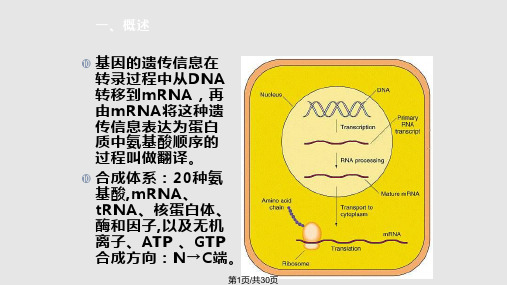
蛋白质生物合成讲课
合集下载
蛋白质的生物合成ppt课件

mRNA
密码子
t R N A 反 密 码 碱 基I
U
C
m R N A 密 码 碱 基 A , C , U A , G C , G , U
通用性
除了动物细胞中的线粒体和植物细胞中的叶绿体外, 从最简单的病毒、原核生物到人类都使用一套遗传密 码。
二、核糖体是肽链合成的场所
• 氨基酸首先在 核糖体内合成 蛋白质,再输 送至细胞其它 组分中。
三、肽链合成的终止
肽链合成终止包括: • 终止密码的辨认 • 肽链从肽-tRNA上水解 • mRNA从核糖体中分离 • 核糖体大小亚基的拆分
三、肽链合成的终止
• 释放因子RF: 终止过程中的蛋白质因子 辨认终止密码 促进肽链C端与tRNA 3’-OH酯键的水解, 使肽链从翻译中的核糖体上释放下来。
GCA CAG
密码之间没有 核苷酸间断
AGU
GUA
………….
• 除了色 氨酸和 蛋氨酸 外,其 余氨基 酸均有2 -6个三 联体为 其编码。
简并性
*
摆动性
• 密码与反密码配对辨认 时,有时并不完全按照 碱基互补规律。尤其是 密码的第三碱基对反密 码的第一位碱基,更常 出现这种摆动现象。
C-G-I 反密码子 G-C-C 密码子
氯霉素
• 能与原核生物的核糖体大亚基结合,抑 制转肽酶活性而阻断翻译的延长过程。
链霉素和卡那霉素
• 能与原核生物核糖体小亚基结合,改变 其构象,引起读码错误,使毒素类的细 菌蛋白失活。
• 结核杆菌敏感
嘌呤霉素
• 结构与酪氨酸-tRNA相似,从而取代一些 氨基酰-tRNA进入翻译进入翻译中的核糖 体的A位。
• 抗生素一般是细菌或真菌所产生的具有 抑制其它生物生长的物质。
蛋白质的生物合成课堂PPT

– GTP酶部位:GTP酶也称转位酶,简称G因子,能分解GTP分子,并将肽酰基tRNA由A部位移到P部位。
返回
tRNA的活化: 氨基酰-tRNA 合成酶催化tRNA与特定氨基酸结合
返回
tRNA的活化
• 氨基酸 + ATP + tRNA + H2O
氨基酰-tRNA + AMP + PPi
• 每一种氨基酸至少有一种对应的氨基酰-tRNA 合成酶。它既催
蛋白质
– A部位:也称受位,主要位于大亚基上,是接受氨酰基核-tR糖N体A的的部功位能。活性部位图示
– P部位:也称肽酰基部位或供位,主要在小亚基上,是肽酰基-tRNA移交新生肽链 的部位。
– E部位:也称退出位点,位于小亚基,是失去氨基酰的tRNA从核糖体退出的地方
– 肽基转移酶部位:肽基转移酶也称肽合成酶,简称T因子,位于大亚基上,其作用 是在肽链合成过程中催化氨基酸与氨基酸间形成肽键。
遗传密码的破译
• 第二个实验(核糖体结合技术)是1964年也是由美国的M.Nirenberg 等人完成的。即是用一个人工合成的三核苷酸代替mRNA,在含有核 糖体和 14C-氨基酰-tRNA的缓冲系统中,无需酶促核糖体便可以和带 有特定氨基酰-tRNA及三核苷酸结合,例如三核苷酸为pUpUpU时, 则只有14C-苯丙氨酰-tRNA结合到核体上。用硝酸纤维制成的滤膜过 滤,结合有氨基酰-tRNA的核糖体保留在滤膜上,未结合的氨基酰- tRNA被洗脱,测定滤膜上的放射性,就可知道和核糖体上的三核苷 酸结合的是哪一种氨基酸。
AA
A G
AGU
GUC
CAU
CAC
UUA
AUG GC
酸推测)
第十章蛋白质的生物合成PPT课件
第15页/共30页
肽链的延长
进位 (氨酰tRNA进入A位点) 参与因子:延长因子EFTu(Tu)、EFTs(Ts)、GTP、氨酰tRNA 肽链的形成 肽酰基从P位点转移到A位点,形成新的肽链 移位(translocase) 在移位因子(移位酶)EF-G的作用下,核糖体沿mRNA(5’-3’) 作相对移动,使原来在A位点的肽酰-tRNA回到P位点
第2页/共30页
第3页/共30页
原核生物 核糖体组 成
真核生物 核糖体组 成
第4页/共30页
2. tRNA
