子的不同排列组合即可精确调控众多基因的转录17. 关于真核生物的转录延长,说法正确的是A. 组蛋白发生磷酸化利于核小体解聚B.组蛋白发生乙酰化利于核小体解聚C.转录延长的同时出现多聚核蛋白体现象D.延长因子辅助RNA聚合酶催化核苷酸加入E.转录前后,核小体解聚并在原位重新聚合18. 下列哪种物质不.需要进行转录后加工即可发挥功能A. 大肠杆菌 mRNAB. 大肠杆菌tRNAC.大肠杆菌rRNAD.酵母mRNAE.酵母tRNA19. 关于加帽修饰,说法正确的是A. snRNA不能被加帽B.由加帽酶催化5' 端加入7甲基鸟苷酸C.加帽酶可以与RNA聚合酶I、II、III相结合D.hnRNA转录终止后才开始加帽E.RNA聚合酶发生磷酸化后,使加帽酶与之脱离20. 关于加尾修饰,说法错误..的是A. 组蛋白的成熟mRNA无需加poly A尾B.加尾信号包括AAUAAA和富含GU的序列C.剪切过程需要多种蛋白质因子的辅助D.加尾不需模板E.多聚A形成的过程是缓慢、匀速的反应21. 关于剪接的说法,正确的是A. 初级转录物中不存在内含子B.内含子一定是非编码序列C.内含子两端的结构通常是5'-GU-----AG-3'D. snRNP催化套索RNA发生转酯反应,使内含子和剪接接口一起被去除E.剪接后的mRNA不能与模板链DNA杂交22. RNA编辑发生在A. 成熟的mRNAB. tRNA和rRNA的前体C. hnRNAD.成熟的tRNA和rRNAE. snRNA23. 下列哪种情况下,DNA双链贮存的遗传信息没有..改变A. RNA编辑B.转换C.错义突变D.倒位E.移码突变24. 关于真核rRNA的转录以及加工,说法正确的是A. rDNA位于核仁,呈串联重复排列B.转录出的5S rRNA组成多顺反子RNA的一部分C.rDNA之间被可转录的基因间隔所分隔D.基因间隔等同于内含子E.45S rRNA前身可剪接成为16S、5.8S和28S rRNA25. 生命活动涉及核酸和蛋白质两类生物大分子之间的相互作用,下列物质中不.属于核酸-蛋白质复合物的是A. telomereB.splicesomeC.ribozymeD.SRPE.mRNP26. 蛋白质的生物合成不直接...需要A. RNAB. RNA剪切因子C.分子伴侣D.帽子结合蛋白E.GTP27. mRNA在蛋白质合成中的功能是A. 运输遗传密码所对应的氨基酸B.与蛋白质结合,提供合成场所C.与帽子结合蛋白结合启动翻译D.由三联体密码指引氨基酸的排列顺序E.通过剪切因子切除poly A尾调控翻译效率28. 某种情况下,基因的点突变可能不会影响它所编码的蛋白质的一级结构,遗传密码的哪种特点对此发挥了重要作用A. 方向性B. 连续性C. 摆动性D. 通用性E. 简并性29. 关于遗传密码说法错误..的是A. 遗传密码的阅读方向是5'→3'B. 细胞核和线粒体的遗传密码完全通用C. 61组有意义密码分别编码20种氨基酸D. 密码子的第三位的作用没有前两位重要E. 不同生物对密码子有不同的偏爱30. 密码子与反密码子之间存在摆动配对现象,如果密码子为CGU,则识别它的反密码子可能是A. ICGB. GCAC. CCGD. UCGE. GCI31. 原核生物的核糖体大亚基是A. 30SB. 40SC. 50SD. 60SE. 70S32. 真核生物的核糖体不.含有A. E位B. P位C. A位D. 5S rRNAE. 转肽酶33. 氨基酰-tRNA合成酶的特点是A. 能水解磷酸二酯键,发挥校正功能B. 对氨基酸有绝对专一性C. 对tRNA有相对专一性D. 对氨基酸和tRNA都有高度特异性E. 能水解肽键,发挥校正功能34. 活化后的氨基酸才能与tRNA结合,每个氨基酸的活化过程需要消耗几个高能磷酸键A. 1B. 2C. 3D. 4E. 535. 关于起始tRNA说法错误..的是A. 在肽链延长阶段,起始tRNA不能继续运输蛋氨酸B. 原核与真核起始tRNA所携带的氨基酸不同C. 真核生物中识别AUG的tRNA至少有两种D. 原核生物的起始tRNA可以辨认AUG、GUG和UUGE. 原核生物的起始tRNA可以运输N-甲酰甲硫氨酸、缬氨酸和亮氨酸36. 原核生物的70S 翻译起始复合物不.包括A. 50S亚基B. IF-2C. fMet-tRNAD. mRNAE. 30S 亚基37. 真核生物参与蛋白质合成的起始因子有几种A. 1B. 2C. 3D. 4E. > 538. 原核生物的翻译起始阶段,帮助fMet-tRNA结合AUG的是A. IF-2B. IF-1C. eIF-2D. eIF-3E. eIF-439. SD序列与下列哪种rRNA相互作用A. 5SB. 23SC. 16SD. 5.8SE. 18S40. 原核生物的翻译过程中,能进入P位的是A.fMet-tRNAB.Met-tRNAC.Val-tRNAD. Leu-tRNAE. Ile-tRNA41. 关于真核生物的翻译起始,不.正确的说法是A. 翻译起始的过程较原核生物复杂B. eIF-2是调节真核翻译起始的关键因子C. 起始氨基酰-tRNA进入核糖体的P位D. mRNA的SD序列与其在核糖体上就位有关Met组成翻译起始复合物E. 由80S核糖体、mRNA和Met-tRNAi42. 真核生物肽链合成的延长阶段,促使氨基酰-tRNA进入A位的蛋白质因子是A. EF-1B. EF-2C. EFTD. EFGE. 转肽酶43. 关于转肽酶的说法正确的是A. 是rRNA的一部分B. 由数种核糖体蛋白组成C. 成肽反应不需要延长因子D. 催化氨基酸的α-羧基与核苷酸的3'-OH形成酯键E. 催化新生的肽酰-tRNA转移到P位44. 狭义的真核生物核蛋白体循环所涉及的蛋白质因子是① IF ②转肽酶③ EF ④ RFA. ①+②+③+④B. ①+③+④C. ②+③+④D. ①+③E. ②+③45. 