第五章 细胞表面及其特化_PPT幻灯片
合集下载
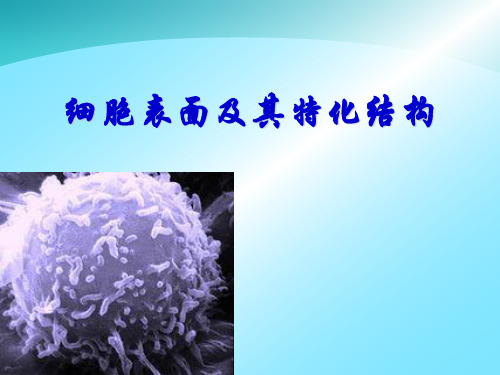
细胞表面及其特化结构
1. Macara, I. G. et al. 2006. Closing the GAP between polarity and vesicle transport. Cell 125: 419419-421. 2. Maeda S, et al. 2009. Structure of the connexin 26 gap junction channel at 3.5 A resolution. Nature 458(7238):597-602. 458(7238):5973. Kawasaki A, et al. 2009. Modulation of connexin 43 in rotenone-induced model of Parkinson's rotenonedisease. Neuroscience 160(1):61-68. 160(1):614. Freidin M, et al. 2009. Connexin 32 increases the proliferative response of Schwann cells to neuregulinneuregulin-1 (Nrg1). Proc Natl Acad Sci U S A. 106(9):3567-72 106(9):35675. Jiang, J. X. & Gu, S. 2005. Gap junction- and hemichannel-independent actions of connexins. junctionhemichannelBiochim. Biophys. Acta 1711: 208-214. 208-
细菌表面的鞭毛 人气管上皮细胞表面的纤毛
细胞表面的一种特化结构---细胞连接 细胞表面的一种特化结构---细胞连接 ---
细菌表面的鞭毛 人气管上皮细胞表面的纤毛
细胞表面的一种特化结构---细胞连接 细胞表面的一种特化结构---细胞连接 ---
细胞膜和细胞表面PPT课件
脂双层结构的形成
the polar head groups in all phospholipids can pack together into the characteristic bilayer structure. Sphingomyelins are similar in shape to phosphoglycerides and can form mixed bilayers with them. Cholesterol and other steroids are too hydrophobic to form a bilayer structure unless they are mixed with phospholipids.
膜脂——生物膜的基本组成成分
Three Classes of Lipids Are Found in Biomembranes
膜脂可分为三大类,磷脂(磷脂甘油), 鞘脂和胆固醇
A typical biomembrane is assembled from phosphoglycerides,sphingolipids, and steroids. All three classes of lipids are amphipathic molecules having a polar (hydrophilic) head group and hydrophobic tail.
in plasma membranes from human erythrocytes, almost all the sphingomyelin and phosphatidylcholine, both of which form less fluid bilayers, are found in the exoplasmic leaflet. In contrast, phosphatidylethanolamine, phosphatidylserine, and phosphatidylinositol, which form more fluid bilayers, are preferentially located in the cytosolic leaflet.
上皮组织细胞表面特化结构及功能PPT文档共25页

60、人民的幸福是至高无个的法。— —西塞 罗
谢谢!
51、 天 下 之 事 常成 于困约 ,而败 于奢靡 。——陆 游 52、 生 命 不 等 于是呼 吸,生 命是活 动。——卢 梭
53、 伟 大 的 事 业,需 要决心 ,能力 ,组织 和责任 感。 ——易 卜 生 54、 唯 书 籍 不 朽。——乔 特
55、 为 中 华 之 崛起而 读书。 ——周 恩来Biblioteka 上皮组织细胞表面特化结构及功能
56、极端的法规,就是极端的不公。 ——西 塞罗 57、法律一旦成为人们的需要,人们 就不再 配享受 自由了 。—— 毕达哥 拉斯 58、法律规定的惩罚不是为了私人的 利益, 而是为 了公共 的利益 ;一部 分靠有 害的强 制,一 部分靠 榜样的 效力。 ——格 老秀斯 59、假如没有法律他们会更快乐的话 ,那么 法律作 为一件 无用之 物自己 就会消 灭。— —洛克
谢谢!
