稻瘟病抗病育种研究进展文献综述
水稻抗稻瘟病育种方法概述

l 杂 交 育 种
杂交育种技术是抗 稻瘟病育种常规的手段 , 也仍然 是 目前抗病育种主要 的技术之一 。一般单基 因抗
性可 通 过 回交 的方式 育成 抗 病 品种 。将具 有 1个或 2个显 性抗 病 基 因的非 轮 回亲 本与 各 种农 艺 性状 好 的
轮回亲本杂交, 再将轮 回亲本与子代杂交 4 次 , ~5 这样既可 以导人抗病基 因又保持了轮 回亲本的优 良性 状 。但 是 , 多基 因抗性 就 很难 通 过简 单 的 回交育成 多 抗或 广谱 抗 性 的品 种 , 时应 采用 复 交 的方 法 才 能将 这 抗病基 因全部传给后代。先将一个或多个授体亲本的抗病基因引入一个共 同的品种, 然后从每一个抗病
维普资讯
2 ( ) 3 ~3 2 2 :1 3
Vo122. . No. 2
广
西
农
学
报
20 0 6年 4月
Ap i 2 0 r,06 l
J u  ̄ o a w i rc l r om fGu n x Ag iut e u
水 稻 抗 稻 瘟 病 育 种 方 法 概 述
whc cu e rsb e d n , ua in b e dn ,elb ed n dmoe ua re i . i i ld dco s re ig m tt r e i c l re ig a l lrb e dn h n o g n c g
Ke l s Rie Re i a c oRieB a t Bre ig ywo d c ssn et c l t s edn
的后 代 选择 抗病 的株 系 。其 次 , 水稻 抗瘟 性 遗传力 因品种 不 同而 有所 差异 , 因此在 进行 抗 病 育 种时 要 注 意
浅析四川稻瘟病发生情况及防治措施

13农业灾害研究2020,10(9):13-15浅析四川稻瘟病发生情况及防治措施刘佳鑫四川省乐山市井研县融媒体中心,四川乐山 613100摘要 水稻栽种过程中容易发生稻瘟病、纹枯病和稻黑穗等病害稻瘟病是世界上最危害水稻的病虫害疾病之一,被称作是水稻的“癌症”。
世界各地的水稻种植区都可能发生稻瘟病,不受年份、区域和季节的限制,且发病率逐年增加。
如不及时控制,将会导致水稻产量大幅减少,甚至绝产,颗粒无收。
对四川稻瘟病发生趋势进行初步剖析并提出有效的综合防治办法。
关键词 稻瘟病;发生情况;防治措施中图分类号: S435.111.41 文献标识码:B 文章编号:2095–3305(2020)09–013–03DOI:10.19383/ki.nyzhyj.2020.09.007西南地区种植稻谷的面积在46万 hm 2左右[1]。
四川发布的四川水稻栽种面积排名全国第7,水稻总产量排名全国主要稻谷生产省份第6。
同时,四川稻谷种植农业合作社名录显示,四川稻谷种植合作社从2016年的1 089家到2018年的2 617家,3年间增加了1 528家,增长率高达140.31%(图1)。
图1 四川省稻谷种植合作社发展趋势图3000250020001500100050002018年,10个超级水稻品种中,四川占比30%,标志着四川水稻从产量型向优质高产型跨越。
四川省农业厅发布预测消息,2016年—2018年稻瘟病平均发生率为中等发生,局部偏重,预测发生病害秧田面积在20万hm 2左右,特别是2018年,全省19个市、州均发生稻瘟病,局部地区发生穗颈病尤为突出,比2017年同时期增加近500%,是自2008年以来第4个高病发年。