真核生物中参与翻译终止的释放因子的数目是A. 1B. 2C. 3D.4E. > 546. 蛋白质合成时消耗能量的步骤发生在①氨基酸活化②进位③成肽④转位⑤新生肽链的释放A. ①+③+⑤B. ①+②+③C. ①+④+⑤D. ①+②+④E. ①+②+④+⑤47. 蛋白质合成时,每增加1个肽键至少需要消耗几个高能磷酸键A. 1B. 2C. 3D. 4E. 548. 具有酯酶活性并能释放新生肽链的蛋白质因子是A. RF-3B. RRC. 转肽酶D. 转位酶E. IF49. 翻译后加工不.包括A. 由分子伴侣辅助折叠成空间构象B. 新生多肽链从tRNA上水解释放C. 亚基聚合D. 氨基酸残基的共价修饰E. 辅基的结合50. 关于分子伴侣的叙述,错误..的是A. 识别并结合非天然构象的蛋白质B. 自身是一类较为保守的蛋白质C. 热休克蛋白是常见的分子伴侣D. 广泛存在于原核和真核细胞E. 其作用不依赖于蛋白质一级结构的信息51. 新生多肽链中,不.发生共价修饰的氨基酸残基是A. CysB. ThrC. MetD. ProE. Lys52. 下列哪项不.是信号肽的特点A. N端一小段富含亲水氨基酸的序列B. 与细胞质中的信号识别颗粒结合C. SRP受体是蛋白进入内质网所必须的D. N端一小段富含疏水氨基酸的序列E. 翻译的同时引导新生多肽链穿过内质网膜53. 引导新生多肽链靶向输送到细胞核的核酸序列称为A. signal peptideB. NLSC. KDELD. MLSE. SD54. 采用翻译后转运机制的蛋白质是A. 免疫球蛋白B. 肽类激素C. 核DNA编码的线粒体蛋白D. 凝血因子E. 水解酶55. 哪一项不.属于基因表达的范畴A. mRNA模板指导的蛋白质合成B. DNA模板指导的hnRNA合成C. DNA模板指导的DNA合成D. DNA模板指导的rRNA合成E. DNA模板指导的snRNA合成56. 下列哪一项不.是转录的原料A. TTPB. ATPC. CTPD. UTPE. GTP57. 转录生成的RNA链中存在A. dAMPC. UDPD. dTTPE. UMP58. 初级转录产物是A. 以转录调控序列为模板转录的序列B. 以结构基因为模板转录的全部序列C. 以外显子为模板转录的序列D. 以内含子为模板转录的序列E. 以上都不对59. 断裂基因的转录过程是A. 不对称、不连续B. 对称、不连续C. 对称、连续D. 不对称、连续E. 以上都不对60. 在复制和转录中均起作用的是A. RNA引物B. DNA聚合酶D. dNTPE. 蛋白质因子61. 比较复制和转录的聚合酶,错误..的说法是A. 底物不同B. 催化聚合反应的方向不同C. 产物不同D. 碱基配对略有不同E. 模板链不同62. 以DNA为模板合成RNA的过程称为A. DNA replicationB. transcriptionC. reverse transcriptionD. translationE. RNA replication63. 转录时模板与产物之间不.存在的碱基对应关系是A. A→TB. U→AC. A→UD. C→GE. G→C64. RNA转录合成需要的酶是A. DNA-dependent DNA polymeraseB. DNA-dependent RNA polymeraseC. RNA-dependent RNA polymeraseD. RNA-dependent DNA polymeraseE. Taq DNA polymerase65. 真核生物的RNA聚合酶有几种A. 1B. 2C. 3D. 4E. 566. 原核生物的RNA聚合酶有几种A. 1B. 2C. 3D. 4E. 567. E.coli RNA聚合酶的核心酶含有的亚基是A. α、β、β'、σB. α、β、σ 、ωC. α、β'、σ 、ωD. α、β、β' 、ωE. β、β'、σ 、ω68. 大肠杆菌的转录全过程始终需要A. αβ2β'ωB. α2ββ'ωC. αββ'2ωD. α2β2β'ωE. αββ'ω69. E.coli RNA聚合酶的全酶与核心酶的差别在于A. α亚基B. β亚基C. β'亚基D. σ亚基E. 以上都包括70. 真核生物中催化合成tRNA的酶是ββ'A. α2ββ'σB. α2C. RNA polymerase ID. RNA polymerase IIE. RNA polymerase III71. 遗传密码的简并性是指A. 一种三联体密码编码一种氨基酸B. 一种三联体密码编码几种氨基酸C. 几种三联体密码编码同一种氨基酸D. 不同的三联体密码编码不同的氨基酸E. 三联体密码中第一位碱基的差异不影响编码氨基酸的特异性72. 关于各种原核生物RNA聚合酶的σ亚基,描述错误..的是A. 种类很多B. 分子量不同C. 参与转录延长过程D. 与转录起始点相结合E. 决定转录的特异性73. 原核生物RNA聚合酶的σ亚基辨认转录起始点的实质是A. DNA-蛋白质相互作用B. RNA-蛋白质相互作用C. 蛋白质-蛋白质相互作用D. DNA-DNA相互作用E. DNA-RNA相互作用74. 下列哪一项不.属于核酸A. 转录因子B. 操纵子C. 转录子D. 复制子E. 顺反子75. 下列位置关系叙述正确的是A. 启动子从不会位于转录起始点下游B. 增强子全部位于结构基因内部C. 大部分TATA盒位于转录起始点下游D. 转录调控序列全部位于基因之外E. 翻译起始密码子位于转录起始点下游76. 不.需要引物的酶是A. Klenow片段B. RNA聚合酶C. 反转录酶D. Taq DNA聚合酶E. DNA-pol δ77. 真核生物的RNA聚合酶发生哪种修饰后,进入转录延长阶段A. 