细胞生物学第十章细胞核与染色体
细胞生物学学习资料(第9-11章)

第九章第一节细胞骨架是指真核细胞内由一些特异蛋白质构成的纤维网架结构。
广义的细胞骨架包括细胞外基质、膜骨架、细胞质骨架和核骨架。
细胞质骨架包括微丝、微管和中间丝3种结构。
一、微丝的组成及其组装(一)结构与组成微丝的主要结构成分是肌动蛋白。
肌动蛋白细胞内有两种存在形式,即肌动蛋白单体(又称球状肌动蛋白,G-ac-tin)和由单体组装而成的纤维状肌动蛋白。
肌动蛋白单体外观呈蝶状,中央有一个裂口,可结合ATP和Mg2+。
每条微丝由二股螺旋状相互盘绕而成,每条丝由肌动蛋白单体头尾相连呈螺旋状排列而成。
(二)微丝的组装及动力学特性当溶液中含有适当浓度的Ca2+,而Na+和K+的浓度很低时,微丝趋向于解聚。
当溶液中含有A TP、Mg+以及较高浓度的Na+和k+时,G-actin趋向于组装成微丝。
组装时,正极组装速度比负极快。
解聚时,正极解聚速度比负极更快。
A TP-G-actin处于临界浓度时,微丝组装和去组装过达到平衡,此时称踏车行为。
体外组装过程中具有踏车行为。
组装过程:成核(Arp2和Arp3等蛋白形成起始复合物)--纤维的延长(ATP帽与ADP帽)--进入稳定期(三)影响微丝组装的特异性药物细胞松弛素可将微丝切断,但对微丝解聚没有明显影响。
鬼笔环肽对微丝的解聚有抑制作用,可使肌动蛋白丝保持稳定状态。
二、微丝网络动态结构的调节与细胞的运动(一)非肌肉细胞内微丝的结合蛋白微丝结合蛋白的类型:成核蛋白、单体-隔离蛋白、单体-聚合蛋白、成束蛋白、封端蛋白、纤维-解聚蛋白、网络-形成蛋白、纤维切割蛋白、膜结合蛋白等。
维丝结合蛋白对微丝组装具有调节作用,如隔离、聚合和成束等。
(二)细胞皮层在紧贴细胞质膜的细胞质区域由微丝结合蛋白将微丝交联形成了凝胶状三维网格结构,这个区域通常称为细胞皮层。
细胞的多种运动方式(如胞质环流、阿米巴运动、变皱膜运动、细胞吞噬)都与细胞凝胶态或胶态的转化有关。
(三)应力纤维在紧贴黏合斑的质膜内侧由微丝紧密排列成束,这种微丝束称为应力纤维。
细胞生物学翟中和编 第十章 细胞核与染色体
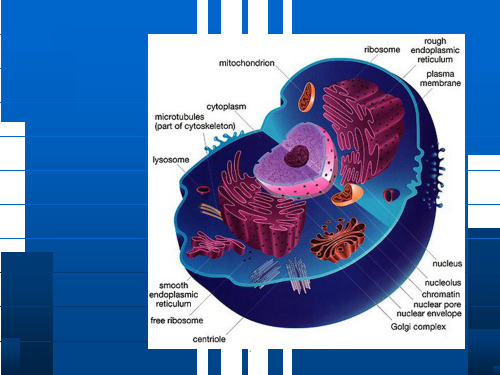
.
染 色 质---成分
➢ DNA ➢ 组蛋白 ➢ 非组蛋白 ➢ 少量RNA
DNA 与组蛋白是染色质的稳定成分 非组蛋白与RNA的含量随细胞生理状态的不同而变化
.
染 色 质---成分--DNA
✓ 凡是具有细胞形态的生物其遗传物质都是DNA ✓ 只有少数病毒的遗传物质是RNA
.
的通道。
核被膜
.
核被膜结构
外核膜
核周隙 内核膜
.