发生的原因在于主要病害菌源充足、主栽品种抗瘟性不高、气象条件总体有利于病害发生。
针对稻瘟病的发生,四川地区主要采用农业、化学、生物防治三管齐下的防治办法,在水稻种子选择、田间栽培管理和疾病控制方面取得良好的效果。
中抗稻瘟病常规早稻新品种冈早籼11号的选育

中抗稻瘟病常规早稻新品种冈早籼11号的选育1. 引言1.1 选育背景稻瘟病是稻谷重要病害之一,对稻谷产量造成了严重影响。
为了解决这一问题,科研人员进行了大量工作,通过选育抗稻瘟病的新品种来提高稻谷的产量和品质。
中抗稻瘟病早稻新品种冈早籼11号的选育背景可以追溯到多年前,当时稻瘟病对稻谷产量造成了严重影响,迫切需要一种高产、抗病性强的新品种来替代传统品种。
由于稻瘟病病原真菌具有较强的适应能力,在选育过程中遇到了诸多困难。
选育背景中包括对稻瘟病病原菌的研究,种质资源的筛选,以及对抗病性状的遗传分析等内容。
选育背景的详细了解对于理解冈早籼11号的选育目的和方法具有重要意义,可以为后续的研究提供参考和指导。
1.2 选育目的选育目的是为了培育出具备优良品质和高抗性的早稻新品种,以应对稻瘟病等病虫害的严重威胁,提高稻谷产量和品质,满足人民对粮食安全和生活质量的需求。
通过选育,希望能够获得一种中抗稻瘟病的早稻新品种,具有较高的产量潜力、稳定的表现、优良的品质和高度的抗性,在不同气候条件和土壤类型下均能展现出较好的适应性和生长表现。
也希望通过选育工作的开展,进一步挖掘和利用优良的遗传资源,提高我国早稻的品质水平和病虫害抗性,不断推动水稻产业的发展,为我国的粮食安全和农业持续发展作出贡献。
2. 正文2.1 选育方法选育方法是培育新品种的关键步骤之一,决定了新品种的遗传特性和表现。
中抗稻瘟病常规早稻新品种冈早籼11号的选育方法主要包括以下几个步骤:1. 亲本选择:选育新品种首先需要选择适合的亲本,亲本的选择将直接影响到后代的遗传特性。
在冈早籼11号的选育过程中,选取了具有较强抗病性和适应性的优良亲本进行杂交。
2. 交配组合:根据目标性状的选择,进行适当的交配组合,利用亲本间的优势互补效应,提高后代的遗传优势。
通过合理的交配组合,可以加速新品种的选育进程。
3. 杂交选种:在选择出有潜力的优良杂交组合后,进行大量的杂交选种。
几丁质酶-葡聚糖酶双价基因导入滇型杂交稻恢复系提高稻瘟病抗性.

将水稻几丁质酶基因 +51’# 或 +51’; 转入粳稻,转基
因植株对两个稻瘟病生理小种表现高抗,而且这种 抗性还能万稳方定数遗据传给下一代[+]。@A:(B )""")将水稻
!3’ 水稻基因枪转化 质粒 2M8ND()CO+ PL)包裹金粉,用基因枪(/5E(
图 ! 植物表达载体质粒 "#$%& 构成示意图 ’()* ! +,- ./-01, 23" 45 "63.2(7 "#$%&
)3)3* 转基因品系 用于 稻 瘟 病 抗 性 鉴 定 的 C 个 品 系 来 源 于 经
EFG7=AH: 杂交分析阳性的“南 *"”转化植株所繁衍出 的后代,经 /D0 分析阳性的 ;# 代品系。 )3)3$ 供试稻瘟病菌菌株
IJ,)(&#、KC"、青 "*(&C(L、K,,(*’, 等 # 个 稻 瘟 病菌株,它们分别属于 #$’、))’L、$$’、&$’7 # 个不同 生理小种。菌种来源于云南省农业科学院植物保护
对吉林省水稻区试品种(系)稻瘟病的抗性鉴定

Jl 6、 、 - 、 等 4 个吉林小种 。 7、 Jl J _J 4、 。 1 13 苗 期 分 菌 系 人 工 接种 鉴 定 .