磷酸化B. 去磷酸化C. 甲基化D. 去甲基化E. 乙酰化78. 大肠杆菌中催化mRNA链延长的酶是A. Klenow fragmentB. αββ'σ ω2C. DNA-pol IIIββ'ωD. α2E. RNA-pol II79. 转录起始生成RNA的第一位核苷酸最常见的是A. ATPB. AMPC. GTPD. GMPE. CMP80. 真核生物合成mRNA的起始阶段不.涉及A. Hogness boxB. release factorC. transcriptional factorD. RNA-pol IIE. cis-acting element81. 狭义的核糖体循环是指A. 翻译起始、延长和终止的全过程B. 翻译的起始过程C. 翻译延长的进位、成肽和转位过程D. 翻译的终止过程E. 翻译与翻译后加工的全过程82. 关于选择性剪接,正确的说法是A. 选择性转录内含子B. 选择性转录外显子C. 初级转录产物相同,后加工过程不同D. 成熟mRNA相同,翻译过程不同E. 初级转录产物的序列发生改变83. RNA编辑发生在哪一个层次A. 复制水平B. 转录水平C. 转录后水平D. 翻译水平E. 翻译后水平84. 真核生物中编码哪种蛋白质的mRNA不.具有poly (A) 尾A. 细胞色素CB. 胰岛素C. 角蛋白D. 血红蛋白E. 组蛋白85. 下列哪一项不.是蛋白质的分拣信号A. SD sequenceB. signal peptideC. MLSD. KDELE. NLS86. 在转录后加工过程中可以观察到A. 套索RNAB. 羽毛状结构C. 锤头状RNAD. 叉状结构E. 转录空泡87. 下列核酸-蛋白质复合物中,哪一项与真核生物的RNA合成无关..A. ribosomeB. hnRNPC. snRNPD. splicesomeE. mRNP88. 以mRNA为模板合成蛋白质的过程称为A. DNA replicationB. RNA replicationC. transcriptionD. reverse transcriptionE. translation89. 关于多聚核蛋白体的描述,正确的是A. 结合了核蛋白体的几条mRNA链聚在一起B. 一个核蛋白体连续翻译多肽链C. 一条mRNA链上结合多个核蛋白体D. mRNA、核蛋白体和多肽链的复合体E. DNA、mRNA、核蛋白体和多肽链的复合体90. 电子显微镜下观察到原核生物的基因表达呈现羽毛状结构,其根本原因在于A. 多聚核蛋白体形成B. 多个转录过程同时进行C. 多个翻译过程同时进行D. 转录与翻译偶联进行E. 新生的多肽链不脱落91. 原核生物mRNA中的哪种结构与核糖体相结合A. AUGB. SD序列C. ORFD. 5' 端非翻译区E. 3' 端非翻译区92. 原核生物的翻译起始阶段,mRNA中的SD序列与哪段序列互补结合A. 23S rRNA的5' 端序列B. 16S rRNA的5' 端序列C. 5S rRNA的5' 端序列D. 23S rRNA的3' 端序列E. 16S rRNA的3' 端序列93. 参与蛋白质合成的蛋白质因子不.包括A. 起始因子B. 延长因子C. 释放因子D. 转肽酶E. 反式作用因子94. 密码子与反密码子之间有时不严格遵守碱基配对规律,这是由遗传密码的哪种性质决定的A. 方向性B. 连续性C. 通用性D. 简并性E. 摆动性95. 遗传密码具有通用性,但下列哪一项例外..A. 大肠杆菌的基因组DNAB. 人的线粒体基因组C. 鲸鱼的核DNAD. 线虫的染色体基因组E. 小麦的染色体DNA96. 不.属于直接进入蛋白质合成的氨基酸是A. 羟赖氨酸B. 精氨酸C. 谷氨酰胺D. 瓜氨酸E. 胱氨酸97. 在翻译起始阶段发挥作用的蛋白质因子是A. IFB. EFC. RFD. RRE. 转肽酶98. 翻译的实质是A. 将4种脱氧核糖核苷酸的排列顺序转换为4种核糖核苷酸的排列顺序B. 将4种核糖核苷酸的排列顺序转换为4种脱氧核糖核苷酸的排列顺序C. 将4种脱氧核糖核苷酸的排列顺序转换为20种氨基酸的排列顺序D. 将4种核糖核苷酸的排列顺序转换为20种氨基酸的排列顺序E. 将蛋白质一级结构的信息转换为空间结构的信息99. 关于多顺反子mRNA,正确的说法是A. 几个mRNA分子有不同的开放阅读框B. 几个结构基因由不同的启动子调控转录C. 一个mRNA分子有几个开放阅读框D. 多个结构基因编码一类蛋白质E. 一个结构基因编码多种蛋白质100. 抑制真核生物蛋白质合成的抗生素是A. 放线菌酮B. 氯霉素C. 四环素D. 新霉素E. 链霉素B型题A. σB. αC. βD. β'E. CTD101. 原核生物的RNA聚合酶中,辨认转录起始点的组分是102. 原核生物的RNA聚合酶中,决定转录速度的亚基是103. 真核生物的RNA聚合酶的结构组成部分A.IFB.σ因子C.snRNPD.ρ因子E.TFII104. 与原核生物转录终止相关的是105.真核生物mRNA转录起始需要106. 真核生物mRNA剪接加工需要107. 原核生物的翻译起始需要A. 胱氨酸B. 谷氨酸C. 鸟氨酸D. 脯氨酸E. 甲硫氨酸108. 不属于...构成蛋白质的氨基酸是109. 能组成蛋白质但没有遗传密码的氨基酸是110. 能作为起始氨基酸的是111. 属于酸性氨基酸的是112. 属于亚氨基酸的是A. GTPB. UMPC. dTTPD. dAMPE. CDP113. 复制的原料包括114. 转录的原料包括115. 复制产物中包含116. 转录产物中包含117. 为翻译过程提供能量的核苷酸是A. U6 snRNAB. mRNAC. tRNAD. 5S rRNAE. 