第八章氨基酸发酵机制.
氨基酸发酵生产工艺学n2ppt课件

(二)pH的影响及其控制
作用机理:主要影响酶的活性和菌的代谢。 在氮源供应充分和微酸性条件下,谷氨酸发 酵转向谷氨酰胺发酵。 pH控制在中性或微碱性。 方法:流加尿素和氨水。
我国味精技术进展情况
制糖工艺进展:酸法水解→酶酸法水解→双酶法水解。 发酵工艺进展:亚适量生物素水平(产酸4~6g/dl)
→高生物素水平(产酸12~15g/dl)。 提取工艺进展:等电点法(少数锌盐法)→等电离交法
→低温连续等电点法(少数厂家采用)。 精制工艺进展:全粉炭脱色、硫化碱除铁→颗粒炭脱
+ 4
生物 谷氨酸 (限量) 乳酸或琥珀酸(充足) 素
pH (酸性)N-乙酰-谷氨酰胺 谷氨酸(中性或微碱性)
磷酸 (适量)谷氨酸 盐
缬氨酸
➢ 菌种扩大培养
1、斜面培养:主要产生菌是棒状杆菌属、 短杆菌属、小杆菌属、节杆菌属。
我国各工厂目前使用的菌株主要是钝齿 棒杆菌和北京棒杆菌及各种诱变株。
生长特点:适用于糖质原料,需氧, 以生物素为生长因子。
2. 不溶性盐ห้องสมุดไป่ตู้淀法
(1)锌盐法
谷氨酸+锌离子 pH6谷.3 氨酸锌沉淀 pH2.谷4 氨酸结晶
溶加液酸
(2)盐酸盐法: Glu在浓盐酸中生成并析出谷氨酸盐酸盐。
这是用盐酸水解面筋生产谷氨酸的原理。 (3)钙盐法:
高温谷氨酸钙溶解度大,与菌体等不溶性杂质 分开,降温,析出谷氨酸钙沉淀,加NaHCO3 直接得 到味精。
3、菌体生长停滞期:谷氨酸合成。
措施:提供必须的氨及pH维持在7.2-7.4。 大量通气,控制温度34-37 ℃。
氨基酸发酵

|
|
和 嘧 啶 的 前 体 物 , 尤 其 是 AT P , 它 还 直 接
参与组氨酸的合成,所以利用嘌呤的结构 类似物抗性突变株解除反馈调节可以达到 增加组氨酸前体物的积累。
途径也能增加HMP途径的流量。
5. HMP途径的阻断 由于合成组氨酸需要HMP途径的中间代谢产物, 所以完整的HMP途径会减少D-核糖的生成,通过 阻断转酮酶来破坏完整的HMP途径,从而使代谢
谷氨酸脱氢酶和谷氨酰胺合成酶是保证α -酮戊二酸向 谷氨酰胺而不是向草酰乙酸的三羧酸循环方向代谢的 关键酶。同时在该循环中还存在着向天冬氨酸、丙氨 酸、缬氨酸的分支代谢,设法减弱分支代谢而强化主 流代谢,主要的方法是减弱催化这些分支代谢酶的酶 活。
3
谷氨酰胺发酵
|
谷氨酰胺合成酶的最佳pH为 6.5 ~ 7.0 , 而 谷 氨 酰 胺 酶 、 N- 乙 酰 谷 氨 酰 胺 脱 乙 酰 酶 的
1
丙氨酸发酵 丙 氨 酸白
1
丙氨酸是构成蛋白质的基本单位,是组成人体蛋 质 的 20 种 氨 基 酸 之 一 。 它 的 分 子 式 是
|
|
(alanine) C3H7NO2 , 有 α - 丙 氨 酸 和 β- 丙 氨 酸 两 种 同 分
异构体。
2
功能
预防肾结石、协 助葡萄糖的代谢, 有助缓和低血糖, 改善身体能量。
3
用途
用于合成新型甜 味剂及某些手性 药物中间体的原 料。 点石演示
简介
点石演示
点石演示
1
丙氨酸发酵
|
73% 93% 80% 78%
|
丙氨酸性状
丙氨酸是构成蛋白质的基本单位, 是组成人体蛋白质的20种氨基酸
氨基酸在发酵中的作用

氨基酸在发酵中的作用一、引言发酵是一种利用微生物代谢产物进行生物转化的过程,广泛应用于食品工业、饲料工业和生物制药等领域。