核被膜-外核膜
外核膜:上有核糖体 ,与糙面内质网相连。
✓外核膜被认为是粗面内质网的特化区域,有利于核被 膜与内质网间的物质交流及核被膜的更新。
✓外核膜的外表面存在网状分布的中间纤维,与细胞核 在细胞质的定位有关。
.
核被膜-内核膜
外核膜 核膜间隙:20-40nm,与糙面内质网腔相通,腔内电子密
胞质
核
纤
层
结
受体
构
模
B
式A
C
图
外核膜
内核膜 核纤层 蛋白
染色质纤维.
核纤层功能:
(1)维持核孔位置和核被膜的形态; (2)为间期染色质提供附着位点; (3)有丝分裂中,与核被膜的解体和重建有关。
.
.
染色质
染色质:光学显微镜下可见间期核中有一种嗜碱性
很强的的物质,是由DNA,组蛋白,非组蛋白及少量 RNA组成的线性复合结构,是遗传物质的存在形式。
.
细胞核
➢ 真核细胞内最大、最重要的细胞器
核质比=细胞核(体积)/细胞质(体积) 多数细胞的核质比约为10%
细胞核改变是病理状况下细胞坏死的主要标志,与正常细 胞相比,肿瘤细胞核质比增高,大小形态参差不齐,呈现异型 性,表现为核外形不规则。
翟中和细胞生物学第十章总结2(名词解释)

第十章细胞核与染色体1.细胞核:真核细胞中由双层膜所包被的,包含由DNA、组蛋白等组织而成的染色质的细胞器,是细胞内储存遗传物质的场所,也是基因组复制、RNA合成和加工、核糖体组装的场所。
它是细胞内最大的细胞器,真核生物的细胞都有细胞核,只有成熟的红细胞和植物成熟的筛管没有细胞核。
核膜上有核孔及其环状结构形成核孔复合体,它与大分子物质的运输有关。
2.核被膜:真核细胞内细胞质与细胞核之间由双层膜构成,分别称为外核膜与内核膜。
双层核膜上镶嵌有核孔复合体,能选择性地运输核内外物质。
内膜面向核质,内、外膜间有20~40nm的透明空隙,称为核周间隙,膜上有核孔。
3.核被膜的功能:一方面,核被膜构成了核、质之间的天然选择性屏障,将细胞分成核与质两大结构与功能区域,使得DNA复制、RNA转录与加工在核内进行,而蛋白质翻译则局限在细胞质中。
这样既避免了核质间彼此相互干扰,使细胞的生命活动秩序更加井然,同时还能保护核内的DNA分子免受损伤。
另一方面,核被膜调控细胞核内外的物质交换和信息交流。
核被膜并不是完全封闭的,核质之间进行着频繁的物质交换与信息交流。
这些物质交换与信息交流主要是通过核被膜上的核孔复合体进行的。
4.内、外核膜各有特点:①外核膜表面常附有核糖体颗粒,且常常与糙面内质网相连续,使核周间隙与内质网腔彼此相通、从这种结构上的联系出发,外核膜可以被看作是糙面内质网的一个特化区域。
②内核膜表面光滑,无核糖体颗粒附着,但紧贴其内表面有一层致密的纤维网络结构,即核纤层。
内核膜上有一些特有的蛋白成分,如核纤层蛋白B受体(lamin B receptor,LBR)。
5.核纤层:位于核膜内侧,由核纤层蛋白组成的纤维状网络结构。
在与核质相邻的核膜内表面有一层厚30~160nm的网络状蛋白质,叫核纤层,对核被膜起支撑作用。
核纤层由3种分子量为6~7万道尔顿的多肽亚单位α、β、γ所组成,核纤层纤维的直径约10 nm,属于中间纤维的一种,其中β亚基与内核膜的特异受体蛋白相结合,α、γ亚单位与β相连接,而α、γ又同染色质的特定部分相结合。
(整理)细胞核与染色体

细胞核与染色体学习方法归纳:第一、认识细胞生物学课程的重要性,正如原子是物理性质的最小单位,分子是化学性质的最小单位,细胞是生命的基本单位。