45S rRNA118. RNA聚合酶I催化合成119. RNA聚合酶II催化合成A. Taq DNA polymeraseB. Klenow fragmentC. RNA polymerase IIIββ'ωD. α2E. αββ'σ2120. 真核生物合成tRNA的过程依赖121. 原核生物合成mRNA的转录延长阶段需要122. 原核生物合成rRNA的转录起始阶段需要123. 体外催化DNA合成并能耐受高温的酶是A. 核糖体小亚基B. 核糖体大亚基C. 起始因子D. 起始氨基酰-tRNAE. mRNA124. 30S翻译起始复合物不.包括125. 70S翻译起始复合物不.包括X型题126. 基因表达的最终产物包括A.蛋白质B.mRNAC.tRNAD.rRNAE. 其他小分子RNA127. 转录的特点是A. 细胞内并非所有的基因都转录B. 不对称转录C. 特异性转录因子决定转录的时序性D. 模板可以分布在DNA的两条链上E. 不同单链上的模板转录方向一致128. 与模板链同义的是A. 正链B. 负链C. 有意义链D. 反意义链E. 编码链129. 原核生物的RNA聚合酶在转录延长过程中不.脱落的组分是A. σB. αC. βD. β'E.核心酶130. 可被RNA聚合酶保护的核酸区域是A. 5' 端帽子B. 顺反子C. Pribnow boxD. 羧基末端结构域E. –35区131. 转录生成的RNA链中,5′端第一个核苷酸通常是A. ATPB. CTPC. GTPD. CMPE. GMP132. 原核生物和真核生物的RNA聚合酶的区别在于A. 能否辨认Pribnow box或者TATA boxB.能否催化生成5' 端的四磷酸二核苷酸结构C.与模板结合需否辅助因子D.能否生成转录起始复合物E.是否对利福平敏感133. 真核生物细胞核内进行的mRNA转录后加工包括A. 5' 端加帽B.3' 端加尾C.修饰生成稀有碱基D.选择性剪接E.RNA编辑134. 基因组序列测定的研究结果表明,真核生物的基因数量远远少于预期值,你认为机体如何满足编码多种蛋白质的生理需要A. 自发突变B.多蛋白的水解修饰C.染色体易位D. RNA编辑E.选择性剪接135. 选择性剪接的作用机制包括A. 使用不同的剪接位点B.选择性使用外显子C.反式剪接D.使用不同的启动子E.使用不同的多腺苷酸化位点136. 不.属于有意义密码的是A. UGAB. UAAC. AUGD. UAGE. GUA137. 当mRNA中密码子的第三位是碱基C时,tRNA中反密码子的第一位碱基可能为A. IB. GC. UD. AE. C138. 30S翻译起始复合物和70S翻译起始复合物含有的相同组分是Met E. IF A. 50S亚基 B. 30S亚基 C. mRNA D. fMet-tRNAf139. 在真核生物的翻译起始阶段,参与大小亚基拆分的蛋白质因子是A. eIF-2B. eIF-3C. eIF-4ED. eIF-6E. eIF-4F 140. 在肽链合成的延长阶段,真核生物与原核生物的区别在于A. 反应体系B. 延长因子C. 卸载tRNA的脱落方式D. 肽链延伸的方向E. 消耗GTP141. 能占据核糖体A位的是A. fMet-tRNAB. IF-1C. eRFD. RF-3E. Val-tRNA 142. 蛋白质的生物合成过程具有高保真性,可能的机制包括A. 密码子与反密码子的碱基配对B. 氨基酰-tRNA合成酶的专一性和校正功能C. EF-GTP的存在时间极短,防止错误进位D. 耗能水解清除任何步骤出现的不正确连接E. mRNA和氨基酰-tRNA在核糖体中的正确就位(二)名词解释1. 基因表达(gene expression)2. 不对称转录(asymmetric transcription)3. 翻译(translation)4. TATA盒(TATA box)5. 套索RNA(lariat RNA)6. 选择性剪接(alternative splicing)7. RNA编辑(RNA editing)8. 密码子的简并性(degeneracy)9. 密码子的摆动性(wobble)10. SD序列(Shine-Dalgarno sequence)11. 多聚核糖体(polyribosome)12. 核糖体循环(ribosome cycle)13. 分子伴侣(molecular chaperon)14. 信号肽(signal peptide)(三)简答题1. 试比较DNA复制和转录的异同。
转录后加工详解
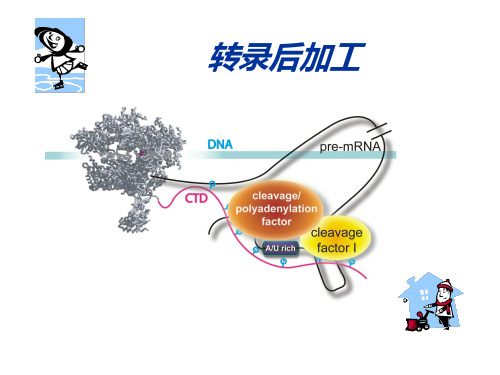
转录后加工
基因转录的直接产物被称为初级转录物。初 级转录物一般是无功能的,它们在细胞内必 须经历一些结构和化→学的变化即所谓的转录 后加工以后才会有功能。转录后加工可能是 RNA的功能所必需的,也可能提供基因表达 调控的一种手段。
RNA所能经历的后加工方式可达10种以上, 但后加工反应的本质要么是增减一些核苷酸 序列,要么是修饰某些特定的核苷酸。
AAUAAA
mRNA
* mRNA前体在AAUAAA序列下游10-30个核苷酸的位 置被CFI/II切开 * 产生的3′-OH被PAP作为添加腺苷酸的位点。PAP不 需要模板,只对ATP有亲和性 * Poly(A)尾巴被 poly(A)-结合蛋白结合(PABP)
AAUAAA
AAAAAAAAAAAAA100-200
* 后加工机制
加帽和甲基化
* 帽子结构和类型 0、I和 II型
* 加帽反应:共转录 1) 酶 磷酸水解酶;mRNA鸟苷酸转移酶;鸟嘌呤-7-甲基转移 酶 2) 步骤
* 为什么只有mRNA才会加帽? * 功能
加帽反应
开始于mRNA 5′-三磷酸的 γ 磷酸根的水解
-
-
g ba
O O O 5’
O-P-O-P-O-P-OCH2 碱基
为什么只有mRNA才会加尾?