51、 天 下 之 事 常成 于困约 ,而败 于奢靡 。——陆 游 52、 生 命 不 等 于是呼 吸,生 命是活 动。——卢 梭
53、 伟 大 的 事 业,需 要决心 ,能力 ,组织 和责任 感。 ——易 卜 生 54、 唯 书 籍 不 朽。——乔 特
55、 为 中 华 之 崛起而 读书。 ——周 恩来Biblioteka 上皮组织细胞表面特化结构及功能
56、极端的法规,就是极端的不公。 ——西 塞罗 57、法律一旦成为人们的需要,人们 就不再 配享受 自由了 。—— 毕达哥 拉斯 58、法律规定的惩罚不是为了私人的 利益, 而是为 了公共 的利益 ;一部 分靠有 害的强 制,一 部分靠 榜样的 效力。 ——格 老秀斯 59、假如没有法律他们会更快乐的话 ,那么 法律作 为一件 无用之 物自己 就会消 灭。— —洛克
最新基础医学细胞生物学PPT课件-细胞膜及其表面-细胞表面及其特化结构教学讲义PPT课件

9
Medical Cell Biology
二、细胞表面的特化结构
细胞表面最明显的特化结 构有:微绒毛、内褶、纤毛和 鞭毛,常还有一些暂时的结构, 如变形足、皱褶等。
微绒毛
皱褶
圆形变形足
糖萼 通道与小泡
尖形变形足
内褶
10
(一)微绒毛
Medical Cell Biology
微绒毛(microvillus)广泛存 在于动物细胞的游离表面。细 长指状突起,直径100nm(内有 微丝),长约100-200nm。扩大 细胞作用的表面积,有利于细 胞的吸收。每个小肠上皮细胞 有1000-3000根微绒毛。
锚 与中间纤维的锚定连接
定
连 接
与肌动蛋白纤维的锚定 连接
桥粒与半桥粒
desmosome and hemi-
粘合带和粘合斑 adhesion belt and focal adhesion
22
Medical Cell Biology
1. 粘合带与粘着斑
粘合带位于紧密连接下方
粘合带
(adhesion belt)
ECM的功能:支持、保护和提供营养外,还控制 细胞的迁移、增殖、分化、细胞识别、粘着、通讯联 络、信号传导、形态建成、代谢、基因的表达与调控 等生理过程和炎症、损伤与修复、免疫应答以及肿瘤 转移等病理过程中具有重要的功能和作用。
42
Medical Cell Biology
四、细胞外基质
43
Medical Cell Biology
呈带状环绕细胞, 一般位于上皮细 胞顶侧面的紧密 连接下方。在粘 合带处相邻细胞 的间隙约15~ 20nm。
23
Medical Cell Biology
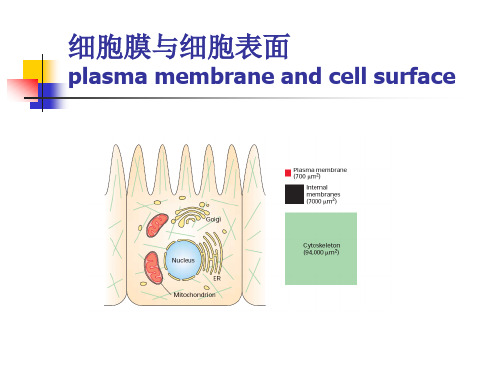
细胞膜与细胞表面(共67张PPT)

二、锚定连接(anchoring junctions):通过细胞骨架系统将细 胞与相邻细胞或将细胞与基质之间连接起来。
与中间纤维相关的锚定连接:桥粒(desmosome)、半桥粒 (hemidesmosome)
与肌动蛋白纤维相关的锚定连接:粘着带(adhesion)、粘着 斑(focal adhesion)
第一节 细胞膜与细胞表面特化结构
膜均衡技术(film balance)
由Langmuir 于1917年设计,至今仍是研究人工膜的重要方法。
膜的结构模型:
1. 双分子片层模型(也称Danielli-Davson 模型)
2. 单位模型(unit membrane model) 3. 流动镶嵌模型 (fluid mosaic model )