而氨基酸作为生物体内重要的有机物,也在发酵过程中发挥着重要的作用。
本文将从氨基酸在发酵中的作用机制、应用及前景等方面进行探讨。
二、氨基酸在发酵中的作用机制1.提供碳源和能源:氨基酸是微生物合成蛋白质的基本单元,可以被微生物利用作为碳源和能源。
在发酵过程中,微生物通过代谢氨基酸产生能量,并将其转化为所需的代谢产物。
2.调节酶活性:氨基酸可以作为酶的辅因子,调节酶的活性。
在发酵过程中,一些关键酶的活性会受到氨基酸的调节,从而影响代谢途径的进行。
3.调节细胞内pH值:氨基酸在细胞内可离子化,释放出氢离子或吸收氢离子,从而调节细胞内的pH值。
适宜的pH值对微生物的生长和代谢具有重要的影响,氨基酸可以在发酵过程中维持适宜的pH值,提高发酵效率。
4.提供氮源:氨基酸中的氨基团含有丰富的氮元素,可以作为微生物合成蛋白质和其他氮化合物的氮源。
发酵过程中,微生物利用氨基酸中的氨基团合成所需的氮化合物,促进代谢产物的合成。
三、氨基酸在发酵中的应用1.食品工业:氨基酸可以作为发酵食品中的营养添加剂,提供微生物生长所需的营养物质,促进发酵过程。
例如,在酱油、酱料和味精等食品的发酵中,氨基酸作为调味品添加剂,不仅能够提高食品的口感和风味,还能够增强食品的营养价值。
2.饲料工业:氨基酸作为饲料添加剂,可以提高动物的生长性能和免疫力。
在畜禽饲料中添加适量的氨基酸,有助于提高饲料的利用率,降低环境污染,达到绿色养殖的目的。
3.生物制药:氨基酸在生物制药中的应用十分广泛。
一方面,氨基酸可以作为生物药物的原料,通过发酵合成所需的蛋白质药物;另一方面,氨基酸也可以作为生物药物的稳定剂,保护药物的活性和稳定性,提高药物的疗效。
四、氨基酸在发酵中的前景随着生物技术的不断发展,发酵工艺在各个领域中的应用越来越广泛。
而氨基酸作为重要的发酵辅助剂,其应用前景也日益广阔。
氨基酸发酵机制及过程概述

葡萄糖和琥珀酸等对异柠檬酸裂解酶起着阻遏作用。
(1)以糖质为原料的谷氨酸发酵中生物素对 DCA循环的影响
在生物素亚适量条件下,琥珀酸氧化力降低, 积累的琥珀酸会反馈抑制异柠檬酸裂解酶活性,并 阻遏该酶的生成,DCA循环基本处于封闭状态, 异柠檬酸高效率地转化为α–酮戊二酸,再生成谷 氨酸。 在生物素充足的条件下,异柠檬酸裂解酶活性 增大,通过DCA循环提供能量,进行蛋白质的合 成,不仅异柠檬酸转化生成谷氨酸的反应减弱使得 谷氨酸减少,而且生成的谷氨酸在转氨酶的催化作 用下又转成其它氨基酸,也不利于谷氨酸积累。
7.醋酸或正石蜡原料发酵谷氨酸的推测途径 在醋酸发酵谷氨酸或石油发酵谷氨酸时,却只能 经乙醛酸循环供给四碳二羧酸,四碳二羧酸经草酰 乙酸又转化为柠檬酸。
二、谷氨酸生物合成的代谢调节机制
分解代谢: 从环境中摄取营养物质,把它们转 微生物 的代谢
化为自身物质,以此来提供能源和 小分子中间体;
合成代谢:合成代谢将分解代谢产生的能量和
6乙酰CoA +2NH3+3O 2谷氨酸 +2CO2+6H2O 2
ቤተ መጻሕፍቲ ባይዱ
3mol葡萄糖可以生成2mol的谷氨酸,谷氨 酸对葡萄糖的质量理论转化率为:
(3)实际转化率:处于二者之间,即54.4%~ 81.7%。 CO2固定反应、乙醛酸循环的比率等对转化率影 响较大。 乙醛酸循环活性越高,谷氨酸越不易生成与积累。
α -酮戊二酸脱氢酶 NH4 异柠檬酸脱氢酶 乙醛酸循环中的两个关键 酶——异柠檬酸裂解酶和 苹果酸合成酶。