* 与加帽反应一样,只有mRNA才会加 尾,也是因为聚合酶II最大亚基上的 CTD重复序列被TFIIH磷酸化,但是 磷酸化位点为Ser2。Ser2的磷酸化将 加尾因子招募到mRNA 前体上进行加 尾反应。
mRNA前体的剪接
剪接这种后加工方式是在发现基因断裂的现象后确定 的。1977年,由Phillip Sharp和Richard Roberts领导两个 实验小组几乎同时在腺病毒的晚期表达基因中发现蛋 白质基因断裂现象。
大学《现代分子生物学(第3版)考试重点

的空隙。
聚合酶Ⅱ:修复紫外光引起的 DNA 损伤
聚合酶Ⅲ:DNA 复制的主要聚合酶,还具有 3’-5’外切酶的校对功能,提高 DNA 复制
的保真性
真核生物中的 DNA 聚合酶
定位
α 细胞核
β 细胞核
γ 线粒体
δ 细胞核
ε 细胞核
3‘-5’外切酶活性 -
-
+
+
+
功能
引物合成 修复作用 线粒体 DNA 的复制 核 DNA 的复制 RNA 引
11、 tRNA 中起作用的重要两个臂是什么臂?
tRNA 有两个关键部位: ● 3’端 CCA:接受氨基酸,形成氨酰-tRNA。 ● 与 mRNA 结合部位—反密码子部位
12、.肽链延伸由许多循环组成,每加一个氨基酸就是一个循环,每个循环包括 哪些步骤?
肽链延伸由许多循环组成,每加一个氨基酸就是一个循环,每个循环包括:AA-tRNA 与核
备接受新的 AA-tRNA,开始下一轮合成反应
(3)移位:核糖体通过 EF-G 介导的 GTP 水解提供的能量向 mRNA 模板的 3‘端移动一
个密码子,使二肽酰—tRNA2 完全进入 P 位,准备开始新一轮的肽链延伸。
13、真核生物和原核生物在翻译的起始过程有哪些区别?
原核生物
真核生物
起始氨基酸
甲酰甲硫氨酸
(4)原核生物常以 AUG(有时 GUG,甚至 UUG)作为起始密码子
真核生物 mRNA 的特征:(1) 5’ 端存在“帽子”结构
(2)多数 mRNA 3’ 端具有 poly(A )尾巴(组蛋白除外)
(3)以单顺反子存在
(4)而真核生物几乎永远以 AUG 作为起始密码子。
生物化学转录后加工

真核细胞tRNA加工过程
真核生物RNA的转录后加工
❖真核生物rRNA前体的加工 ❖真核生物tRNA前体的转录后加工 ❖真核生物mRNA前体的加工
Figure 6-21 Molecular Biology of the Cell (© Garland Science 2008)
原核生物rRNA前体的加工过程
初转录产物内部碱基互补配对, 折叠成一些茎环结构
continue
茎环结构有助于一些蛋白质结合形成核糖核 蛋白复合体(RNP, ribonucleoprotein)
(甲基化)
(M16)
(RNaseIII和RNaseE酶的剪切)
(M23) (M5)
原核生物RNA的转录后加工
分或全部poly(A),此时mRNA的寿命亦接近终点。
加尾反应所需的特定序列元件
特定序列元件:1)5‘-AAUAAA-3’’ 聚腺苷酸信号序列; 2)在其后11~20nt处紧随着一个“ 5‘-YA-3’ ”结构; 3)在其下游方向的GUGUGUG 。这些序列共同决定了聚 腺苷酸化位点(polyadenylation site)。
真核生物mRNA poly (A)长度并非固定不变 细胞核中的poly (A)长度平均为210±20 nt,细胞质poly (A )长度
190±20 nt。 输送到细胞质中的mRNA其poly (A)可由RNase切短,但又能经细胞质
poly(A)酶重新加长,保持有限的长度。 细胞质中mRNA的poly (A)长度总的趋势是逐渐变短,直到丢失大部
真核生物rRNA的特点
真核生物有4种rRNA,即5.8S rRNA、18S rRNA、 28S rRNA和5S rRNA。其中前三者的基因组成一个转录单位, 由RNA pol Ⅰ合成,产生47S的前体,并很快转变成45S前 体;真核生物的5S rRNA由RNA pol Ⅲ合成121nt的转录产 物,几乎不需要加工。
现代分子生物学(第3版)_课后答案-五章

第一章 绪论1, 简述孟德尔、摩尔根和沃森等人对分子生物学发展的主要贡献。
答:孟德尔的对分子生物学的发展的主要贡献在于他通过豌豆实验,发现了遗传规律、分离规律及自由组合规律;摩尔根的主要贡献在于发现染色体的遗传机制,创立染色体遗传理论,成为现代实验生物学奠基人;沃森和克里克在1953年提出DAN 反向双平行双螺旋模型。
反向双平行双螺旋模型。
2, 写出DNA 和RNA 的英文全称。
答:脱氧核糖核酸(答:脱氧核糖核酸(DNA, Deoxyribonucleic acid DNA, Deoxyribonucleic acid DNA, Deoxyribonucleic acid)), 核糖核酸(核糖核酸(RNA, Ribonucleic acid RNA, Ribonucleic acid RNA, Ribonucleic acid))3, 试述“有其父必有其子”的生物学本质。
答:其生物学本质是基因遗传。
子代的性质由遗传所得的基因决定,而基因由于遗传的作用,其基因的一半来自于父方,一般来自于母方。
自于父方,一般来自于母方。
4, 早期主要有哪些实验证实DNA 是遗传物质?写出这些实验的主要步骤。
答:一,肺炎双球菌感染实验,答:一,肺炎双球菌感染实验,11,R 型菌落粗糙,菌体无多糖荚膜,无毒,注入小鼠体内后,小鼠不死亡。
型菌落粗糙,菌体无多糖荚膜,无毒,注入小鼠体内后,小鼠不死亡。
22,S 型菌落光滑,菌体有多糖荚膜,有毒,注入到小鼠体内可以使小鼠患病死亡。
型菌落光滑,菌体有多糖荚膜,有毒,注入到小鼠体内可以使小鼠患病死亡。
33,用加热的方法杀死S 型细菌后注入到小鼠体内,小鼠不死亡;后注入到小鼠体内,小鼠不死亡;二,噬菌体侵染细菌的实验:二,噬菌体侵染细菌的实验:11,噬菌体侵染细菌的实验过程:吸附→侵入→复制→组装→释放。
,噬菌体侵染细菌的实验过程:吸附→侵入→复制→组装→释放。