结合氨基酸:一种氨基 酸有几种tRNA携带, 结合需要ATP供能,氨 基酸结合在tRNA3‘CCA的位置。
反密码子:每种tRNA的 反密码子,决定了所带 氨基酸能准确的在 mRNA上对号入座 。 反密码子与mRNA的第 三个核苷酸配对时,不 严格遵从碱基配对原则
三、遗传密码子
• 为一个氨基酸编码进入蛋白质多肽链特定线性 位置的三个核苷酸单位称为密码子(Coden) 或三联体密码。
• 密码子的发现
• 统计学方法 • 人工合成仅由一种核苷酸组成的多聚核苷酸,推测由
哪一种氨基酸合成的多肽 • 核糖体结合试验 1965年,Nirenberg用poly u加入
C14标记的20种aa,仅有苯丙氨酸的寡肽,UUU=苯丙 氨酸,用此法破译了全部密码,编出遗传密码表。
第22页/共30页
五、真核细胞蛋白质合成的特点
核糖体为80S,由60S的大亚基和40S的小亚基组成 起始密码AUG 起始tRNA为Met-tRNA 起始复合物结合在mRNA 5’端AUG上游的帽子结构,真核mRNA无富含嘌
呤的SD序列(除某些病毒mRNA外) 已发现的真核起始因子有近9种(eukaryote Initiation factor,eIF) 复合物称
肽链的延长
进位 (氨酰tRNA进入A位点) 参与因子:延长因子EFTu(Tu)、EFTs(Ts)、GTP、氨酰tRNA 肽链的形成 肽酰基从P位点转移到A位点,形成新的肽链 移位(translocase) 在移位因子(移位酶)EF-G的作用下,核糖体沿mRNA(5’-3’) 作相对移动,使原来在A位点的肽酰-tRNA回到P位点
第2页/共30页
第3页/共30页
原核生物 核糖体组 成
真核生物 核糖体组 成
第4页/共30页
2. tRNA
结合氨基酸:一种氨基 酸有几种tRNA携带, 结合需要ATP供能,氨 基酸结合在tRNA3‘CCA的位置。
反密码子:每种tRNA的 反密码子,决定了所带 氨基酸能准确的在 mRNA上对号入座 。 反密码子与mRNA的第 三个核苷酸配对时,不 严格遵从碱基配对原则
三、遗传密码子
• 为一个氨基酸编码进入蛋白质多肽链特定线性 位置的三个核苷酸单位称为密码子(Coden) 或三联体密码。
• 密码子的发现
• 统计学方法 • 人工合成仅由一种核苷酸组成的多聚核苷酸,推测由
哪一种氨基酸合成的多肽 • 核糖体结合试验 1965年,Nirenberg用poly u加入
C14标记的20种aa,仅有苯丙氨酸的寡肽,UUU=苯丙 氨酸,用此法破译了全部密码,编出遗传密码表。
第22页/共30页
五、真核细胞蛋白质合成的特点
核糖体为80S,由60S的大亚基和40S的小亚基组成 起始密码AUG 起始tRNA为Met-tRNA 起始复合物结合在mRNA 5’端AUG上游的帽子结构,真核mRNA无富含嘌
呤的SD序列(除某些病毒mRNA外) 已发现的真核起始因子有近9种(eukaryote Initiation factor,eIF) 复合物称
第十一章蛋白质的生物合成ppt课件

一、翻译模板mRNA及遗传密码
(一) mRNA是遗传信息的携带者 遗传学将编码一个多肽的遗传单位称为顺
反子(cistron)。 原核细胞中数个结构基因常串联为一个转
录单位,转录生成的mRNA可编码几种功 能相关的蛋白质,为多顺反子(polycistron)。 真核mRNA只编码一种蛋白质,为单顺反 子(single cistron) 。
翻译的延长(elongation)
翻译的终止(termination )
一、原核生物的翻译过程
(一) 起始阶段
1.核蛋白体大小 30S 亚基分离; 50S
2.mRNA与小亚
IF-3
IF-1
30S + 50S
解聚
mRNA
基结合; 3.起始氨基酰-
IF-2,GTP
AUG
tRNA与小亚基
fMet
fMet
溶酶体蛋白 Man-6-P(甘露糖-6-磷酸)
分泌蛋白的靶向输送
真核细胞分泌蛋白等前体合成后靶向输送过 程首先要进入内质网。
信号肽(signal peptide)--
各种新生分泌蛋白的N端有保守的氨基酸序 列称信号肽。
信号肽的一级结构