供试材料播种于育苗盘 中, 常规管理 , 播种前 4d施用一次氮肥。 病菌接种是在幼苗长 至 3 4叶 ~ 时, l 以 0菌系分 生孢 子悬浮液 ( 浓度为 10 显微镜视野 2 3 0倍 0~ 0个孢子) 分别定量 隔离 喷雾接种 , 接 种后在 2 —3 5 7℃下 黑暗保湿 2 左右 , 0h 然后 在 2 3 0~ O℃及高湿条件 ( 喷水) 连续 下培育 。8 0d调 ~1
J f J . J7 、47 Ji、43 J5 、47J4 、41J 、43 J3 、37 J2 、 26 J5 、16 J4 、l4 J0 、l j 7 55 加1 47Jf 、 . Ji、46J4 、 45 J 、4o J2 、 37 J 、33 J 、26 J6 、17 J 、 l4j【 7 、 4 、 、 i 45 f . f . . 4 4 2 7 4 1
种( 进行 了抗 稻 瘟病 鉴 定 ,0 系) 19个 品 种完 成 了 3年 的连 续 鉴 定 , 中通 粳 6 2 通 院 5 通 育 0 U 0 、 其 1、 、 1 3 7 九
1B1 、 1B 、 20 F 7 吉 2 O F 6、 院 6号 、 育 O— 2 、 2 0 F 9、 系 18 通 丰 8号 、 研 7 0 4 九 0 3 吉 0O 2 、 0O 4 通 通 12 1 吉 0 0 5 通 5、 通 号 、 0 G 2 白 12 吉 O — 3 1 九 0 G 5、 2O G 8 九 0Y 1 九 0 Y 1 、 0 Y 5 通 院 7号 、 院 9 九 2 A 、 2 、 13 4 、 2 B 吉 0 O 1 、 2 C1 、 2 C 7 九 2 C 、 通
水稻区试品种抗稻瘟病鉴定结果分析
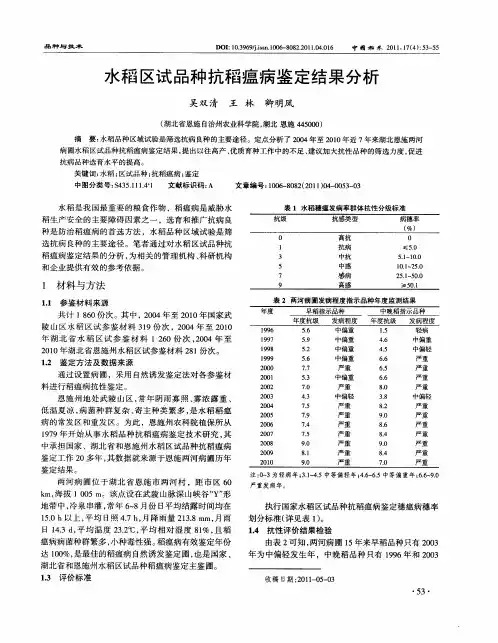
执行 国家水稻区试 品种抗 稻瘟 病鉴定穗瘟病穗率 划分标 准( 详见表 1。 )
水稻是 我 国最重要 的粮 食作物 ,稻瘟病是威 胁水
稻生产安全 的主要障碍 因素 之一 ,选育和推广抗 病 良 种是 防治稻瘟 病的首选方法 ,水 稻 品种 区域试验 是筛 选抗病 良种 的主要 途径 。笔者通 过对水稻 区试 品种抗
稻瘟病鉴定结果 的分析 , 为相关 的管理机构 、 科研机构 和企业提供有效 的参考依 据。
21 不同年度 间参鉴材料抗性结果分析 . 由表 3 可知 ,各年度 间参鉴材料抗性水平 分布趋 势基本一致 ,感病 材料在各级水 稻区试 品种 中占有极 显著优势 ,但年度 间由于环境 和参鉴 品种不 同存在 一 定差异 。
22 早、 晚稻间参 鉴材料 抗性结果分析 . 中、 从表 4可知 , 、 、 早 中 晚稻 间参 鉴材 料抗性 水平 分
1 抗性评价结果检验 . 4 由表 2 可知 , 两河病 圃 1 5年来早稻品种只有 2 0 03 年 为中偏轻发 生年 ,中晚稻 品种 只有 19 年 和 2 0 96 03
收稿 日期 :0 1 0 — 3 2 1- 5 0
・
瘟 病病菌种群繁多 , 小种毒性强 。 稻瘟病有效鉴定年份 达 10 是最佳 的稻瘟病 自然诱发鉴定 圃 , 0 %, 也是 国家 、
21 0 0年湖北省恩施州水稻 区试参鉴 材料 2 1 8 份次 。
分子标记在水稻抗稻瘟病育种中的应用
文 章 编 号 : 10 -75 20 )30 4 -3 D 1 0 (0 7 0 - 40 4 0
形式表现, 在生物体的任何组织 、 各个发育阶段均可检 测 到 , 受季 节 、 境 限制 , 存 在 表 达 与 否 等 问题 ; 不 环 不 () 2 数量极 多, 布整个基 因组 , 遍 可检测座位几 乎无