谷氨酸 (胞内)
转移到胞外
(二)谷氨酸合成的理想途径
生物素充足菌EMP所占比例约为62%; 在发酵产酸期,EMP所占比例更大,约为74%。
天冬氨酸族氨基酸发酵机制

优化发酵条件提高产量
通过实验手段优化天冬氨酸族氨基酸发酵的条件,如温度、pH、溶氧浓 度、碳氮源等,以提高发酵效率和产物产量。
开发新型的细胞培养技术,如连续培养、高密度培养等,以实现细胞的高 密度生长和产物的高效积累。
结合代谢工程和基因工程技术,对菌株进行遗传改造,提高其发酵性能和 产物产量。
开发新型天冬氨酸族氨基酸生产菌株
营养强化剂
天冬氨酸族氨基酸可添加到婴儿食品、运动员食品等特殊食品中,以满足特定营养需求。
在农业领域的应用前景
植物生长调节剂
天冬氨酸族氨基酸可促进植物生长,提 高产量,可作为潜在的植物生长调节剂 。
VS
饲料添加剂
天冬氨酸族氨基酸可添加到动物饲料中, 提高动物生长性能和饲料转化率。
06
研究展望
深入探究天冬氨酸族氨基酸发酵机制
天冬氨酸族氨基酸发酵机 制
• 引言 • 天冬氨酸族氨基酸发酵机制概述 • 天冬氨酸族氨基酸发酵的生物合成途
径 • 天冬• 研究展望
01
引言
氨基酸发酵的重要性
生命活动的基础
氨基酸是构成蛋白质的基本单位,是 生物体进行生命活动不可或缺的物质。
04
天冬氨酸族氨基酸发酵的代谢调控
代谢调控机制
酶的活性调节
通过调节酶的活性来控制代谢过 程,例如通过调节天冬氨酸族的 合成酶活性来控制氨基酸的合成。
代谢流控制
通过控制代谢流的方向和流量来调 节氨基酸的合成,例如通过控制糖 酵解和三羧酸循环的代谢流来影响 天冬氨酸族的合成。
代谢物阻遏
通过调节细胞内代谢物的浓度来控 制基因的表达和酶的活性,从而影 响氨基酸的合成。
通过诱变育种、基因重组等手段,发掘和培育具有优良发酵性能的新型天冬氨酸族 氨基酸生产菌株。
《氨基酸发酵工艺》PPT课件

3 氨基酸发酵的工艺控制
培养基
2、氮源:铵盐、尿素、氨水,同时调整pH值。 营养缺陷型需要添加适量氨基酸时,主要添加有 机氮源水解液。 需生物素和氨基酸时,以玉米浆作氮源。 尿素灭菌时分解或形成磷酸铵镁盐,须单独 灭菌,40%的尿素可在108℃40min,高温会生产 缩脲。 氨水用pH自动控制连续流加。
3 氨基酸发酵的工艺控制
培养基
3、碳氮比
氮源除用于菌体生长外,还用于氨基酸合成, 在氨基酸发酵中常常是用氨水、尿素来调节pH, 所以氨基酸发酵所用的C/N比一般微生物发酵的 低,或者说氮源用量更高。
谷氨酸发酵在C/N小于100:11时才开始 积累谷氨酸,所以通常情况下谷氨酸发酵 C/N为100:(15~30)。
4 谷氨酸生产工艺
日本和德国为世界主要氨基酸生产国。日本的 味之素、协和发酵及德国的德固沙是世界氨基酸生 产的三巨头。它们能生产高品质的氨基酸,可直接 用于输液制剂的生产。日本在美国、法国等建立了 合资的氨基酸生产厂家,生产氨基酸和天冬甜精等 衍生物。
1 概述
氨基酸的国内生产概况
天津氨基酸公司、湖北八峰氨基酸公司 生产 规模及产品质量与国外大厂有较大差距。
2 氨基酸发酵的代谢控制
消除终产物的反馈抑制与阻遏作用
消除终产物的反馈抑制与阻遏作用,是通过使用 抗氨基酸结构类似物突变株的方法来进行。
例:利用抗性突变株消除S-(β-氨基乙酸)-L-半胱氨酸 (即AEC)(赖氨酸的结构类似物)与L-苏氨酸的协 同抑制。
2 氨基酸发酵的代谢控制