究中有广阔应用前景。
三、膜蛋白
(一)类型
膜周边蛋白(peripheral proteins)或外在膜蛋白
(extrinsic protein) 膜内在蛋白(integral proteins)
(二)膜内在蛋白与膜脂结合的方式 (三)去垢剂(detergent)
(四)、膜的流动性
(一)膜脂的流动性
膜的流动性由以下几个因素决定: 脂分子中脂肪酸链的长短、饱和程度、温度。 胆固醇分子 对膜的流动性也起着重要的调节作用。
分子,含糖量可达90%-95%。
软骨中的蛋白聚糖是已知的最大巨分子之一。单个分子 长达4微米,体积比细菌大。
功能:保护作用(抗变形能力等);蛋白聚糖还可与多种生
长因子结合,是细胞外的激素富集与储存库,利于调控分子 与细胞表面受体结合,有效完成信号的传导。 三、层粘连蛋白和纤连蛋白
1.层粘连蛋白(laminin)
(二)膜蛋白的流动性
与中间纤维相关的锚定连接:桥粒(desmosome)、半桥粒 (hemidesmosome)
与肌动蛋白纤维相关的锚定连接:粘着带(adhesion)、粘着 斑(focal adhesion)
第一节 细胞膜与细胞表面特化结构
膜均衡技术(film balance)
由Langmuir 于1917年设计,至今仍是研究人工膜的重要方法。
膜的结构模型:
1. 双分子片层模型(也称Danielli-Davson 模型)
2. 单位模型(unit membrane model) 3. 流动镶嵌模型 (fluid mosaic model )
究中有广阔应用前景。
三、膜蛋白
(一)类型
膜周边蛋白(peripheral proteins)或外在膜蛋白
(extrinsic protein) 膜内在蛋白(integral proteins)
(二)膜内在蛋白与膜脂结合的方式 (三)去垢剂(detergent)
(四)、膜的流动性
(一)膜脂的流动性
膜的流动性由以下几个因素决定: 脂分子中脂肪酸链的长短、饱和程度、温度。 胆固醇分子 对膜的流动性也起着重要的调节作用。
分子,含糖量可达90%-95%。
软骨中的蛋白聚糖是已知的最大巨分子之一。单个分子 长达4微米,体积比细菌大。
功能:保护作用(抗变形能力等);蛋白聚糖还可与多种生
长因子结合,是细胞外的激素富集与储存库,利于调控分子 与细胞表面受体结合,有效完成信号的传导。 三、层粘连蛋白和纤连蛋白
1.层粘连蛋白(laminin)
(二)膜蛋白的流动性
细胞膜及其表面细胞生物学优秀课件

(一)膜的不对称性
1、膜蛋白分布的不对称性 1) 膜蛋白在脂双分子层中、膜内外的位置分布是不对称的 2) 膜蛋白在膜内的排布方向是不对称的
2、膜脂的不对称性 1) 磷脂:磷脂酰胆碱和鞘磷脂多分布在膜外层
磷脂酰乙醇胺和磷脂酰丝氨酸多分布在膜内层 2) 胆固醇:主要分布在膜外层。 3) 糖脂 :全部分布在膜外层。
3、脂锚定蛋白 (lipid-anchored protein)
又称脂连接蛋白,这类膜蛋白定位于膜的两侧,类似于 膜周边蛋白,但与其不同的是以共价键与脂双层分子结 合,因此,分离锚定蛋白须用去垢剂或有机溶剂处理。
一、膜的化学组成
(三) 膜糖
共价键
单糖或多聚糖 + 膜 脂
共价键
单糖或多聚糖 + 膜蛋白
一、膜的化学组成
(二) 膜蛋白
1、膜内在蛋白 (integral protein)
贯穿脂双层,两端露出膜内外——跨膜蛋白
非胞质面 脂 双 分 子 层
胞质面
1 单次穿膜: 单条a-螺旋贯穿脂质双层