50年代以来诺贝尔生理与医学奖大都授予了从事细胞生物学研究的科学家,可见细胞生物学的重要性。
如果你将来打算从事生物学相关的工作,学好细胞生物学能加深你对生命的理解。
第二、明确细胞生物学的研究内容,即:结构、功能、生活史。
生物的结构与功能是相适应的,每一种结构都有特定的功能,每一种功能的实现都需要特定的物质基础。
如肌肉可以收缩、那么动力是谁提供的、能量从何而来的?第三、从显微、超微和分子三个层次来认识细胞的结构与功能。
一方面每一个层次的结构都有特定的功能,另一方面各层次之间是有机地联系在一起的。
第四、将所学过的知识关联起来,多问自己几个为什么。
细胞生物学涉及分子生物学、生物化学、遗传学、生理学等几乎所有生物系学过的课程,将学过的知识与细胞生物学课程中讲到的内容关联起来,比较一下有什么不同,有什么相同,为什么?尽可能形成对细胞和生命的完整印象,不要只见树木不见森林。
另一方面细胞生物学各章节之间的内容是相互关联的,如我们在学习线粒体与叶绿体的时候,要联想起细胞物质运输章节中学过的DNP、FCCP 等质子载体对线粒体会有什么影响,学习微管结构时要问问为什么β微管蛋白是一种G蛋白,而α微管蛋白不是,学习细胞分裂时要想想细胞骨架在细胞分裂中起什么作用,诸如此类的例子很多。
第五、紧跟学科前沿,当前的热点主要有“信号转导”、“细胞周期调控”、“细胞凋亡”等。
细胞生物学是当今发展最快的学科之一,知识的半衰期很短(可能不足5年),国内教科书由于编撰周期较长,一般滞后于学科实际水平5-10年左右,课本中的很多知识都已是陈旧知识。
有很多办法可以使你紧跟学科前沿:一是选择国外的最新教材,中国图书进出口公司读者服务部那里可以买到很多价廉物美的正宗原版教材(一般200-400元,只相当于国外价格的1/5);二是经常读一些最新的期刊资料,如果条件所限查不到国外资料,可以到中国期刊网、万方数据等数据库中查一些综述文章,这些文章很多是国家自然科学基金支助的,如在中国期刊网的检索栏输入关键词“细胞凋亡”,二次检索输入关键词“进展”,你会发现一大堆这样的文章,都是汉字写的比读英文省事。
细胞生物学第10章2细胞核与染色体

多线染色体
◆存在于双翅目昆虫的幼虫组织细胞、某些植 物细胞
◆多线染色体的来源:核内有丝分裂
◆多线染色体的带及间带:
带和间带都含有基因,可能“管家”基因 (housekeeping gene) 位于间带, “奢侈”基因 (luxury gene)
◆多线染色体与基因活性:胀泡是基因活跃转 录的形态学标志
Structure of the nucleolus
二、核仁的功能
核仁是细胞制造核糖体的装置。 ◆rRNA的合成 ◆rRNA前体的加工 ◆参与核糖体大小亚基的装配 ◆控制蛋白质合成的速度
rRNA基因转录的形态及组织特征
组织特征位于NORs的rDNA是rRNA的信 息来源。
形态特征:“圣诞树”样结构。
内部着丝粒蛋白INCENP(inner centromere
protein) 染色单体连接蛋白clips(chromatid linking proteins) roteins)
着 丝 粒 与 动 粒
染 色 体 主 要 结 构
二.染色体DNA的三种功能元件 (functional elements)
第三节 染色体
●中期染色体的形态结构 ●染色体DNA的三种功能元件(functional elements) ●巨大染色体(giant chromosome)