这种常规的育种方法不仅周期长, 易受环境影响 , 而且 抗病基因又与不良性状连锁, 加之稻瘟病菌生理小种 复杂易变, 使得稻瘟病 的抗病育种工作进展缓慢。2 0
世 纪 8 代 以来 , O年 随着 分子 生物学及 分 子克隆 技术 的 发展 和完善 , 诞生 了 D A分子标 记技 术 。该 技术 是建 N
限 ;3 多 态性 高 , () 自然 界 存 在许 多 等 位 变异 , 须人 无 为创 造 ;4 表现 为 中性 , () 不影 响 目标 性 状 的表 达 ;5 ()
由子囊 菌 Manp r e r e H br) ar无 性 g aot i a( eetB r[ hgs
世代为 P r u r re Coe Sc. 引起 的稻瘟 yi l i gia( ok ) ac ] c aa s 病是广泛发生在世界各稻 区的最重要水稻病害之一。 据估计 , 17 19 在 95~ 90年 的 1 6年间 由稻瘟病引起的 全球稻谷损失高达 15 亿 t .7 , 年平均 1 千万 t 左右…。
凝胶 电 泳一F P( ea r ggai tgle cohis R L D nt n rde e l t pri u i n e r s—
5 %, 0 局部 田块甚至颗粒无收 。 j 实践证明, 防治稻瘟病最有效、 最经济、 最根本 的 方法是选育和利用抗病品种。长期以来的抗稻瘟病育 种研 究主要 是利用筛 选 出 的抗 源对 已有 的品种进 行改 良或作为亲本进行杂交配组 , 中选育抗 病新品种 。 从
同源序列法克隆植物抗病基因研究进展及其在野生稻中的应用
1
植物抗病基因编码蛋白的结构特点
尽管抗病基因之间的序列同源性很低,但Leis—
过抗病基因识别病原物的入侵,从而诱导植物产生 过敏反应或者随后引起系统抗性,是植物抵御病原 物入侵的重要方式。因此,抗病基因的克隆对于植 物抗病性研究及进行植物基因工程改良具有重要
意义。
ter等【l】通过分析已克隆的植物抗病基因编码产物的 结构特征,发现虽然不同抗病基因互作的病原物种
as
been cloned.Related studies
showed that most of the genes have
kinase ere.The structural
leucine-fich
repeat,nucleotide binding site,serine/threonine
not
非rI’IR—NBS—LRR类抗病基因(cc—NBS— 这类基因编码的蛋白含有一个核心的NBS结
LRR,CNL)
构域,近N端为non—TIR结构域,近C端为LRR结 构域。同样,此类基因也是在胞质中起作用。如水稻
e删驴司被克隆后,利用同源基因克隆方法,在大麦
等其他农作物中也分离克隆到了相应基因【l蚴1;还
brane
从玉米中克隆到抗圆斑病菌的胁以基因以来,截至 2004年底,已有近50个抗病基因从单子叶和双子 叶中被克隆【11。5】,其中大部分基因来自双子叶植物 番茄和拟南芥。在粮食作物中已克隆到15个抗病基
因【垌。
植物抗病基因的克隆一般采用产物导向法
(product-orientated approaches)、鸟枪射击法(shot-
X订基因[sl、抗谷子锈病基因R吣J【饲。
1.3
类受体蛋白(receptor—like protein,RLP)抗病
稻瘟病防治研究及其抗性鉴定基地
稻瘟病防治研究及其抗性鉴定基地和服务五十年防治在危害水稻生产的各类病害中,以稻瘟病为首,该病害遍布世界80多个国家,一般造成减产10%~20%,严重时可达40%~50%,特别严重的田块甚至颗粒无收。