2 2 2,,DNA 中P 的含量多,蛋白质中P 的含量少;蛋白质中有S 而DNA 中没有S ,所以用放射性同位素35S 标记一部分噬菌体的蛋白质,用放射性同位素32P 标记另一部分噬菌体的DNA DNA。
分子生物学-转录后加工

小鼠免疫球蛋白μ重链基因可变剪接
14-14
可变剪接的多种模式
• 转录物可以按不同的模式进行可变剪接,产生具有多样性的转录本, 许多基因有2种以上的剪接方式,有的甚至达上千种。
• 几种常见的选择性剪接模式: • 1)不同启动子;2)忽略外显子;3)5’选择性剪接;4)3’选择性 剪接;5)保留内含子;6)多腺苷酸化
• 如果装配在snRNP的剪接因 子识别外显子,则称为外显子 界定(exon definition)
• 如果装配在snRNP的剪接因 子识别内含子,则称为内含子 界定(intron definition)
• 通过外显子-内含子边界(剪 接位点)突变,可以分析外显 子-内含子的界定类型
14-12
RNA Pol II的CTD参与外显子界定
内含子类型
特征
剪接体内含子(spliceosomal introns)
细胞核,mRNA,由剪接体催化切除
tRNA内含子(tRNA introns)
细胞核或古菌tRNA基因,由蛋白催化切除
自切割I类内含子(self-splicing group I introns) 细胞器,由RNA催化切除
自切割II类内含子(self-splicing group II introns)细胞器,由RNA催化切除
• 如果在6个位点发生2个不同事件,将会产生26 = 64种结果
14-15
可变剪接示例:果蝇性别决定
• 果蝇的性别决定涉及sxl(sex lethal,性别致死)、 tra (transformer, 转换)和dsx(doublesex, 双性别)三个基因前体mRNA的可变剪接
• 这3个基因的剪接存在级联反应:sxl基因雌性特异性剪接可产生活性 蛋白,进一步增强sxl基因的雌性特异性剪接,同时引发tra基因的雌性 特异性剪接,再进一步引发dsx的雌性特异性剪接
5第五章现代分子生物学研究方法——DNA、RNA及蛋白质操作技术

DNA的基本操作技术——核酸凝胶电泳 以琼脂糖凝胶电泳为例:
DNA的基本操作技术——核酸凝胶电泳
凝胶浓度的高低影响凝胶介质孔隙的大小,浓度越高,孔隙越小, 其分辨能力就越强。反之,浓度降低,孔隙就增大,其其分辨能力 就越弱。
DNA的基本操作技术——核酸凝胶电泳
溴化乙锭(ethidium bromide,EB)能插入到DNA或RNA分子的相 邻碱基之间,并在紫外灯光照射下发出荧光,所以常用EB来检测凝 胶介质中的核酸条带。
x 25 中的聚合酶可能很多正处于复制状态,
如果此时降到室温,将会影响最终产 率。所以再留出5分钟,以使正在复制 中的DNA能够复制完全,以合成更多 的目的分子。
DNA的基本操作技术——重组载体构建 PCR完成之后,需要有什么操作?
DNA的基本操作技术——重组载体构建
DNA的基本操作技术——重组载体构建
转化(transformation):是指重组质粒DNA分子通过与膜蛋白结合 进入受体细胞(一般指细菌),并在受体细胞内稳定维持与表达的 过程。
转染(transfection):是真核细胞主动或被动导入外源DNA片段而 获得新的表型的过程。(与转化类似,只是受体细胞不同)
转导(transduction):是指通过病毒(如λ噬菌体)颗粒感染宿主细 胞将外源DNA分子导入到受体细胞内并稳定遗传的过程。
DNA的基本操作技术——聚合酶链式反应技术
DNA的基本操作技术——聚合酶链式反应技术
常用的PCR反应体系:引物、DNA聚合酶、dNTP、模板、缓冲液。
常用的PCR反应程序:
预变性 95℃ 3 min
变性 95℃ 30 s
退火 55℃ 30 s
x 25
延伸 72℃ 1 min
《分子生物学》期末试卷及答案(A)

《分子生物学》期末试卷(A)一、术语解释(20分,每题2分)1、cDNA2、操纵子3、启动子4、内含子5、信号肽6、单顺反子mRNA7、顺式作用元件8、复制叉9、密码子的简并性10、转录因子二、选择题(20分)1.指导合成蛋白质的结构基因大多数为: ( ) A.单考贝顺序 B.回文顺序C.高度重复顺序D.中度重复顺序2. 下列有关Shine-Dalgarno顺序(SD-顺序)的叙述中错误的是: ( )A.在mRNA分子的起始密码子上游7-12个核苷酸处的顺序B.在mRNA分子通过SD序列与核糖体大亚基的16s rRNA结合C.SD序列与16s rRNA 3'端的一段富含嘧啶的序列互补D. SD序列是mRNA分子结合核糖体的序列3.原核生物中起始氨基酰-tRNA是: ( )A.fMet-tRNAB.Met-tRNAC.Arg-tRNAD.leu-tRNA4.下列有关TATA盒(Hognessbox)的叙述,哪个是错误的: ( )A. 保守序列为TATAATB.它能和RNA聚合酶紧密结合C. 它参与形成开放转录起始复合体D.它和提供了RNA聚合酶全酶识别的信号5. 一个mRNA的部分顺序和密码的编号是140 141 142 143 144 145 146CAG CUC UAU CGG UAG AAC UGA以此mRNA为模板,经翻译生成多肽链含有的氨基酸为: ( )A.141B.142C.143D.1446. DNA双螺旋结构模型的描述中哪一条不正确:( )A.腺嘌呤的克分子数等于胸腺嘧啶的克分子数B.同种生物体不同组织中的DNA碱基组成极为相似C.DNA双螺旋中碱基对位于外侧D. 维持双螺旋稳定的主要因素是氢键和碱基堆集力。