一.中期染色体的形态结构
中期染色体的典型形态 类型 染色体的主要结构
染 色 体 的 电 镜 照 片
类型
中着丝粒染色体(metacentric chromosome) 近(亚)中着丝粒染色体
活性染色质是具有转录活性的染色质 活性染色质的核小体发生构象改变,具
有疏松的染色质结构,从而便于转录调 控因子与顺式调控元件结合和RNA 聚合 酶在转录模板上滑动。 非活性染色质是没有转录活性的染色质
细胞生物学思考题及答案

第八章细胞信号转导1、名词解释细胞通讯: 指一个细胞发出的信息通过介质传递到另一个细胞并与其受体相互作用,产生特异性生物学效应的过程。
受体: 指能够识别和选择性结合某种配体(信号分子)的大分子。
多数为糖蛋白,少数为糖脂或二者复合物。
第一信使: 由信息细胞释放的,经细胞外液影响和作用其它信息接收细胞的细胞外信号分子第二信使: 第一信使与受体作用后在胞内最早产生的信号分子称为第二信使。
2、细胞信号分子分为哪两类?受体分为哪两类?细胞信号分子:亲脂性信号分子和亲水性信号分子;受体:细胞内受体:位于细胞质基质或核基质,主要识别和结合脂溶性信号分子;细胞表面受体:主要识别和结合亲水性信号分子(三大家族;G 蛋白耦联受体,酶联受体,离子通道耦联受体)3、两类分子开关蛋白的开关机制。
GTPase开关蛋白:结合GTP活化,结合GDP失活。
鸟苷酸交换因子GEF引起GDP从开关蛋白释放,继而结合GTP并引起G蛋白构象改变使其活化;随着结合GTP水解形成GDF和Pi,开关蛋白又恢复成失活的关闭状态。
GTP水解速率被GTPase促进蛋白GAP和G蛋白信号调节子RGS所促进,被鸟苷酸解离抑制物GDI所抑制。
普遍的分子开关蛋白:通过蛋白激酶使靶蛋白磷酸化和蛋白磷酸酶使靶蛋白去磷酸化活性调节蛋白质活性。
4、三类细胞表面受体介导的信号通路各有何特点?(1)离子通道耦联受体介导的信号通路特点:自身为离子通道的受体,有组织分布特异性,主要存在与神经、肌肉等可兴奋细胞,对配体具有特异性选择,其跨膜信号转导无需中间步骤,其信号分子是神经递质。
(2)G蛋白耦联受体介导的信号通路特点:信号需与G蛋白偶联,其受体在膜上具有相同的取向,G蛋白耦联受体一般为7次跨膜蛋白,会产生第二信使,G蛋白在信号转导过程中起着分子开关的作用。
(3)酶连受体信号转导特点:a.不需G蛋白,而是通过受体自身的蛋白酶的活性来完成信号跨膜转换; b.对信号的反应较慢,且需要许多细胞内的转换步骤;c. 通常与细胞生长、分裂、分化、生存相关。
细胞生物学细胞核与染色体

细胞核与细胞质细胞核是真核细胞内最大、最明显和最重要的细胞器。
是区别原核细胞与真核细胞最显著的特征之一。
一般一个细胞只有一个细胞核,但在有些特殊细胞中,有多个细胞核。
细胞核主要由核被膜、核纤层、染色质、核仁及核体组成。
细胞核是遗传信息的储存场所,与细胞遗传及代谢活动密切相关的基因复制、转录和转录初产物的加工过程均在此进行。
核被膜核被膜的形态结构核被膜是包围在细胞核外的界膜,核被膜含有两层核膜,内层核膜的内表面存在一层由中间丝相互交织成的搞电子密度的蛋白质网络结构,为核纤层。
核被膜的外核膜外表面结合有核糖体。
内外核膜之间隔有间隙,为核间隙。
在核膜的许多部位,内外核膜相互融合,成为通道,为核孔。
每一核空由一个极为精密复杂的结构所组成,此结构为核孔复合体。