信号肽引导真核分泌蛋白进入内质网
EF-Ts 调节亚基
EF-1-βγ
有转位酶活性,促进mRNA-肽 EFG 酰-tRNA由A位前移到P位,促
进卸载tRNA释放
EF-2
1.进位
又称注册(registration)
指根据mRNA下 一组遗传密码指导, 使相应氨基酰-tRNA 进入核蛋白体A位。
Tu TGsTP
Tu GDP
Ts GTP
遗 传 密 码 表
目录
从mRNA 5端起始密码子AUG到3端终止密码 子之间的核苷酸序列,各个三联体密码连续排列编 码 一 个蛋 白 质多 肽 链 , 称 为开 放 阅读 框 架 (open reading frame, ORF)。
蛋白质生物合成PPT课件演示教学.ppt

缬 脯 苏 天冬
缬 丙 酪 甘
缬 丙 丝 精
3. 简并性(degeneracy)
1. 核糖体大小亚基分离; 2. 核糖体小亚基结合于mRNA的起始密码子附近; 3. fMet-tRNAfMet结合在核糖体P位 ; 4. 核糖体大亚基结合形成起始复合物。
一、翻译起始复合物的装配启动肽链合成
(a)起始复合物的装配过程;(b)rRNA识别mRNA的核糖体结合位点,保证翻译起始在起始密码子处
密码子(codon)
起始密码子和终止密码子:
遗传密码表
遗传密码的特点
1. 方向性(directional)
翻译时遗传密码的阅读方向是5→3,即读码从mRNA的起始密码子AUG开始,按5→3的方向逐一阅读,直至终止密码子。
N
C
肽链延伸方向
5
3
读码方向
2. 连续性(non-punctuated)
23S-rRNA 5S-rRNA
18S-rRNA
28S-rRNA 5.8S-rRNA 5S-rRNA
蛋白质
rpS 21种
rpL 36种
rpS 33种
rpL 49种
不同细胞核蛋白体的组成
核蛋白体的组成
核糖体在翻译中的功能部位
四、肽链生物合成需要酶类和 蛋白质因子
氨基酰-tRNA合成酶(aminoacyltRNA synthetase),催化氨基酸的活化; 转肽酶(peptidase),催化核蛋白体P位上的肽酰基转移至A位氨基酰-tRNA的氨基上,使酰基与氨基结合形成肽键;并受释放因子的作用后发生变构,表现出酯酶的水解活性,使P位上的肽链与tRNA分离; 转位酶(translocase),催化核蛋白体向mRNA3’-端移动一个密码子的距离,使下一个密码子定位于A位。
缬 丙 酪 甘
缬 丙 丝 精
3. 简并性(degeneracy)
1. 核糖体大小亚基分离; 2. 核糖体小亚基结合于mRNA的起始密码子附近; 3. fMet-tRNAfMet结合在核糖体P位 ; 4. 核糖体大亚基结合形成起始复合物。
一、翻译起始复合物的装配启动肽链合成
(a)起始复合物的装配过程;(b)rRNA识别mRNA的核糖体结合位点,保证翻译起始在起始密码子处
密码子(codon)
起始密码子和终止密码子:
遗传密码表
遗传密码的特点
1. 方向性(directional)
翻译时遗传密码的阅读方向是5→3,即读码从mRNA的起始密码子AUG开始,按5→3的方向逐一阅读,直至终止密码子。
N
C
肽链延伸方向
5
3
读码方向
2. 连续性(non-punctuated)
23S-rRNA 5S-rRNA
18S-rRNA
28S-rRNA 5.8S-rRNA 5S-rRNA
蛋白质
rpS 21种
rpL 36种
rpS 33种
rpL 49种
不同细胞核蛋白体的组成
核蛋白体的组成
核糖体在翻译中的功能部位
四、肽链生物合成需要酶类和 蛋白质因子
氨基酰-tRNA合成酶(aminoacyltRNA synthetase),催化氨基酸的活化; 转肽酶(peptidase),催化核蛋白体P位上的肽酰基转移至A位氨基酰-tRNA的氨基上,使酰基与氨基结合形成肽键;并受释放因子的作用后发生变构,表现出酯酶的水解活性,使P位上的肽链与tRNA分离; 转位酶(translocase),催化核蛋白体向mRNA3’-端移动一个密码子的距离,使下一个密码子定位于A位。
蛋白质的生物合成课件.ppt

如何转变?