湖南是水稻犬省,控制稻瘟病危害对保证我省水稻优良品种的研究与推广有着重要意义。
我省稻瘟病防治工作起始于二十世纪50年代,1954年,湖南省农业试验总场植保系(省植保所前身)与原华中农科所植保系协作,在古丈县黑泥潭对稻瘟病进行了调查研究。
次年,总场植保系提出胡麻叶斑病、稻热病(即稻瘟病)的调查计划,并于1956年对稻热病单独列题进行研究。
此后,稻瘟病的防治研究一直是省植保所的重点课题,并随着研究的不断深入,其内容也不断增加,领域不断拓宽。
其中,20世纪50年代中期,侧重于药剂防治的研究,其中对汞制剂防治稻瘟病的有效浓度、施用剂型、施药适期以及雨天施药技术等进行了系统研究;20世纪60年代,主要开展了科学的水肥管理防治稻瘟病效果的研究,通过研究,探明了化肥和绿肥与稻瘟病发生流行的关系;1972年以来,与浙江省农科院植保所一道在全国测报与全国稻瘟病科研协作等会议上,率先提出以推广抗病丰产品种为中心的综合防治措施是防治稻瘟病的经济而有效的途径;此后至今,省植保所作为全国科研协作组抗病性研究单位,组织省内外有关协作单位,开展了抗病丰产品种及抗源鉴定筛选的研究,并进行了以抗病丰产品种为中心的综合防治试验示范。
1、持久抗瘟性与抗性育种在大田生产中,由于一些抗性品种的抗瘟性在3--0 a中逐渐消失,因此给稻瘟病的防治提出了新的挑战。
为解决这一国际难题,省植保所在国家自然科学基金的资助下,由彭绍裘研究员主持,于1990年开始了稻类持久抗瘟性基因的发掘与利用研究,首次全面系统地提出了以病区时空动态效应强度抗病信息流质量反应为核心的持久抗瘟性鉴定方法,揭示了持久抗瘟性应具有的4个特征,分析了稻类持久抗瘟性形成和衰变的原因。
抗稻瘟病优质早籼湘早143的选育及其应用
13 4 全生育期 17 4 ,比对照浙 73 25天, 0 .天 3短 . 适合在长江
流域双季稻区作早 、 中熟 早 稻 种 植 。
5 农 艺性 状 .
湘 早 13 期 分 蘖力 较 强 , 株 松 散适 中 , 长 清 秀 , 4前 植 生 在 孕穗 期 遇 上 连 续 阴 雨 或 光 照 不 足等 不 良天 气 后 , 穗 时 穗 顶 抽 或 穗 基 部 会 出 现 部 分 颖 花 退化 。后 期 植 株 较 为紧 凑 , 壮抗 矮 倒 。株 高 7 8 m, 叶较 短 、 、 立 。稃 尖 无 色 , 易 掉 5 5c 剑 厚 直 不 粒 。穗 长 1 m 左 右 , 效 穗一 般 为 30穗 / , 穗 总 粒 数 8 c 有 6 每
维普资讯
育 种 与 品 种
中国 米 20 # g 5 06 - 期
抗 稻 瘟 病 优 质 早 籼 湘 早 13的选 育 及 其应 用 4
王联芳 余 应弘 吴云天 王建 龙 肖放华 龚浩如 张秋标
摘 要 :围绕 改 良早籼 稻食 用 品质 、 强其稻 瘟 病抗性 的 目 增 标, 利用 已有 的优 质稻 中 间材料 杂交 , 采用 本地选 育 与病 圃筛选
湘 早 13明 显强 于对 照 ,是 当前 优 质 稻 中一 个 难 得 的抗 稻 瘟 4 病的品纪 9 年 代开始 , 0 针对早籼稻育种 中产量 、 米质
与抗 性 难 以协 调 的难 题 , 课 题 组 加 强 了优 质早 籼 的 稻瘟 病 本
13 4 。该 品种 于 20 0 4年通 过湖 南省 审定 ,0 5年 通过 国家 审定。 20