核被膜是有内外两层大致平行的膜组成,向着胞质侧的一层核膜称为外核膜,常常与糙面内质网相连,其胞质面上附有大量的核糖体。
近核质一侧核膜为内核膜,其内表面光滑,含有一些特异的蛋白质。
内外核膜之间存在间隙,与糙面内质网腔相通。
有贯穿核被膜的细胞质和核质间的环形通道为核空。
靠近核孔的核膜在化学组成上与其它处的核膜不同,特称核孔区,其特征蛋白为一种跨膜糖蛋白gp210.核被膜的功能及生物学意义一方面,核被膜构成了核、质之间的天然选择性屏障,将细胞分成核质结构和功能区域,使得DNA复制,RNA转录在核内进行。
而蛋白质的翻译则局限在细胞质中。
这样既避免了核质间彼此相互干扰,使细胞的生命活动秩序更加井然。
同时还能保护核内的DNA分子免受损伤。
另一方面,核被膜调控细胞核内外的物质交换和信息交流。
核被膜并不是完全封闭的,核质之间进行着频繁的物质交换和信息交流。
这些物质交换与信息交流主要是通过核被膜上的核孔复合体进行的。
核孔复合体的结构核孔是胞质与核质之间物质交换的通道,每一核孔都是由结构精密的核孔复合体构成,组成核孔复合体的蛋白叫核孔蛋白,核孔复合体的数量随细胞种类、转录活性不同而有较大差异。
细胞生物学 第10章 细胞核与染色体

可变的连接组蛋白(linker histone)即H1。
H1是多样性,具有属(genus)和组织特异性
染色质中的组蛋白与DNA的含量之比为:1∶1。
2. 组蛋白
(2) 功能
核小体组蛋白作用是与DNA组装成核小体
H1不参加核小体的组建, 在构成核小体时起连
接作用,并赋予染色质以极性。
3. 非组蛋白
三、染色质包装的结构模型
(一)染色质包装的多级螺旋模型(multiple coiling model)
但是在电镜下观察用温和方法分离的染色质是直径30nm的
纤维,这种纤维的形成有两种解释:①由核小体螺旋化形
成,每6个核小体绕一圈,长度压缩6倍;②由核小体纤维
Z字形折叠而成,长度压缩40倍。
对运输颗粒大小的限制。
是一个信号识别与载体介导的过程,需消耗
ATP,表现出饱和动力学特征
具有双向性。
爪蟾卵母细胞核质蛋白质注射实验
二、核孔复合体
2. 主动运输 (1) 亲核蛋白运输机制
基本概念
亲核蛋白:在细胞质内合成后,需要或能够进入细胞核 内发挥功能的一类蛋白质 核定位信号(nuclear localization signals,NLS):存在于亲 核蛋白内,具有定向、定位作用的特殊氨基酸序列。 输入蛋白(importin):仅有核定位信号的蛋白质自身不能 通过核孔复合体,它必须与水溶性的NLS受体结合才可 穿过NPC,这种受体称为输入蛋白。
新核膜来自旧核膜������ 核被膜的去组装是非随机的,具有区域特异性 (domain-specific)。������ 核被膜的解体与重建的动态变化受细胞周期调 控因子的调节,调节作用可能与核纤层蛋白、 核孔复合体蛋白的磷酸化与去磷酸化修饰有关。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第十章细胞核与染色体一、间期核的性质(一)形状:一般来说,间期核的形状是与细胞形状相对应的。
当细胞呈等直径形(圆球形、立方形、对称多角形),核呈圆形;当细胞呈长形(柱状、管状、棱状)核则呈椭圆形;当细胞是扁平状,核呈扁盘形。