2 氨基酸发酵的代谢控制
4 谷氨酸生产工艺
谷氨酸生产概述
➢ 起初工业化生产采取小麦或大豆蛋白质水解制取。
➢ 1957年,日本率先采用微生物发酵生产,并实现工 业化,被誉为现代发酵工业的重大创举,使发酵工业 进入调节调控水平。
氨基酸发酵机制及过程

④沿着由柠檬酸至α–酮戊二酸的氧化途径,谷氨酸产 生菌有两种NADP专性脱氢酶,即异柠檬酸脱氢酶和L谷氨酸脱氢酶。 在谷氨酸的生物合成中,谷氨酸脱氢酶和异柠檬 酸脱氢酶在铵离子存在下,两者非常密切地偶联起 来,形成强固的氧化还原共轭体系,不与NADPH2 的末端氧化系相连接,使α–酮戊二酸还原氨基化生 成谷氨酸。
2.三羧酸循环(TCA循环)的调节
谷氨酸产生菌在代谢途径中,三羧酸循环的调节主要 是通过5种酶的调节进行的。这五种酶是磷酸烯醇式丙酮 酸羧化酶、柠檬酸合成酶、异柠檬酸脱氢酶、谷氨酸脱氢 酶和α-酮戊二酸脱氢酶。
草酰乙酸+谷氨酸
谷氨酸转氨酶
天冬氨酸+α -酮戊二酸
谷氨酸比天冬氨酸优先合成,谷氨酸合成过量后, 谷氨酸抑制谷氨酸脱氢酶的活力并阻遏柠檬酸合成酶 的合成,使代谢转向天冬氨酸的合成。
在谷氨酸发酵生产中,生物素缺陷型菌在 NH4+存在时,葡萄糖消耗速率快而且谷氨酸收率 高; NH4+不存在时,葡萄糖消耗速率很慢,生 成物是α–酮戊二酸、丙酮酸等物质,不产生谷氨 酸。
四、细胞膜通透性的调节
对谷氨酸发酵的重要性: 当细胞膜转变为有利于谷 氨酸向膜外渗透的方式, 谷氨酸才能不断地排出细 胞外,这样既有利于细胞 内谷氨酸合成反应的优先 性、连续性,也有利于谷 氨酸在胞外的积累。
4.CO2固定反应的调节
CO2固定反应主要通过以下途径完成:
C02的固定反应的作用:补充草酰乙酸; 在谷氨酸合成过程中,糖的分解代谢途径与C02固定 的适当比例是提高谷氨酸对糖收率的关键问题。
5.NH4+的调节
谷氨酸脱氢酶也能催化谷氨酸氧化脱氨反应,脱 氨过程以NAD+作为辅酶,该酶催化的反应虽然偏向 氨合成谷氨酸一边,但是脱氢过程产生的NADH被氧 化成NAD+,同时产生的NH3很容易被除去。 脱氨反应被NH4+和α–酮戊二酸所抑制,这对于谷 氨酸的积累也起到了很好的作用。
第8章发酵机制 51-100
实验证明,谷氨酸产生菌的谷氨酸生物合成途 径与一般微生物是一样的,没有特殊的途径, 就是糖经过酵解途径(EMP)和单磷酸己糖途 径(HMP)生成丙酮酸。一方面,丙酮酸氧化 脱羧生成乙酰-CoA;另一方面,经CO2固定作 用生成草酰乙酸;两者合成柠檬酸进入三羧酸 循环(TCA循环),由三羧酸循环的中间产物 α–酮戊二酸,在谷氨酸脱氢酶催化下,还原氨 基化合成谷氨酸。
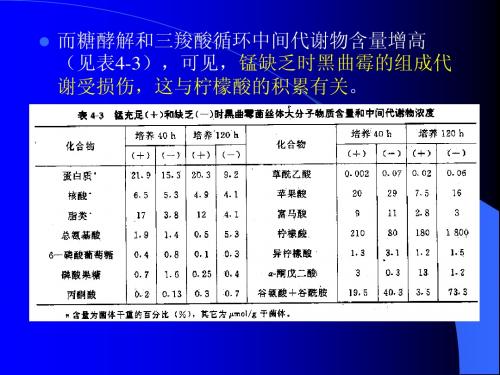
Mn2+ 对棒杆菌核苷酸积累的专一性影响 和上述例子极为相似。关于核苷酸的积 累,人们推断是因为催化核糖核苷酸形 成聚合阶段第一步反应的酶需要锰。
某些真菌的丙酮酸是酵解过程第二个调 节点已被证明,但是关于黑曲霉尚未被 证明。用交换定理未能确定这一步反应 是否调节性。测定柠檬酸发酵时酵解中 间代谢物浓度可推断流经丙酮酸激酶的 通量增加。
当乙酰 -CoA 氧化,使 ATP 水平提高,结 果丙酮酸激酶受 ATP抑制, PEP又向 CO2 固定。当生物合成和需能反应使 ATP 浓 度降低,则 ATP 对丙酮酸激酶的抑制被 解除,此酶被 AMP激活,使乙酰-CoA生 成量增加。