湘 早 13稻瘟 病 抗 性 强 。02年 及 20 4 20 03年湖 南 省 两 年 区 试平均 , 湘早 13 性 叶瘟 25 、 颈 瘟 1 , 稻 瘟 病 , 4抗 .级 穗 级 抗 是 同组 参 试 品 系 中 唯 一 抗 稻 瘟 病 的 品 系 。20 年 及 20 03 04年两 年 国 家 区试 平 均 , 早 13 瘟 病 抗 性 加 权 平均 13 , 瘟 湘 4稻 .级 穗
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
造后,借助载体或化学物导入受体细胞,抗病基因插入受体细胞之后能在受付细 胞中表达且稳定遗传。抗稻瘟病基因包括水稻自身的抗病基因和与防卫反应有关 的基因。目前来源于小稻的两个抗稻瘟病基因(P a-b和Pa-ta)已被克隆。我国利 用基因工程在抗稻瘟病育种上取得了一定的感绩。吉林省延边朝鲜族自治区农科 所与吉林农业大学合作,成功地将孤的总DA N导入水稻,育成抗病高产的水稻新 品种”通32”[21]等。中山大学生物工程研究牛心[22]将Rc24基因转入竹粕B中,对 竹转68进行抗性鉴定表明,竹转68对93-203a菌株表现出免疫或高抗,而竹籼B 对竹籼B表现为高感。 3.1.4 细月包育种 3.1.4.1 花药培养
Abstract Rice blast is one of the most important three rice diseases which af
fect the production of rice seriously, the research of the prevention of blast pla
抗性育种防治稻瘟病的研究进展文献综述
姓名:周云云
班级:植保 11-2
学号:2011638
摘要:稻瘟病是严重影响水稻生产三大水稻病害之一,对稻瘟病的防治的研究对
其防治具有重要的意义,本文主要从稻瘟病的遗传育种防治上的最新研究进展进
行全面综述,并对我国稻瘟病防治面临的问题和未来前景进行了讨论和展望。
关键词:水稻 稻瘟病 防治 遗传育种
The literature review on the progress of the prevention and treatment of
resistancห้องสมุดไป่ตู้ breeding on blast
Name : ZHOU Yun-yun Class: Plant Protection 11-2 Student ID: 20116384
d discuss the problems our country is facing and the future prospects we can
expect.
Key words Rice; Blast; Prevention and treatment; resistance breeding;
诱变育种是指用物理、化学因素诱导植物的遗传特性发生变异,再从变异群 体中选择符合人们某种要求的单株,进而培育成新的品种或种质的育种方法。它 是继选择育种和杂交育种之后发展起来的一项现代育种技术。诱发突变的物理因 素主要指某些射线,如γ射线、X射线、β射线和中子流等;化学诱变剂主要指某 些烷化剂、碱基类似物、抗生素等化学药物。这些物理、化学因素能引起植物的 分子结构发生变化,细胞内生物化学过程发生改变,从而导致细胞器的结构及其 组成发生深刻变化,包括染色体畸变和基因突变。我国水稻诱变,以y射线为主。 近些年发展起来的空间诱变育种技术又叫航天育种技术,即利用卫星将作物种子