2 多次穿膜: 数条a-螺旋折返穿越脂质双层
内在膜蛋白具有双亲性,其亲水区域暴露在膜的内外表面与水相吸, 它们的疏水区域嵌入膜内,与脂类分子疏水尾部通过疏水键结合, 不易分离提纯。
二、膜的分子结构
★★(三)液态镶嵌模型(fluid mosaic model)
1972年,Singer 和 Nicolson 总结提出,主要论点:
1. 流动的脂双分子层构成生物膜的连续主体。 2.球形的膜蛋白以各种形式镶嵌在脂双分子层中或附着在膜表面。 3.强调了膜的流动性和不对称性。
评价: 液态镶嵌模型可以解释
(一) 膜脂
生物膜上的脂类统称膜脂。
1、膜蛋白分布的不对称性 1) 膜蛋白在脂双分子层中、膜内外的位置分布是不对称的 2) 膜蛋白在膜内的排布方向是不对称的
2、膜脂的不对称性 1) 磷脂:磷脂酰胆碱和鞘磷脂多分布在膜外层
磷脂酰乙醇胺和磷脂酰丝氨酸多分布在膜内层 2) 胆固醇:主要分布在膜外层。 3) 糖脂 :全部分布在膜外层。
3、脂锚定蛋白 (lipid-anchored protein)
又称脂连接蛋白,这类膜蛋白定位于膜的两侧,类似于 膜周边蛋白,但与其不同的是以共价键与脂双层分子结 合,因此,分离锚定蛋白须用去垢剂或有机溶剂处理。
一、膜的化学组成
(三) 膜糖
共价键
单糖或多聚糖 + 膜 脂
共价键
单糖或多聚糖 + 膜蛋白
一、膜的化学组成
(二) 膜蛋白
1、膜内在蛋白 (integral protein)
贯穿脂双层,两端露出膜内外——跨膜蛋白
非胞质面 脂 双 分 子 层
胞质面
1 单次穿膜: 单条a-螺旋贯穿脂质双层
2 多次穿膜: 数条a-螺旋折返穿越脂质双层
内在膜蛋白具有双亲性,其亲水区域暴露在膜的内外表面与水相吸, 它们的疏水区域嵌入膜内,与脂类分子疏水尾部通过疏水键结合, 不易分离提纯。
二、膜的分子结构
★★(三)液态镶嵌模型(fluid mosaic model)
1972年,Singer 和 Nicolson 总结提出,主要论点:
1. 流动的脂双分子层构成生物膜的连续主体。 2.球形的膜蛋白以各种形式镶嵌在脂双分子层中或附着在膜表面。 3.强调了膜的流动性和不对称性。
评价: 液态镶嵌模型可以解释
(一) 膜脂
生物膜上的脂类统称膜脂。
细胞膜及其表面结构PPT课件

三)膜糖类
➢与膜脂或膜蛋白结合成糖脂或糖蛋白,仅分布于 膜的外表面。
➢主要为一些低聚糖组成,如半乳糖、甘露糖、岩 藻糖、半乳糖胺、葡萄糖、葡萄糖胺、唾液酸等 七种。
第19页/共59页
细 胞 被
膜
蛋
脂
白
双
层
细胞质
第20页/共59页
血型糖蛋白
第21页/共59页
二、细胞膜的分子结构
第22页/共59页
主要形式为紧密连接(tight junction) ; 分子机制:相邻细胞的特殊膜蛋白相互交联
(嵴线)封闭了细胞之间的空隙。
特点:细胞膜之间连接紧密,无空隙; 分布:广泛存在于各种上皮细胞之间。
第41页/共59页
细胞膜 细胞间隙 (嵴线)
第42页/共59页
紧密连接的功能
1、形成渗漏屏障,起重要的封闭作用; 例:血脑屏障和血胎屏障
3)膜流动性的生理意义
➢ 质膜的流动性是保证其正常功能的必要条件。 ➢ 当膜的流动性低于一定的阈值时,许多酶的活动和跨膜运输将停止; ➢ 反之如果流动性过高,又会造成膜的溶解。
第29页/共59页
2、质膜的不对称性
➢膜脂分布的不对称性:脂类双分子层内外两层
脂质分子种类、含量和比例均不相同;
➢膜蛋白分布的不对称性:没有任何一种蛋白是