密码子:mRNA分子中,每三个相邻的核苷酸组
成的三联体代表某一种氨基酸或其它信息,称为 密码子或三联密码.一个氨基酸密码子决定着一 个氨基酸。
遗传密码:mRNA中的核苷酸排列序列与蛋白
质中的氨基酸排列序列的关系。生物的遗传密码
是通用的。
四种核苷酸编成三联体可形成 43个即64个密码子.其中: 1.一个起始密码:AUG
小亚基:沿mRNA结合,沿5’ 3’ 方向移动.
大亚基:受位(A位): 结合氨基酰- tRNA
给位(P位):成肽
给位 (P位)
蛋 苏
大亚基
UGU
5’AUG ACA GUU
受位 (A位)
小亚基
3’
蛋白质生物合成过程
1、准备阶段: 氨基酸的活化与转运。
2、中心环节: 核蛋白体循环-活化氨基酸 在核蛋白体上的缩合组装。
氨基酸的活化与转运
1、反应式:
氨基酰-tRNA合成酶
AA + tRNA + ATP
氨基酰-tRNA+AMP+PPi 2、AA结合位置:
AA的α-羧基与tRNA活末端腺苷酸中 核糖2 ’或3’羟基以酯键相结合。
tRNA-CCA-OH(R-3’-OH)
核蛋白体循环(三阶段)
(1)、起始阶段 (2)、延伸阶段 (3)、终止阶段
基因操纵子调节系统示意图
调节基因 转录
操纵子
控制区
信息区
启动基因 操纵基因 RNA聚合酶
结构基因
DNA
(-)
(+) 转录
翻译
mRNA
阻遏蛋白
诱导剂
翻译 蛋白质
血红素对起始因子-2的调节作用
血红素
密码子:mRNA分子中,每三个相邻的核苷酸组
成的三联体代表某一种氨基酸或其它信息,称为 密码子或三联密码.一个氨基酸密码子决定着一 个氨基酸。
遗传密码:mRNA中的核苷酸排列序列与蛋白
质中的氨基酸排列序列的关系。生物的遗传密码
是通用的。
四种核苷酸编成三联体可形成 43个即64个密码子.其中: 1.一个起始密码:AUG
小亚基:沿mRNA结合,沿5’ 3’ 方向移动.
大亚基:受位(A位): 结合氨基酰- tRNA
给位(P位):成肽
给位 (P位)
蛋 苏
大亚基
UGU
5’AUG ACA GUU
受位 (A位)
小亚基
3’
蛋白质生物合成过程
1、准备阶段: 氨基酸的活化与转运。
2、中心环节: 核蛋白体循环-活化氨基酸 在核蛋白体上的缩合组装。
氨基酸的活化与转运
1、反应式:
氨基酰-tRNA合成酶
AA + tRNA + ATP
氨基酰-tRNA+AMP+PPi 2、AA结合位置:
AA的α-羧基与tRNA活末端腺苷酸中 核糖2 ’或3’羟基以酯键相结合。
tRNA-CCA-OH(R-3’-OH)
核蛋白体循环(三阶段)
(1)、起始阶段 (2)、延伸阶段 (3)、终止阶段
基因操纵子调节系统示意图
调节基因 转录
操纵子
控制区
信息区
启动基因 操纵基因 RNA聚合酶
结构基因
DNA
(-)
(+) 转录
翻译
mRNA
阻遏蛋白
诱导剂
翻译 蛋白质
血红素对起始因子-2的调节作用
血红素
蛋白质的生物合成医学课件
蛋白质生物合成的过程包括起始、延长、终止和翻译后修饰四个阶段。
起始阶段:核糖体与mRNA、tRNA结合形成翻译起始复合物,然后通过一系列构象变化形成翻译延长复合物。
延长阶段:核糖体沿着mRNA移动,通过进位、成肽、转位等步骤循环进行,最终合成完整的多肽链。
终止阶段:当核糖体移动到mRNA的终止密码子时,多肽链合成停止,核糖体释放出多肽链并从mRNA上脱落。
恶性肿瘤
蛋白质生物合成异常与神经退行性疾病如帕金森病、阿尔茨海默病等的发生发展密切相关。
神经退行性疾病
抗生素和抗病毒药物
抗肿瘤药物
神经保护药物
蛋白质生物合成与药物发现及开发
蛋白质生物合成是分子生物学研究的核心内容之一,其研究方法如基因克隆、RNA干扰等技术为医学研究提供了重要手段。
蛋白质生物合成的研究方法在医学中的应用
DNA转录遵循中心法则,即遗传信息从DNA传递给RNA,再由RNA传递给蛋白质,最终表达为特定的生物学功能。
DNA转录的机制
DNA双链在转录开始前需要解旋,解旋后的单链DNA才能作为模板进行转录。
解旋