经过深入研究,他们通过实验证明在黄 色短杆菌中存在α-酮戊二酸脱氢酶络合 物,并发现该酶络合物极不稳定。该酶 的活力很弱,使糖代谢流进入TCA循环 后受阻在α-酮戊二酸处,在有NH存在下, 由谷氨酸脱氢酶催化还原氨基化生成谷 氨酸。
这一点也为木下等对TCA循环中有机酸的氧化 能的研究结果所证实,如表5-2所示。柠檬酸、 顺乌头酸、异柠檬酸和α-酮戊二酸的氧化能力 几乎没有,即α-酮戊二酸脱氢酶活性很弱。
二、TCA、DCA和CO2固定作用 1. TCA循环 糖经过 EMP、HMP 途径生成丙酮酸,在 有氧条件下,氧化脱羧生成乙酰 -CoA, 另一方面经CO2固定生成草酰乙酸,两者 在柠檬酸合成酶催化下生成柠檬酸,进 入TCA循环。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
• • • • • • • •
4.根据代谢作用将酶分类 (1)调节酶 变构酶 同工酶 多功能酶 (2)静态酶 (3)潜在酶 关键酶:参与代谢调节的酶的总称。
5.氨基酸合成的关键酶
(1)磷酸果糖激酶 (2)柠檬酸合酶 (3)N-乙酰谷氨酸激酶 (4)鸟氨酸转氨基甲酰酶 (5)天冬氨酸激酶 (6)高丝氨酸脱氢酶 (7)苏氨酸脱水酶 (8)a-乙酰羟酸合成酶 (9)DNAP合成酶 (10)分支酸变位酶 (11)预苯酸脱水酶 (12)预苯酸脱氢酶
① ② ⑨
⑩ ⑿ ⑾
⑤ ③ ⑥
⑦ ④
⑧
第二节、酶活性的控制
• 别构效应、共价修饰、寡聚酶的解聚与聚合、 蛋白酶水解激活 • 一、变构调节 • (一)变构效应(别构效应) • 1.别构效应:某种不直接涉及蛋白质活性的物质,结
合于蛋白质活性部位以外的其他部位(别构部位), 引起蛋白质分子的构象变化,而导致蛋白质活性改变 的现象。
目前已知可以受化学修饰调节的酶几乎又都是别构酶。
共价调节酶最典型的例子是动物组织的糖原磷酸化酶 1. 糖原磷酸化酶催化的反应
催化糖原的磷酸解,生成G-1-P。 + H3PO4
糖原磷酸化酶
糖原
G-1-P + 糖原
(n-1个特征
3、调节机理 磷酸化酶的磷酸化与脱磷酸化 磷酸化酶构象的变化(包括亚基的聚合与 解离 化学信号的放大(级联放大)
ATCase活性调节的机理
C C C
汞盐
R R R R R R
C C
C C
C
+
C
催化亚基 (三聚体)
R R R R
R R
C
C
C
完整的 ATCase (活性)
调节亚基 (二聚体)
CTP ATP
有催化活性的构象
无催化活性的构象
5.负协同效应(negative cooperative effect)
• • • • • •
2.氨基酸发酵的种类 (1)使用野生型菌株发酵 (2)使用营养缺陷型菌株发酵 (3)使用抗类似物突变株发酵 (4)添加前体法 (5)酶法转化法
• • • • • • • • •
3.微生物调节控制因素: (1)酶的浓度 诱导 阻遏 (2)酶的活性 终产物的抑制或激活 辅酶水平的活性调节 酶原的活化 潜在酶的活化
大肠杆菌天冬氨酸转氨甲酰酶(E-Coli aspartate transcarbamylase, ATCase)
氨甲酰磷酸 天冬氨酸 氨甲酰天冬氨酸
• (1)Asp浓度增加,ATC酶与底物亲和 力及协同性增加 • (2)CTP反馈抑制ATC酶活性,但不能 全部抑制 • (3)底物浓度饱和而达到最大反应速度 不受抑制剂的影响