总之:锚定连接将相邻 细胞的骨架系统或将细 胞与基质相连形成一个 坚强、有序的细胞群体。
第53页/共59页
三、通讯连接(communicating junction)
• 除了连接作用以外,还具有介导细 胞间通讯的作用; • 是动物细胞(包括人)间最主要的 一种连接方式; • 包括:间隙连接和突触连接。
旧称脑磷脂; ④ 磷脂酰肌醇phosphatidylinositol,PI。
上皮组织—细胞表面的特化结构(组织学与胚胎学课件)

细胞表面的特化结构
上皮细胞有极性,在各表面 形成了与功能相适应的结构。 这些结构也可见于其他组织 的细胞。
细胞表面的特化结构
1、游离面 微绒毛ቤተ መጻሕፍቲ ባይዱ纤毛
2、侧面 — 细胞连接
基膜 3、基底面 质膜內褶
半桥粒
紧密连接 中间连接 桥粒 缝管连接
微绒毛(LM) 纹状缘、刷状缘
微绒毛 (EM)
纤毛(LM)
纤毛的摆动
纤 毛(EM)
细胞连接
紧密连接
连
中间连接
接 复
合
桥粒
体
缝管连接
细胞连接
左:模式图 右:电镜像
Mv 微绒毛 TJ 紧密连接 IJ 中间连接 De 桥粒 GJ 缝隙连接
连接复合体
缝隙连接示意图
基膜 光镜结构图
PAS染
银染 HE染
质膜内褶超微结构模式图
基膜
质膜内褶与基膜电镜像 (肾小管)
上皮细胞有极性,在各表面 形成了与功能相适应的结构。 这些结构也可见于其他组织 的细胞。
细胞表面的特化结构
1、游离面 微绒毛ቤተ መጻሕፍቲ ባይዱ纤毛
2、侧面 — 细胞连接
基膜 3、基底面 质膜內褶
半桥粒
紧密连接 中间连接 桥粒 缝管连接
微绒毛(LM) 纹状缘、刷状缘
微绒毛 (EM)
纤毛(LM)
纤毛的摆动
纤 毛(EM)
细胞连接
紧密连接
连
中间连接
接 复
合
桥粒
体
缝管连接
细胞连接
左:模式图 右:电镜像
Mv 微绒毛 TJ 紧密连接 IJ 中间连接 De 桥粒 GJ 缝隙连接
连接复合体
缝隙连接示意图
基膜 光镜结构图
PAS染
银染 HE染
质膜内褶超微结构模式图
基膜
质膜内褶与基膜电镜像 (肾小管)
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
紧密连接(tight junction) 黏合连接(anchoring junctions) 通讯连接(communicating junctions)
细胞连接
紧密连接
存在部位:体内管腔及腺体上皮细胞靠腔面的一端, 结构特点:相邻细胞紧密相贴,无间隙。 功 能: 1.机械封闭作用; 2.保证物质转运的方向性; 3.形成屏障,限制胞膜顶部与侧部和基底部之间的蛋白 质扩散。
点状桥粒位于粘合带下方
点状桥粒的结构模型
半桥粒:细胞膜上的膜蛋白整合素将上皮细胞固着在基底 膜上, 中间丝终止于半桥粒的致密斑内。
半桥粒的结构图
通讯连接
分布广泛,包括间隙连接和化学突触。 间隙连接:多见于上皮细胞深部侧表面。连接处相邻细胞
膜中含有连接子。 功能: 1、介导细胞之间的连接;
2、偶联细胞间通讯,包括电偶联和化学偶联,即 通过细胞间的离子和分子的传递进行细胞通讯。
直径约为0.1μm。内芯由肌动蛋白丝束组成。 作用:扩大了细胞的表面积,有利于细胞同外
环境的物质交换。如小肠上微绒毛,使细胞表 面积扩大了30倍
微绒毛
纤毛和鞭毛
纤毛(cilia)和鞭毛(flagella)是细胞表 面伸出的条状运动装置。二者在发生和结构 上并没有什么差别。 纤毛和鞭毛都来源于中心粒。详细结构和 功能可参见细胞骨架一章。