转录的启动需要一个起始序列,称为启动子,它与RNA聚合酶结合,启动转录过程。
启动
RNA聚合酶沿着DNA模板链向前移动,逐个将核苷酸添加到新生RNA链的3'端,直到遇到终止信号为止。
延伸
当RNA聚合酶遇到终止序列时,转录停止,新生RNA链从DNA模板链上释放出来。
终止
DNA转录的调节
DNA上的某些特定序列可以与调节蛋白结合,影响RNA聚合酶的结合和转录效率。
顺式作用元件
反式作用因子
代谢调节
细胞信号调节
某些调节蛋白可以与RNA聚合酶直接结合,影响其活性或招募其他调节蛋白参与转录过程。
蛋白质的生物合成(共74张PPT)
3)校正tRNA
tRNASu3+Tyr反密码子为CUA,能识别变异 产生的终止密码子UAG。
三、氨酰-tRNA合成酶
❖ 氨基酸在掺入肽链前,必须活化生成氨酰tRNA,获得足够的能量。
❖ 活化反应由各种氨酰-tRNA合成酶(AARS) 催化。
A.氨酰-tRNA
氨基酰-tRNA合成酶
氨基酸 + tRNA
❖ 包括:氨酰-tRNA与核糖体结合,转肽 与肽键形成和转位三个步骤。
肽链合成的延长因子
(一) 进位
Tu GTsTP
Tu GDP
Ts
GTP
5'
AUG
3'
目录
Ts 移走GDP,并与Tu 结合生成Tu-Ts复合体, 然后GTP替换Ts,生 成有活性的Tu-GTP
Poly(U) Poly(C) Poly(A) Poly(G)
poly(Phe) peptide
无细胞抽取物
poly(Pro) peptide poly(Lys) peptide poly(Gly) peptide
•核糖体 •各种tRNA •氨基酸 •AARS
•ATP, GTP
+ mRNA = 蛋白质
氨基酰- tRNA
ATP
AMP+PPi
B. 氨酰-tRNA合成酶特点
a、专一性:
•对氨基酸有极高的专一性,每种氨基酸都有专 一的酶,只作用于L-氨基酸,不作用于D-氨基酸。
•对tRNA具有极高专一性:一种AARS只能识别装
载同一种氨基酸的一组同工受体(tRNA分子)
b、校对作用:
• AARS具有底物结合位点(包括tRNA、氨 基酸和ATP结合位点)和水解位点。
5'
tRNASu3+Tyr反密码子为CUA,能识别变异 产生的终止密码子UAG。
三、氨酰-tRNA合成酶
❖ 氨基酸在掺入肽链前,必须活化生成氨酰tRNA,获得足够的能量。
❖ 活化反应由各种氨酰-tRNA合成酶(AARS) 催化。
A.氨酰-tRNA
氨基酰-tRNA合成酶
氨基酸 + tRNA
❖ 包括:氨酰-tRNA与核糖体结合,转肽 与肽键形成和转位三个步骤。
肽链合成的延长因子
(一) 进位
Tu GTsTP
Tu GDP
Ts
GTP
5'
AUG
3'
目录
Ts 移走GDP,并与Tu 结合生成Tu-Ts复合体, 然后GTP替换Ts,生 成有活性的Tu-GTP
Poly(U) Poly(C) Poly(A) Poly(G)
poly(Phe) peptide
无细胞抽取物
poly(Pro) peptide poly(Lys) peptide poly(Gly) peptide
•核糖体 •各种tRNA •氨基酸 •AARS
•ATP, GTP
+ mRNA = 蛋白质
氨基酰- tRNA
ATP
AMP+PPi
B. 氨酰-tRNA合成酶特点
a、专一性:
•对氨基酸有极高的专一性,每种氨基酸都有专 一的酶,只作用于L-氨基酸,不作用于D-氨基酸。
•对tRNA具有极高专一性:一种AARS只能识别装
载同一种氨基酸的一组同工受体(tRNA分子)
b、校对作用:
• AARS具有底物结合位点(包括tRNA、氨 基酸和ATP结合位点)和水解位点。
5'
蛋白质的生物合成课件(0003)
白
蛋白质生物合成的一般过程(原核) 氨基酸的激活 氨基酸在氨酰-tRNA合成酶的作用下,生成氨酰-
tRNA的过程称为氨基酸的激活 氨酰-tRNA合成酶的专一性 对氨基酸一般有高度专一性 只作用于L-氨基酸 有些对氨基酸专一性不高的酶,对tRNA有极高的