载上太空,利用太空特殊的环境(空间宇宙射线、微重力、高真空、重离子、变 化磁场等因素)的作用下产生遗传变异来选育新品种的方法。变异来选育新品种 的方法。广东省农科院将桂朝2号经1.5kR的γ射线在合子期处理,育成高抗稻瘟 病早熟品种辐桂1号,用55个稻瘟病菌株鉴定,其中81.8%表现抗病,而桂朝2号 只有27. 1%[19]。在水稻航天诱变育种上尤为突出,已育成8个抗病高产优质的水 稻新品种(组合)并通过省级品种审定委员会审定这些品种有赣早粕47号、航育1 号、华航1号、宇航2号和II优航1号等。 3.1.3 分子育种
1. 稻瘟病概述
稻瘟病是由异宗配合子囊菌(Magnaporthe grisea (hebert) Barr)的无性世代 Pyicularia grisea (Cook) Sacc.产生的无性孢子浸染水稻引起的真菌性病害[1]。
是最严重的水稻病害之一,稻瘟病的危害面积和危害程度较大,严重阻碍水稻高
产稳产。全球每年由稻瘟病导致的直接经济损失约50亿美元[2]。稻瘟病在水稻秧
y an important role in the production of rice. In this paper,I will review the la
test progress on the prevention and treatment of resistance breeding on blast an
稻瘟病菌很容易发生变异,存在着不同的生理小种。我国在1976-1979年间 以珍龙13、东农363、关东51、合江18、丽江新团黑谷等鉴别品种,对来自28个 雀(市)、自治区的827个有效稻瘟病分离物进行测定纪果,可区分为7群43个生理 小种,建立了一套寄生—病菌互作清晰的鉴别系统[11]。日本山田提出9个单基因 鉴别品种。段永嘉等以日本9个单基因鉴别品种作对照,对云南55个菌株测定, 筛选出9个单基因鉴别品种[12],但不适应于鉴别中国粕稻栽培地区的生理小种。 比较理想的鉴别方法是选育近等位基因系C7AF种[12]。在稻瘟病菌的变异研究上, 前人做了很多的工作。何秋月等[13]认为稻瘟病菌生理小种或致病类型多样性是寄
所谓分子育种,指的是在经典遗传学和现代分子生物学、分子遗传学理论指 导下,将现代生物技术手段整和于经典遗传育种方法中,结合表现型和基因型筛 选,培育优良新品种。目前,抗稻瘟病分子育种主要包括分子标记辅助选择和基 因工程育种。 3.1.3.1 分子标记辅助育种
传统的抗病育种主要通过抗性鉴定与植株表型选择来进行。这种方法不仅需 要育种家具有丰富的育种经验,同时也有很多限制因素,如稻瘟病发病的环境条 件不易控制,稻瘟病菌生理小种的稳定性很难把握和育种周期长等。利用分子标 记辅助技术(MAS)来选择,可通过分析与目标性状基因紧密连锁的分子标记来判 断基因的存在与否。因分子标记不受基因表达时 间、显隐性关系和环境条件及稻瘟病生理小种的影响,故可在早期进行选择,甚 至在播种前用不含胚的半粒种(含胚的半粒用于播种)借助分子标记进行选择。分 子标记辅助选择不仅减少了选择的盲目性,而且缩短了育种年限,大大提高了选 择效率。目前MAS已成功应用于抗稻瘟病基因的聚合育种。Zheng等(1995)[20]通 过MAS将抗稻瘟病基因Pi1, Pi2, Pi4聚合到同一品种中。Hittalmani[21] ( 2000)将 稻瘟病抗性基因Piz5,Pa1,Pita聚合到同一个品种。随着水稻基因组研究的迅速发 展,特别是基于PCR的多种方法和技术的不断开发运用,检测技术的简化和技术 成本的降低,MAS应用于性状改良的条件已日趋成熟。MAS和田间鉴定技术相结 合会极大地提高抗稻瘟病育种的选择效率。 3.1.3.2 抗稻瘟病基因工程育种
优异的广谱抗稻瘟病基因以及多抗性基因聚合育种。本文从稻瘟病的病原菌角度
出发,概述了稻瘟病菌的致病机理以及接种稻瘟病菌的方法,然后从水稻和稻瘟