7. DNA聚合酶III的描述中哪条不对:( )A.需要四种三磷酸脱氧核苷酸作底物B.具有5′→3′外切酶活性C. 具有5′→3′聚合活性D. 是DNA复制中链延长反应中的主导DNA聚合酶8.与mRNA的GCU密码子对应的tRNA的反密码子是: ( )A.CGAB.IGCC.CIGD.CGI9.组蛋白在生理pH条件下的净电荷是:()A. 正 B . 负 C. 中性 D. 无法确定10.转录需要的原料是( )A. dNTPB. dNDPC. dNMPD. NTP11.DNA模板链为 5’-ATTCAG-3 ’ , 其转录产物是: ( )A. 5 ’ -GACTTA-3 ’B. 5 ’ -CTGAAT-3 ’C. 5 ’ -UAAGUC-3 ’D. 5 ’ -CUGAAU-3 ’12.DNA复制和转录过程有许多相同点,下列描述哪项是错误的? ( )A.转录以DNA一条链为模板,而以DNA两条链为模板进行复制B. 在这两个过程中合成均为5`-3`方向C. 复制的产物通常情况下大于转录的产物D. 两过程均需RNA引物13.下面那一项不属于原核生物mRNA的特征()A:半衰期短 B:存在多顺反子的形式C:5’端有帽子结构 D:3’端没有或只有较短的多聚A.结构14.真核细胞中的mRNA帽子结构是()A. 7-甲基鸟嘌呤核苷三磷酸B. 7-甲基尿嘧啶核苷三磷酸C. 7-甲基腺嘌呤核苷三磷酸D. 7-甲基胞嘧啶核苷三磷酸15.下面哪一项是对三元转录复合物的正确描述()A.σ因子、核心酶和双链DNA在启动子形成的复合物B.全酶、模板DNA和新生RNA形成的复合物C.三个全酶在转录起始点形成的复合物D.σ因子、核心酶和促旋酶形成的复合物16.下列哪组氨基酸只有一个密码子?()A.苏氨酸、甘氨酸B.脯氨酸、精氨酸C.丝氨酸、亮氨酸D.色氨酸、甲硫氨酸E.天冬氨酸和天冬酰胺17.tRNA分子上结合氨基酸的序列是()A.CAA-3′B.CCA-3′C.AAC-3′D.ACA-3′E.AAC-3′18.下列关于遗传密码的知识中错误的是()A.20种氨基酸共有64个密码子B.碱基缺失、插入可致框移突变C.AUG是起始密码D.一个氨基酸可有多达6个密码子19.下列不是蛋白质生物合成中的终止密码是( )。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
各种参与剪接的成分形成一个剪接体系, 称为剪接体(spliceosome)。该体系由 几种snRNP和大量的其他的蛋白质分子 组成,这些蛋白质分子称为剪接因子, 估计有40多种。 剪接点和分支点序列由剪接体识别, snRNA和蛋白质都参与了识别,特别是 snRNA之间以及与mRNA间的碱基配对 起重要作用。
RNA印迹法(RNA blotting)可用于分析核 RNA剪接过程中的中间体,从而确定内含 子的去除顺序。 实验中可以发现含有不同内含子的中间产 物,因此内含子的去除似乎没有特定的顺 序,但也发现有一定的规律,说明剪接有 特定的途径。
剪接反应并不是按内含子在RNA前体上 的顺序进行的。 RNA的构象影响剪接点的选择。在去除 特定的内含子后,RNA的构象发生一定 的变化,再选择新的剪接点。
剪接体Splicesome: snRNA:U1, U2, U4, U5, U6 snRNP:U+蛋白 质
剪接体按一定的顺序组装,已分离到一些 组装的中间体。只有组装完整的剪接体才 有功能。在剪接反应中,剪接体还会释放 和添加某些成分。 转酯反应只是磷酸酯键的直接转移,没有 水解反应的出现,因此不需要外部的能量, 也不需要供能物质如ATP或GTP的参与。 剪接体中起催化转酯反应的成分尚未弄清 楚,也不知道是蛋白质还是RNA在起作用。
一、核基因mRNA的剪接 一、核基因mRNA的剪接
mRNA的基因在低等真核生物中只有少数含 有内含子,为不连续基因。随着进化程度的 增高,不连续基因的数目不断增加。 细胞核中有一类RNA,十分不稳定,平均长 度比mRNA要长,序列的复杂程度非常高, 称为核内不均一RNA( heterogeneous nuclear RNA,hnRNA )。
RNA processing 1
5’ end capping
The “cap”: a methylated guanine joined to the RNA transcript by a 5’-5’ linkage The linkage contains 3 phosphates 3 sequential enzymatic reactions Occurs early
真核生物mRNA的稳定性可能取决于去稳定 元件(destablizing element)。将这一元件 与mRNA结合,可引起该mRNA的降解。 参与RNA降解的核酸酶类可能比参与RNA 成熟过程的核酸酶特异性差。
第六节 RNA的剪接 RNA的剪接
割裂基因(split gene)即不连续基因 (interrupted gene)是指编码某一RNA的基 因中有些序列并不出现在成熟RNA序列中, 成熟RNA的序列在基因中被其他的序列间隔 开,这些序列称为介入序列,又称为内含子 (intron)。被内含子隔开的、出现在成熟 RNA中的序列称为外显子(exon)。
(2) 3’ poly(A)尾巴
多数真核生物(酵母除外)的mRNA 3’末端具 有长约200 bp的poly(A)尾巴。 poly(A)尾巴不是由模板DNA所编码,而是 在转录后由RNA末端腺苷酸转移酶(RNA terminal riboadenylate transferase)催化下,以 ATP为前体,添加到mRNA的3’末端形成的。
第二个阶段,内含子的3’剪接点被切断 而内含子以套索状释放,与此同时右侧 外显子与左侧外显子连接在一起。 第三个阶段,内含子的套索被切断,形 成线状并很快被降解。
参与真核细胞核mRNA前体拼接反应的顺式元件 前体拼接反应的顺式元件 参与真核细胞核
完成剪接所需要的条件只有三个序列,即 5’GT、3’AG和分支点序列。内含子和外显子 其他序列的缺失并不影响剪接。也说明剪接 过程不需要内含子或外显子的特定构象。 分支点周围的序列也有一定程度的保守性。 突变实验证明,分支点的作用是选择距之最 近的3’剪接点作为与5’剪接点相连的靶位点。