第一节 细胞外被与胞质溶胶
细胞外被的概念 细胞外被的功能 胞质溶胶的概念
细胞外被
细胞外被概念
细胞膜外表面覆盖的一层粘多糖物质。用重金属染料, 如钌红染色后,在电镜下可显示厚约10~20nm的结构,边界 不甚明确。
膀胱上皮细胞表面的糖 被,钌红染色的电镜超 薄切片
第二节 细胞间的连接
紧密连接
黏合连接
存在部位:在上皮组织,心肌和子宫颈等组织中含量丰富。 分类:带状桥粒、点状桥粒和半桥粒。 结构特点:将细胞机械的黏着方,相邻细胞间形成一 个连续的带状结构。能增强组织的机械性能。
点状桥粒:铆接相邻的细胞,提供细胞内中间丝的黏合位点,起 支持和抵抗外界压力的作用,连接处细胞膜内侧胞质面有对称而 平行的2个圆盘形斑,称为胞质斑。
纤毛和鞭毛
连接子是由6个穿膜的膜内在蛋白分子围成,中 央有2nm的隧道。构成了细胞间的直接通道。连接
子大小为6-8nm的颗粒,呈六角形。
间隙连接
第三节 细胞表面的特化结构
细胞膜常带有许多特化的附属结构。如: 微绒毛、纤毛、鞭毛等等,这些特化结构在 细胞执行特定功能方面具有重要作用。
微绒毛
细胞表面伸出的细长突起,广泛存在于动物细 胞表面。
细胞连接
紧密连接
存在部位:体内管腔及腺体上皮细胞靠腔面的一端, 结构特点:相邻细胞紧密相贴,无间隙。 功 能: 1.机械封闭作用; 2.保证物质转运的方向性; 3.形成屏障,限制胞膜顶部与侧部和基底部之间的蛋白 质扩散。
点状桥粒位于粘合带下方
点状桥粒的结构模型
半桥粒:细胞膜上的膜蛋白整合素将上皮细胞固着在基底 膜上, 中间丝终止于半桥粒的致密斑内。
半桥粒的结构图
通讯连接
分布广泛,包括间隙连接和化学突触。 间隙连接:多见于上皮细胞深部侧表面。连接处相邻细胞
膜中含有连接子。 功能: 1、介导细胞之间的连接;
2、偶联细胞间通讯,包括电偶联和化学偶联,即 通过细胞间的离子和分子的传递进行细胞通讯。
直径约为0.1μm。内芯由肌动蛋白丝束组成。 作用:扩大了细胞的表面积,有利于细胞同外
环境的物质交换。如小肠上微绒毛,使细胞表 面积扩大了30倍
微绒毛
纤毛和鞭毛
纤毛(cilia)和鞭毛(flagella)是细胞表 面伸出的条状运动装置。二者在发生和结构 上并没有什么差别。 纤毛和鞭毛都来源于中心粒。详细结构和 功能可参见细胞骨架一章。
第一节 细胞外被与胞质溶胶
细胞外被的概念 细胞外被的功能 胞质溶胶的概念
细胞外被
细胞外被概念
细胞膜外表面覆盖的一层粘多糖物质。用重金属染料, 如钌红染色后,在电镜下可显示厚约10~20nm的结构,边界 不甚明确。
膀胱上皮细胞表面的糖 被,钌红染色的电镜超 薄切片
第二节 细胞间的连接
紧密连接
黏合连接
存在部位:在上皮组织,心肌和子宫颈等组织中含量丰富。 分类:带状桥粒、点状桥粒和半桥粒。 结构特点:将细胞机械的黏着方,相邻细胞间形成一 个连续的带状结构。能增强组织的机械性能。
点状桥粒:铆接相邻的细胞,提供细胞内中间丝的黏合位点,起 支持和抵抗外界压力的作用,连接处细胞膜内侧胞质面有对称而 平行的2个圆盘形斑,称为胞质斑。
纤毛和鞭毛
连接子是由6个穿膜的膜内在蛋白分子围成,中 央有2nm的隧道。构成了细胞间的直接通道。连接
子大小为6-8nm的颗粒,呈六角形。
间隙连接
第三节 细胞表面的特化结构
细胞膜常带有许多特化的附属结构。如: 微绒毛、纤毛、鞭毛等等,这些特化结构在 细胞执行特定功能方面具有重要作用。
微绒毛
细胞表面伸出的细长突起,广泛存在于动物细 胞表面。