另外,亦有细胞核呈不规则形的,例如:白血细胞(核呈多叶形),纤毛虫(核呈链珠形),蚕丝腺细胞(核呈分枝形),胚乳细胞(核呈网状)。
(二)大小:一般来说,间期核的体积与细胞体积成正比关系,但不同发育时期也有变化。
(三)数量:通常细胞中都是单核,但也有双核或多核的。
例如。
乳管细胞(菊科植物)及骨藻细胞中,核有几百个。
动物横纹肌细胞及骨骼内的破骨细胞中,核也达一百个左右。
这些多核细胞是由于核分裂次数多于胞质分裂次数所导致的,或者是由于天然发生的细胞融合所造成的(合胞体)。
此外,还有少数类型细胞是无细胞核的,例如人的成熟红细胞及植物的成熟筛管细胞,皆是由于细胞分化而导致丧失了核,故再不能分裂增殖了,寿命亦十分有限。
(四)位置:胚胎细胞和幼龄细胞内,细胞核居中,但随着细胞生长和分化,有时核会移位和变形。
例如成熟的植物细胞之中,细胞核常被中央液泡挤到一侧边位置。
二、间期核的结构(一)核膜nuclear envelope1.形态结构:电镜下观察,核膜是由两层平行排列的单位膜组成,即核外膜和核内膜,每层膜的厚度约7.5nm,在内、外膜之间有宽为20-50nm的间隙,称为核周隙(perinuclearspace)。
核外膜的外表面附有核糖体,其部分区域与糙面内质网膜相连,∴核周隙与内质网腔是连通的。
核内膜上无核糖体附着,其内侧有一层纤维网状结构。
称为核纤层nuclear lamina,核纤层的厚度因细胞而异,一般在30nm以下,组成核纤层的蛋白纤维是由3种多肽——核纤层蛋白A、B、C(MW60—75KD)装配而成,这种纤维可与核内膜中的laminB受体结合,又可与染色质的特定区段(异染色质)连接,∴核纤层是维系核膜及染色质的结构支架。
核膜上具有能贯通内外的孔道,称为核孔nulear pore,核孔在核膜上的数量和密度因细胞类型和生理状态而异,凡代谢旺盛、转录活跃的细胞则核孔多而密。
核孔中有复杂结构,故统称为核孔复合体nuclear pore complex,动、植物细胞核膜上都具有此结构。
其具体构型为:在核孔外缘和内缘各有一胞质环和核质环,由这两环分别朝核内外各伸出8条纤丝,胞质纤丝短而卷曲,核质纤丝细长伸入核内,末端还形成一小环(由8个颗粒组成),型似捕鱼笼。
此外,在核孔复合体内部又有一平面对称分布的8个颗粒及1个中央颗粒(或称中央栓),这些结构物皆是核糖核蛋白构成。
总之,核孔复合体的基本结构特点是:对垂直于核膜的中心轴是呈八重对称分布格局,而对核膜内外则是不对称分布。
应用电镜免疫测定,核孔复合体的标志蛋白是gp210(跨膜糖蛋白),是起锚定核孔复合体作用。
另外,中央颗粒上还有一种P62蛋白。
从酵母到人,各类生物细胞的核孔蛋白都具有同源性,故核孔复合体在进化上是高度保守的。
2、核膜的主要功能(1)是核内外隔离屏障,使细胞核成为相对独立、稳定的生理功能系统。
核内的渗透压、pH值,电位差、化学成分和电磁效应,均有别于细胞质且维持相对恒定,因而细胞核内的生理生化活动能实现专门化。
该屏障功能,除依靠核内、外膜之外,还有核周隙和核纤层的作用不应忽视,核周隙是介于核质与胞质之间的生理缓冲区;核纤层能配合核膜维持核的完整性,核纤层蛋白对分裂时的核膜解体以及分裂后的核膜重建起重要调控作用。
(2)控制核内外的物质和信息交换。
以核孔复合体通道进行的双向选择性物质运输方式有两种:被动扩散和主动运输。
经微量注射胶体金测试,核孔通道有效直径约9nm,离子、小分子代谢物皆由此通道可进行自由扩散,而大分子物质则需主动运输。