核基因mRNA的剪接位点(splicing site) 是指内含子与外显子的断裂点以及相邻 外显子的连接点。 经过比较mRNA与其基因的序列发现, 在内含子两端分别有两个非常保守的碱 基,左剪接位点为GU,右剪接位点为 AG。内含子在剪接位点的这种特征称为 GU-AG规则(GU-AG role) 。
有些mRNA的3’末端没有poly(A)尾巴, 如组蛋白的mRNA,其3’末端的正确形成 依赖于RNA本身形成的茎环结构,即转录 终止于此处。 对茎环结构进行突变,只要是维持其二级 结构,就不影响转录的终止。说明二级结 构所提供的信息比序列本身更重要。
加尾反应需要的顺式元件
1. CPSF (cleavage and polyadenylation specificity factor) & CstF (cleavage stimulation factor) bind to the poly-A signal, leading to the RNA cleavage 2. Poly-A polymerase (PAP) adds ~ 200 As at the 3’ end of the RNA, using ATP as a substrate
左(5’)剪接点称为供体位点,右(3’)剪 接点称为受体位点。点突变研究证明,剪 接位点GT或AG的突变均可以阻止剪接的出 现。 几乎所有的真核细胞核基因的内含子均遵 循GU-AG规则,这也暗示这类内含子存在 着共同的剪接机制。但线粒体、叶绿体和 酵母tRNA的内含子不遵循这一规则。
核RNA的剪接点及周围并没有自身的互补序列, 5’剪接点总是能正确地与3’剪接点相连。 剪接位点具有通用性,而内含子的其他序列 (包括二级结构)不提供特定的信息,对剪接 不产生影响。剪接装置没有组织特异性,RNA 只要含有正确的剪接点,在任何细胞中都可以 正确地剪接。
套索是通过内含子5’的保守G与分支点A 之间形成磷酸二酯键而形成的,这两个 碱基在所有的真核生物中高度保守。 在剪接过程中,没有酶的参与,但存在 一种机制在反应前对上述保守的序列元 件进行检查。只有这些元件都存在的情 况下剪接反应才会开始,而任一元件的 严重缺失都会抑制剪接反应。
SR proteins, bound to exonic splicing enhancers (ESEs), interact with components of splicing machinery, recruiting them to the nearby splice s核细胞核 的加帽反应
不同真核生物的mRNA可有不同的帽子结构, 同一种真核生物的mRNA也常有不同的帽子 结构。 帽子结构的作用: 1.为核糖体识别RNA提供信号 2.增加mRNA的稳定性 3.为mRNA向胞质的运输提供信号 4 与某些RNA病毒的正链RNA的合成有关。
2. 核基因mRNA剪接的机制 核基因mRNA剪接的机制
核提取物能完成mRNA前体的剪接,说 明mRNA的剪接与转录过程无关。 mRNA的剪接与RNA的修饰状态也没有 关系,没有帽子结构或poly(A)尾巴的 RNA都能完成剪接。
剪接可以分为三个阶段进行: 第一阶段,内含子的5’端切开,形成游离的 左侧外显子和右侧的内含子和外显子分子。 左侧的外显子呈线状,而右侧的内含子和 外显子并不呈线状。与在距内含子的3’端约 30个碱基处有一个高度保守的A,称为分支 位点(branching site)。内含子游离的5’端 以5’-2’磷酸二酯键与A相连,形成一个套索 (lariat)结构。
2. mRNA的转录后加工 mRNA的转录后加工
在真核生物中,几乎所有的成熟mRNA有 5’帽子(cap)结构,多数还有3’末端的 poly(A)尾巴。这些结构都是在转录后 经过修饰的结果。
(1)5’帽子结构 成熟真核生物的mRNA并没有游离的5’端, 而是一种被称为帽子的结构。 真核生物的帽子结构可归纳为三种: m7GpppX为帽子0 m7GpppXm为帽子1 m7GpppXmYm为帽子2
3. 后加工与mRNA的稳定性 后加工与mRNA的稳定性
转录后加工是产生各种成熟RNA分子的重要 过程。 各种RNA分子的加工都是在特定的酶参与下 完成的,包括核酸内切酶和核酸外切酶。核 酸酶除了参与后加工过程外,还负责过剩 RNA的降解。
原核生物mRNA降解的方向为5’至3’ ,可能 由内切酶在最后一个核糖体后面发挥作用。 余下的片段由外切酶按3’至5’的方向水解成 核苷酸。 mRNA分子结构中的二级结构可以阻止外切 酶的作用。
Polyadenylation and termination
真核细胞mRNA前体加尾反应模式图 真核细胞 前体加尾反应模式图
RNA polII CTD的磷酸化与加尾和加帽反应的关系 的磷酸化与加尾和加帽反应的关系
poly( A )在分子生物学实验中有很大的 应用价值。 1. 应用mRNA的该特性,可用寡聚T(oligo dT)为引物,反转录合成cDNA。 2. 将oligo( dT )与介质相连,用于从总 RNA中分离纯化mRNA。
二、真核生物的转录后加工
真核生物转录后加工除了mRNA外, tRNA和rRNA也要经过后加工的过程。 有关真核生物tRNA的加工处理了解还不 多,可能和原核生物有相似之处。
1. rRNA的转录后加工 rRNA的转录后加工
真核生物有4种rRNA,即5.8S rRNA、18S rRNA、28S rRNA和5S rRNA。其中前三者 的基因组成一个转录单位,产生47S的前 体,并很快转变成45S前体。 45S前体上有许多甲基化的位点,在转录 过程中或转录后被甲基化。甲基基团主要 是加在核糖上。甲基化是45S前体最终成 为成熟rRNA区域的标志。
hnRNA以与蛋白质结合的状态存在,形成 hnRNP(核糖核蛋白颗粒)。体外研究表明, hnRNP的形状为一个球体和一个与之相连的 纤维状结构。 在RNA转录生成以后,在核内完成加帽和 加尾的同时,内含子的去除即剪切过程也 在进行。然后RNA转运到胞浆。