专一性 起始复合体的组装 翻译并不是从mRNA的5’-端第一个核苷酸开始,
嘌呤霉素 链霉素、氯霉素等抗生素 亚胺环己酮 白喉毒素
In bacteria, translation and transcription are tightly coupled
磷酸化 特殊基团修饰 糖基的修饰
辅酶辅基的结合 蛋白原的加工 分子伴侣等辅助折叠
蛋白质翻译的调节
mRNA的调节
mRNA的寿命 mRNA的核输出
核糖体的调节
与mRNA的选择性结合 肽链释放的调节
翻译因子的调节
磷酸化/去磷酸化的平衡 翻译因子的数量调控
蛋白质合成抑制剂
合体相互作用, IF3 释放 50S大亚基与前述复合体结合, IF2催化GTP水解成GDP和磷
酸
IF1.IF2释放 fMet- tRNA处于核糖体的P位点 肽链的延伸
延长因子:EFTu,EFTs的活化 EFTu,EFTs的激活循环 氨酰-tRNA与EFTu的结合
侧链基合成的概述 翻译 定义: 生物体的核糖体以mRNA为模板,利用氨基
酸从头合成蛋白质的过程 合成场所: 真核: 内质网,原核: 拟核区 翻译时读取mRNA上信号的顺序是5’ 3’ 蛋白质合成的方向是N C 核糖体的结构 原核核糖体 直径18nm,含60-65%rRNA,30-35%的蛋白质。 大亚基: 50S,含23 S、5 S rRNA,34 种核糖体蛋
蛋白质生物合成的一般过程(原核) 氨基酸的激活 氨基酸在氨酰-tRNA合成酶的作用下,生成氨酰-
tRNA的过程称为氨基酸的激活 氨酰-tRNA合成酶的专一性 对氨基酸一般有高度专一性 只作用于L-氨基酸 有些对氨基酸专一性不高的酶,对tRNA有极高的
专一性 起始复合体的组装 翻译并不是从mRNA的5’-端第一个核苷酸开始,
嘌呤霉素 链霉素、氯霉素等抗生素 亚胺环己酮 白喉毒素
In bacteria, translation and transcription are tightly coupled
磷酸化 特殊基团修饰 糖基的修饰
辅酶辅基的结合 蛋白原的加工 分子伴侣等辅助折叠
蛋白质翻译的调节
mRNA的调节
mRNA的寿命 mRNA的核输出
核糖体的调节
与mRNA的选择性结合 肽链释放的调节
翻译因子的调节
磷酸化/去磷酸化的平衡 翻译因子的数量调控
蛋白质合成抑制剂
合体相互作用, IF3 释放 50S大亚基与前述复合体结合, IF2催化GTP水解成GDP和磷
酸
IF1.IF2释放 fMet- tRNA处于核糖体的P位点 肽链的延伸
延长因子:EFTu,EFTs的活化 EFTu,EFTs的激活循环 氨酰-tRNA与EFTu的结合
侧链基合成的概述 翻译 定义: 生物体的核糖体以mRNA为模板,利用氨基
酸从头合成蛋白质的过程 合成场所: 真核: 内质网,原核: 拟核区 翻译时读取mRNA上信号的顺序是5’ 3’ 蛋白质合成的方向是N C 核糖体的结构 原核核糖体 直径18nm,含60-65%rRNA,30-35%的蛋白质。 大亚基: 50S,含23 S、5 S rRNA,34 种核糖体蛋
《蛋白质的合成》课件
特异性。
翻译水平的调控
翻译起始
翻译后修饰
翻译起始是蛋白质合成的关键步骤之 一,涉及核糖体与mRNA的结合以及 起始密码子的识别。
翻译后的蛋白质还需要经过一系列的 修饰才能成为有功能的分子,如磷酸 化、乙酰化、糖基化等。
翻译延长与终止
在翻译过程中,核糖体的移位速度和 肽酰-tRNA的形成受到多种因素的调 节,如mRNA的结构、tRNA的浓度 和种类、核糖体的构象等。
04
CATALOGUE
蛋白质合成的应用
蛋白质工程
蛋白质工程是通过修改或设计蛋白质 的氨基酸序列,以达到改善蛋白质的 某些功能或创建新功能的技术。
蛋白质工程在医药、农业和工业领域 有广泛应用,例如设计新的药物、改 良酶的催化效率和稳定性、优化蛋白 质的表达和纯化等。
药物设计与开发
01
蛋白质合成在药物设计与开发中 发挥着关键作用,因为许多药物 是直接与蛋白质相互作用来发挥 其治疗作用的。
蛋白质合成的分子机制