病菌互作的角度概述水稻抗源筛选、抗性基因定位和无毒基因研究进展,并对育
种家改良水稻稻瘟病抗性的有效手段进行了总结。 2. 水稻稻瘟病菌的研究 2.1 稻瘟病病菌的致病机理
稻瘟病菌是稻梨孢菌,属半知菌亚门真菌[4]。日本学者最早发现了稻瘟病菌 的侵染过程,包括分生孢子的产生和萌发、附着胞形成、侵染栓形成以及次生菌 丝生长扩展。稻瘟菌以分生孢子和菌丝体在稻草和稻谷上越冬,翌年产生分生孢 子借风雨传播到稻株上,萌发侵入寄主并向邻近细胞扩展发病,形成中心病株。 病部形成的分生孢子,借风雨传播进行再侵染。稻瘟病侵入寄主主要是机械作用, Unchigama等[5]Rodrigues[6]等研究究认为稻瘟病菌分生孢子的完整形成和接触面 的蜡质层、角质层等疏水表面有关,蜡质能促进附着胞的形成。稻瘟病菌的侵染 存在诱导机制,李成云[7]研究认为稻瘟菌中的分生孢子发芽液中含有侵染诱导物 质,能使致病性菌株的发病指数增加,但目前并没有分离出这种活性物质。
杂交育种技术是抗稻瘟病育种常规的手段,也仍然是目前抗病育种主要的技 术之一。杂交育种是通过不同亲本间有性杂交产生遗传基因的重组,再经过若干 世代的性状分离,选择和鉴定来获得符合育种要求的新品种的方法。由于水稻抗 性遗传力因品种不同而有所差异,因此在进行抗病育种时要注意选用稻瘟病抗性 传递力强的亲本,同时还要选择各种农艺性状适 合本地特点的杂交亲本进行配组,才能较容易获得各种性状好的后代。如由湖南 省水稻研究所选育的抗稻瘟病优质早粕稻湘早143[17],江西农科院育成的赣晚籼2 7 。 [18] 3.1.2 诱变育种
苗期至抽穗期均可发生,苗期或者分蘖期发病严重时可使水稻植株死亡;穗期发
病将导致白穗或半饱和穗,大大降低产量,严重时可造成水稻绝收。稻瘟病病原
菌的生理小种多,并且变异复杂,对稻瘟菌群体的研究还停留在初步阶段。实践
证明,培育抗性品种是防治稻瘟病最经济、有效的方法。但由于稻瘟菌变异性强,
很多抗性品种种植几年就丧失了抗性[3]。解决这一问题的有效途径是分离和克隆
花药培养技术是快速培育抗病新品种的一种有郊途径,已被成功地应用于植 物的育种实践。与常规育利相比,花药培养除了能够缩短育种年限外,还具有以 下优点所获得的单倍体经染色体加倍,当代就能纯合程定,缩短了育种年限。单 倍体的自然加倍使两倍体一沙纯合化,后代系统遗传迅速稳定,有利于抗病类型 与撼病类型的鉴定与选拔,提高了选择效率。花培育种能仗各种配子类型在植株 水平较充分地显现出来,避免隐性基因为隐性基因的上位性作用,使隐性基因控 制的性状显示出来;花培育种出现纯合的抗病基因型的概率为1. 2%,大大高于常 规育种的概率;花培育种后什选择的群体(或系统)大小是常规育种的1/5~1/2。 但是,利用花药培养技术筛选抗性突变体的后代,筛选鉴定的工作量较大。此外, 花药培养的药源一般来自选抖的个体,而不是群体。如以双抗个体为药源,花培 后什出现纯合的抗病基因型的亲本基因组间发生重组的初会少于自交的重组会, 如果抗病基因与不良性状基因连锁,抗病基因型出现的机率将降低,此时可采用 在药培养的聚合育种法来打破不良连锁,以累积多个孚本的优良基因[23]。如黑龙 江省农科院水稻所于1982年以适于寒地生态条件的日本北海道优良品种松前为 母本,朝鲜早熟优质品种雄基9号为父本进行首次人工杂交。到1987年又用其稳 定后代龙交82133为母本,拜高抗稻瘟病的桥梁亲本N 193一做父本进行第二次六 工杂交。为缩短育种年限,提高选择效率,又于1988年将第二次杂交的F2代花药 进行人工接种离体培养,塌后选育出符合育种目标的单株。 3.1.4.2 细胞融合