酶的磷酸化和脱磷酸化主要在高等动物细 胞中进行,而腺苷酰化和脱腺苷酰化在细菌中 比较常见。
如E.Coli 谷氨酰胺合成酶 腺苷酰化无活性 脱腺苷酰化有活性
第八章
氨基酸发酵机制
第一节、概述 1.代谢控制发酵 遗传学的方法或其他生物化学的方法, 人为的改变、控制微生物的代谢,使有 用产物大量生成、积累的发酵。 条件: (1)搞清楚了微生物细胞各种氨基酸代谢的 遗传控制,代谢调节机制。 (2)可人为的在DNA水平上改变微生物的代 谢 (3)合理的控制环境条件
脱敏作用: 经特定处理后,不丧失酶活性而失去对变 构效应物的敏感性,称脱敏作用。 (1)酶解聚 (2)基因突变
4.正协同效应(positive cooperative effect) 调节物与变构酶结合后,酶的构象发生变化,新的 构象有利于后续底物分子或调节物的结合。
正协同效应别构酶与米氏酶动力学比较
调节物与变构酶结合后,酶的构象发生变化,新的构象不 利于后续底物分子或调节物的结合。
• 3-磷酸甘油酸脱氢酶 • 3-磷酸甘油酸脱氢酶对NAD浓度的较大变化 不受大的干扰。
(二)序变模型和齐变模型
对称或协同模型(symmetry or concerted model,也称齐变模型、MWC模型) 1965年由Monod、Wyman和Changeux提出。
模型的几个假定:
该模型的要点 :主张别构酶所有的亚基或者全部是坚固
紧密的,不利于结合底物的“T”状态,或者全部是松散 的,利于结合底物的“R”状态。这两种状态间的转变对 每个亚基都是同步的,齐步发生的,“T”状态中的亚基 的排列是对称的,变为R状态后,蛋白亚基的排列仍然 是对称的。
序变模型(sequential model,也称KNF模型) 1966年由Koshland、Nemethy和Filmer提出。
• 别构酶:受别构效应调节的酶。 • 效应物:凡使酶发生别构效应的物质,
• 通常为小分子代谢物或辅助因子。
2.别构酶结构上的特点:多个亚基、有四 级结构、活性中心与别构中心可处于不 同的亚基或同一亚基不同的部位。
3.别构酶性质上的特点:当专一性底物与 别构中心诱导楔合时,随着别构中心构 象的改变,该酶的活性中心构象也会相 应发生变化,从而引起酶催化活性的改 变。
模型的几个假定:
该模型的要点 :一个配体可以诱导它要结合的亚基的三级 结构的变化。这个亚基-配体复合体又能够改变相邻亚基的 构象。这一模型特别强调对于配体只有一种形状是亲和力 最高的,但与齐变模型不同的是在具有部分饱和的一个寡 聚分子中允许存在着高亲和力和低亲和力的亚基。
• 调节酶总结:
(1)调节酶一般为变构酶; (2)调节酶反应速度与底物浓度关系多呈S型,有协 同性; (3)调节酶多处于合成代谢途径上而且一般在代谢分 支点,多受终产物反馈抑制; (4)中间产物不影响调节酶的活性; (5)调节酶后面的酶对终产物不敏感; (6)有脱敏作用; (7)调节酶与负效应物非共价结合引起的变构调节, 是酶活性控制的一种快速、敏感、高效的细调方式。
(二)共价调节酶(covalently modulated enzymes)
什么是共价调节酶?是一类由其它酶对其结构进行可逆共价
修饰,使其处于活性和非活性的互变状态,从而调节酶活性。
常见的是磷酸化/脱磷酸化,腺苷酰化/脱腺苷酰化, 乙酰化/脱乙酰化,尿苷酰化/脱尿苷酰化,甲基化/脱甲 基化,S-S/SH相互转变,共6种类型。