mRNA在蛋白质合成中的作用
信息传递者
mRNA是DNA遗传信息的传递者 ,将遗传信息从DNA传递到核糖 体,用于指导蛋白质的合成。
遗传信息的解码
mRNA上的密码子与tRNA上的反 密码子互补配对,实现遗传信息 的解码,确保氨基酸按照正确的 顺序连接起来。
tRNA在蛋白质合成中的作用
氨基酸转运
tRNA作为氨基酸的转运载体,将氨基酸按照mRNA上的指令准确运送到核糖 体的A位点,参与蛋白质的合成。
反密码子识别
tRNA的反密码子能够与mRNA上的密码子进行互补配对,确保氨基酸按照正确 的顺序加入到多肽链中。
rRNA在蛋白质合成中的作用
核糖体的组成
rRNA是核糖体的主要组成部分,核 糖体是蛋白质合成的场所。
翻译水平的调控
翻译起始
翻译后修饰
翻译起始是蛋白质合成的关键步骤之 一,涉及核糖体与mRNA的结合以及 起始密码子的识别。
翻译后的蛋白质还需要经过一系列的 修饰才能成为有功能的分子,如磷酸 化、乙酰化、糖基化等。
翻译延长与终止
在翻译过程中,核糖体的移位速度和 肽酰-tRNA的形成受到多种因素的调 节,如mRNA的结构、tRNA的浓度 和种类、核糖体的构象等。
04
CATALOGUE
蛋白质合成的应用
蛋白质工程
蛋白质工程是通过修改或设计蛋白质 的氨基酸序列,以达到改善蛋白质的 某些功能或创建新功能的技术。
蛋白质工程在医药、农业和工业领域 有广泛应用,例如设计新的药物、改 良酶的催化效率和稳定性、优化蛋白 质的表达和纯化等。
药物设计与开发
01
蛋白质合成在药物设计与开发中 发挥着关键作用,因为许多药物 是直接与蛋白质相互作用来发挥 其治疗作用的。
蛋白质合成的分子机制
mRNA在蛋白质合成中的作用
信息传递者
mRNA是DNA遗传信息的传递者 ,将遗传信息从DNA传递到核糖 体,用于指导蛋白质的合成。
遗传信息的解码
mRNA上的密码子与tRNA上的反 密码子互补配对,实现遗传信息 的解码,确保氨基酸按照正确的 顺序连接起来。
tRNA在蛋白质合成中的作用
氨基酸转运
tRNA作为氨基酸的转运载体,将氨基酸按照mRNA上的指令准确运送到核糖 体的A位点,参与蛋白质的合成。
反密码子识别
tRNA的反密码子能够与mRNA上的密码子进行互补配对,确保氨基酸按照正确 的顺序加入到多肽链中。
rRNA在蛋白质合成中的作用
核糖体的组成
rRNA是核糖体的主要组成部分,核 糖体是蛋白质合成的场所。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第二步,天然的两条链合成成功之后,再用一个天然链与 一个人工合成的链进行合成,这就是所谓的半合成。最初 用人工合成的B链同天然的A链相连接,这种半合成的牛胰 岛素在1964年获得成功。 第三步,把经过半合成考验的人工合成的A链与B链相连接。 终于在1965年9月17日完成了结晶牛胰岛素的全合成。通 过小鼠惊厥实验,证明一r纯化结晶的人工合成胰岛素确实 具有和天然胰岛素相同的活性。
(2)转肽 转肽 (transpeptidation)包 括转位反应和肽键的形成,这个过程 是和延伸因子从核糖体上解离下来同 时进行的。
(3)移位 移位 (translocation)指肽酰-tRNA和 mRNA相对于核糖体的移动,这一过程需要移位因子 EF-G,并需要TP水解功能。
(三)原核生物肽链合成的终止
▲ 真 核 生 物 肽 链 合 成 的 起 始
▲真核生物肽链的延伸 真核生物的在延伸阶段的反应也是不断地经历进 位、转肽和移位的循环,只是由eEF-1代替了原核系 统中的EF-Tu和EF-Ts,eEF-2代替了EF-G,转肽酶 的活性可能由核糖体蛋白提供。真菌还需要第三种 延伸因子eEF-3,该延伸因子的作用是维持翻译的忠 实性。
蛋白质前体的加工
1、N端甲酰甲硫氨酸或甲硫氨酸的切除 2、二硫键的形成与重排
3、肽链的剪切
4、氨基酸侧链的修饰
5、肽链的折叠
6、亚基的聚合
An overview of protein synthesis.