但也有些直径小于9nm的物质并不能自由扩散过膜,反而直径达26 nm的物质经主动运输却顺利通过,这是因为核孔复合体中的有效通道直径会自行调节,能有选择性地控制穿过核孔的物质双向运输。
例如:输出核外的物质,有在核内组装的核糖体亚单位(RNP 颗粒)、mRNA和tRNA前体等。
在核内加工完毕的mRNA可通过核孔输到细胞质中去,而核内不均一RNA(hnRNA,即mRNA的前体)却不能穿过,这是由于mRNA的5’端加上了m7GPPPG帽子的选择信号。
另外,还有些蛋白具有核输出信号NES序列。
再例如输入核内的物质,有在细胞质中合成的DNA聚合酶、RNA聚合酶,以及组装染色质的组蛋白和非组蛋白,组装核糖体的r蛋白,还有激素受体复合体蛋白等亲核蛋白质,它们之所以都能被选择性地主动运输进入核内,是由于皆含有一段核定位信号nuclear localization signal,NLS序列,这是定向进入核孔和在核内定位的信号序列。
NLS序列与信号肽及导肽不相同,它可存在亲核蛋白质的各种部位,并且在引导蛋白质输入后不会被切除,NLS 序列是被核孔复合体中的NBP s受体蛋白所识别、结合及介导穿过核孔的。
总之,核孔复合体上主动运输的特点可归纳为:①信号识别;②载体介导;③需GTP供能;④双向选择;⑤具饱合动力学规律。
(3)蛋白质合成作用:核外膜外表附着有核糖体,其合成产物是由核周隙与内质网相连通的管道输走。
(二)核仁nucleolus核仁是间期核中的重要结构之一。
其形状、数目和大小在不同生物细胞中表现不同。
一般是圆球形,也有呈不规则形的;有的是单个,也有的呈多个的;凡蛋白质合成旺盛的细胞中核仁明显偏大,而蛋白质合成差的细胞中核仁小。
此外,核仁在细胞周期过程中表现出周期性的消失和重现规律,这是进行rRNA合成、加工及核糖体亚单位装配的动态表现。
核仁中,蛋白质占80%,RNA占10—15%,另外还有5—10%的DNA和微量脂类。
核仁无膜包围,其内可大致分三种区域:纤维中心FC,致密纤维组分DFC,颗粒组分GC。
纤维中心区域有10nm粗的纤丝,是含有rRNA基因的染色质纤丝。
在此区域外围的致密纤维组分区域由大量2—5nm的细纤丝交织成海绵状,这些细纤丝是结合了蛋白质的45S rRNA前体。
而在颗粒组成区域则含有许多直径10—20nm的颗粒,这是正在装配中的成熟程度不同的核糖体亚单位,主要是大亚单位。
细纤丝和颗粒皆是核糖核蛋白RNP s,除去这些结构物剩下的是以可溶性蛋白质组成的核仁基质。
核仁组织区nucleolus organizing region ,NOR,是指某一对(或某几对)同源染色体上的一种特殊区域,含有许多rRNA基因拷贝。
在间期核中,染色体解螺旋时,NOR区域的染色质纤丝就是插入核仁中纤维中心区的10nm粗纤丝;而到分裂期,核仁中的染色质纤丝螺旋化再重新形成染色体上的NOR区段。
有些生物的NOR是位于染色体的次缢痕,例如玉米的NOR在第6对染色体短臂的次缢痕上,人的NOR 在第13、14、15、21、22这五对染色体的次缢痕处(不过,也有部分生物的NOR不在染色体次缢痕位置)。
具有NOR 的染色体可统称为核仁染色体。
间期核内的核仁之所以明显,是由于其中编码rRNA 基因的10nm粗的染色质纤丝解螺旋,正在活跃转录,指导核仁内的rRNA合成和加工以及核糖体大、小亚单位的装配。