牛胰岛素的合成过程
第一步,先把天然胰岛素拆成两条链,再把它们重新合成 为胰岛素。这由“拆合组”负责,任务是将天然胰岛素的 A链和B链上的3个二硫键打开,从而把胰岛素拆分成两条 链,随后再把它们重新合成。如果重合成的胰岛素具有生 物活性,那就为人工全合成胰岛素指明了一条路径。维格 纳奥德曾多次做过这个拆合实验,但没有一次成功。经过 不懈的努力,我国科学家小组于1959年突破了这一难题, 重新合成出LJ原来活力相同、形状一样的胰岛素,并成功 使胰岛素产率达到了5%一10%,到1963年已经提高到50%。
二、氨酰-tRNA的合成
蛋白质合成中的质量控制
为了防止潜在的蛋白质错误合成,某些氨基酰-tRNA合 成酶具有编校功能去除错误活化的氨基酸或者错误接载的 氨基酰化tRNA,通过控制蛋白质的生物合成原料的质量, 对蛋白质合成进行质量控制。该实验室已经证明亮氨酰tRNA合成酶(LeuRS)能够通过不依赖-或依赖tRNA转移前 编校途径水解误活化的非对应氨基酸,即使误活化的氨基 酸转移到tRNA分子上也可以通过转移后编校途径将其水 解。 LeuRS通过多条编校途径在反应的各个步骤清除错 误的反应产物和中间物。
小组成员:张夙夙 王振楠
潘维扬 韩盟 严聪
翻译 模板
蛋白质的生物合成 mRNA
氨基酸的运载体
蛋白质合成场所 各种辅因子
tRNA
核糖体 起始因子 延伸因子 释放因子 核糖体循环因子
一、遗传密码
1、遗传密码是三联体
简并性
2、遗传密码的特点
通用性 变偶性
表9-4 密码子和反密码子配对的变偶性
I U G tRNA反密码子的 第一位碱基 mRNA密码子的第 U,C, A,G U,C A 三位碱基 A U C G
▲真核生物肽链合成的终止 真核生物的肽链终止也需要蛋白因子,只需 一种终止因子eRF,此终止因子可识别3种终止 密码子,并需要三磷酸鸟苷。在人、爪蟾、酵 母等中,eRF有非常类似的肽链结构。
雷帕霉素可提高蛋白质 合成品质
雷帕霉素(rapamycin)可延长不同的生物寿命的发现, 掀起了研究其潜在的分子机制的研究热潮。雷帕霉素复合物 1(mTORC1)的哺乳动物靶点可调节细胞生长,并可能通过 控制mRNA的翻译调控生物体的衰老。然而,抑制 mTORC1和降低蛋白质的合成为何可以延长寿命仍然是一 个悬而未决的问题。
三、蛋白质生物合成过程
(一)原核生物肽链合成的起始
(1) 形成mRNA-30S复合物: IF3-30S-mRNA复合物 (2) 30S起始复合物与fMe-tRNAfMet结合: 与fMet-tRNAfMet-GTP-IF-2结合 (3) 形成70S起始复合物
(二)原核生物肽链的延伸 1)进位 进位 (entrance)指 氨酰-tRNA的反密码子与 mRNA的密码子在核糖体识别, 使氨酰-tRNA进入核糖体的A 位。
研究发现DNA密码子包含 蛋白质合成速度信息
“遗传密码一直被认为存在富余现象,但冗余密码子 显然并非完全一样,”加州大学霍华德休斯医学研究所 的Jonathan Weissman博士说。 许多的生物会明显偏好编码同一个氨基酸的数个密 码子中的其中一个,即便是在不管选用哪个密码子其结构 都相同的情况下。这就出现一个无法回避的问题:既然富 余的密码子作用都是一样的,为什么自然选择会对其中一 个有偏好。新的研究完美地回答了这个问题。