而分裂期时,rRNA转录停止,那段10nm粗的染色质纤丝螺旋化卷到核仁染色体上去,并且原已合成及装配的核糖体亚单位都从核孔输出到细胞质中去了,∴核仁变小而消失。
核仁的主要功能:1)是rRNA前体的转录合成及加工场所;2)是核糖体亚单位的组装场所。
以染色质铺展技术在电镜下观察rDNA转录情况(非洲爪蟾、蝾螈、伞藻等的核仁中):伸展的DNA纤维上间隔分布有若干段由平行细丝组成“圣诞树”状结构,这每一段即是rDNA的一个转录单位,而那些细丝即是由RNA聚合酶Ⅰ附着在DNA上的新转录合成rRNA链,RNA聚合酶Ⅰ从基因起始端开始边读码转录边向基因终止端移动(每段上约有100—200个RNA聚合酶Ⅰ在依次移动工作)因此形成顺转录方向细丝逐渐增长的“圣诞树”。
在RNA细丝的游离端可见球状的RNP颗粒。
在基因终止端已完成转录的RNA细丝即脱离DNA链,游离至核仁基质中去。
rRNA基因属于中等重复序列DNA,非洲爪蟾的含600个拷贝,人的有200个拷贝,这种串联重复排列的rRNA基因,可保证RNA聚合酶能连续高效大量转录,以便能大量装配核糖体。
这是因为每个正常细胞都必需有107个核糖体才能确保其蛋白质合成的顺利运行。
注意,上述在核仁中的rRNA基因转录单位,仅包含18S、5.8S和28S rRNA,而5S rRNA基因不在NORs区域,其转录亦不在核仁之中。
人的细胞中约有2000个5S rRNA基因拷贝,也是成簇串联排列,由RNA聚合酶Ⅲ转录,转录后经加工进入核仁颗粒区参与核糖体大亚单位的组装。
rRNA前体(45S)合成后,须经过一系列的加工修饰(甲基化、切割降解),才能分成18S、5.8S和28S。
注意此加工的对象都不是游离状态的rRNA,而是RNP复合物,经过多步骤剪切,加工组装成核糖体的大、小亚单位,由核孔复合体输出到细胞质中去,小亚单位组装快,而大亚单位组装较缓慢。
∴颗粒区所见的颗粒主要是大亚单位。
(三)染色质chromatin1、概念及类型染色质:是分裂期染色体的组成物质,也是间期核内能被碱性染料(即胞核染料,如:苏木精、卡红、甲绿、碱性品红等)染色的物质。
是由DNA、组蛋白、非组蛋白以及少量的RNA所组成的串珠状复合物。
通常分为两大类型:常染色质euchromatin是在分裂间期能正常解螺旋,而被染色较淡的染色质部分。
这种状况是其中的基因能进行转录的必要条件。
异染色质heterochromatin是在分裂间期保持凝缩状态而被浓染的染色质部分。
无转录活性,其DNA复制行为比常染色质的复制较晚,而凝缩较早。
可分为两种类型:结构异染色质constitutive heterchromatin,是在各种细胞类型和发育阶段都呈凝缩状态的异染色质,分布于染色体的着丝粒区、端粒区、次缢痕及某些染色体的特定区段。
兼性异染色质facultative heterochromatin,是在某种特定组织细胞里或某个发育阶段,由常染色质转变成的异染色质,其丧失了转录活性。
例如:正常女性的体细胞中两条X染色体的染色质原本都是具转录活性的常染色质(追溯其来源分别是来自父母双亲),但受精后16—18天时,其中一条随机性失活凝缩为兼性异染色质,即成为体细胞间期核中呈现的巴氏小体。
Lyon 理论(1960年)解释,X染色质失活是剂量补偿效应。
活性染色质:具转录活性的常染色质。
非活性染色质:是不转录的染色质,既包括异染色质,也包括基因处于暂时关闭状态的常染色质。