抗体的基本结构
简述抗体的基本结构

简述抗体的基本结构抗体(又称免疫球蛋白)是一类重要的免疫分子,具有识别和中和病原体的能力。
抗体的基本结构由四个亚单位组成,包括两个轻链和两个重链。
本文将以简述抗体的基本结构为标题,详细介绍抗体的组成和结构特点。
1. 抗体的重链和轻链抗体的重链和轻链是由氨基酸序列组成的多肽链。
在人类的抗体中,重链有五种亚型,分别为IgM、IgG、IgA、IgD和IgE,而轻链有两种亚型,分别为kappa和lambda。
重链和轻链通过二硫键相连形成抗体的骨架结构。
2. 抗体的结构域抗体的结构域是指抗体分子上具有特定功能和结构的区域。
抗体的结构域包括可变区(variable region)和恒定区(constant region)。
可变区负责与抗原结合,决定了抗体的特异性;而恒定区则决定了抗体的功能和效应。
3. 抗体的可变区抗体的可变区由重链和轻链的氨基酸序列组成,其中包含了抗体的抗原结合位点。
可变区的序列在不同的抗体中表现出较高的变异性,这是由于免疫系统的基因重组和突变机制所致。
可变区的变异使得抗体能够识别多样性的抗原。
4. 抗体的恒定区抗体的恒定区在不同的抗体中表现出较低的变异性,这是由于恒定区的序列由基因家族所决定。
恒定区的序列决定了抗体的功能和效应,如中和病原体、激活免疫细胞和促进免疫效应等。
5. 抗体的抗原结合位点抗体的抗原结合位点是指抗体与抗原结合的区域,也称为抗体的亲和位点。
抗原结合位点由可变区的氨基酸序列决定,具有高度的特异性。
抗体通过与抗原结合来识别和中和病原体,从而发挥免疫效应。
6. 抗体的异构型抗体的异构型是指抗体分子中恒定区的结构差异。
在不同的抗体亚型中,恒定区的结构会发生变化,从而影响抗体的功能和效应。
例如,IgM抗体在结构上较大,能够聚集形成多价抗体,增强其中和病原体的效果。
7. 抗体的二级结构抗体的二级结构主要由α螺旋和β折叠构成。
重链和轻链中的可变区和恒定区都具有这种二级结构。
这种特殊的二级结构使得抗体能够具有稳定的空间构象,并保持其与抗原的特异性相互作用。
抗体的基本结构和功能

抗体的基本结构和功能介绍抗体(又称免疫球蛋白)是一种由免疫细胞产生的蛋白质,广泛存在于人体的血液和组织液中。
抗体在人体的免疫系统中起着至关重要的作用,能够识别和中和病原体、调节免疫反应、参与细胞间信号传导等。
本文将详细介绍抗体的基本结构和功能,以便更好地理解免疫过程和临床应用。
一、抗体的结构1.1 Fab和Fc区域抗体由两个相同的轻链(light chain)和两个相同的重链(heavy chain)组成。
每条轻链和重链都由一系列氨基酸残基连接而成,形成抗体的基本结构。
在抗体分子中,Fab(antigen binding fragment)区域负责与抗原结合,Fc(fragment crystallizable)区域则负责与免疫细胞相互作用。
1.2 IGH和IGL基因抗体的结构由基因编码决定,人体中有数百个IGH(immunoglobulin heavy chain)和IGL(immunoglobulin light chain)基因,它们通过基因重排和突变形成多样的抗体。
IGH基因编码重链的变量(V)区域、多样(D)区域、连接(J)区域和常规(C)区域,而IGL基因编码轻链的V区域和C区域。
1.3 亲和力成熟抗体的变量区域包含了可以识别和结合抗原的亲和力决定区(CDR, complementarity-determining region)序列,这些序列的组合能够使抗体与多种抗原结合并启动免疫反应。
亲和力的形成是通过基因突变和选择过程中的变异和筛选完成的,亲和力成熟是个体免疫系统应对病原体进化的重要机制。
二、抗体的功能2.1 识别和中和病原体抗体通过其变量区域与抗原结合,从而识别和中和潜在的致病病原体。
当抗体与抗原结合时,可以阻止病原体侵入宿主细胞、中和细菌毒素、聚集病毒颗粒等。
这一过程对于防御感染和预防疾病的发生起着重要作用。
2.2 调节免疫反应抗体不仅能够识别和中和病原体,还能够调节免疫反应的进程。
高中生物抗体知识点归纳总结

高中生物抗体知识点归纳总结一、抗体的基本概念抗体,全称为免疫球蛋白抗体,是由B淋巴细胞分泌的一类具有免疫功能的蛋白质。
它们能够识别并结合特定的抗原(如细菌、病毒、异物等),从而发挥免疫防御作用。
抗体主要存在于血清中,但也可以在组织液和外分泌液中找到。
二、抗体的结构抗体由四条多肽链组成,包括两条重链和两条轻链。
轻链和重链通过二硫键连接形成Y字形结构。
在Y字形的两个臂端部分,存在一个可变区,称为抗原结合位点,是抗体与抗原特异性结合的部位。
抗体的另一端,即Fc区,与免疫细胞上的Fc受体结合,参与免疫反应的调节。
三、抗体的分类根据结构和功能的不同,抗体可以分为五大类:IgA、IgD、IgE、IgG 和IgM。
各类抗体在免疫反应中扮演不同的角色。
例如,IgA主要存在于粘膜表面,保护粘膜免受病原体侵害;IgE与过敏反应有关;IgG是血清中含量最高的抗体,具有广泛的免疫功能;IgM是初次免疫应答时产生的第一种抗体,具有很强的抗原结合能力。
四、抗体的产生抗体的产生是适应性免疫反应的一部分。
当病原体侵入人体时,B淋巴细胞能够识别并结合到病原体上的抗原。
通过一系列的细胞活化、增殖和分化过程,B淋巴细胞转化为浆细胞,开始大量分泌抗体。
同时,部分B细胞成为记忆B细胞,长期存在于体内,为未来可能的再次感染提供快速响应。
五、抗体的功能抗体的主要功能包括中和、凝集、沉淀、补体激活和抗体依赖性细胞介导的细胞毒性(ADCC)等。
通过这些功能,抗体能够直接或间接地清除病原体,保护机体免受感染。
六、抗体的应用在医学领域,抗体被广泛应用于疾病的诊断和治疗。
例如,单克隆抗体技术可以制备特异性强、纯度高的抗体,用于治疗癌症、自身免疫疾病等。
此外,抗体还可以作为诊断试剂,帮助检测病原体或疾病标志物。
七、抗体与免疫调节抗体不仅能够清除病原体,还能够调节免疫系统的功能。
例如,某些抗体能够通过调节T细胞的活性,影响免疫应答的强度和持续时间。
此外,抗体还能够参与免疫耐受的形成,防止免疫系统对自身组织的攻击。
简述抗体的基本结构和生物学功能

简述抗体的基本结构和生物学功能抗体,也称为免疫球蛋白,是一种由哺乳动物免疫系统产生的蛋白质分子,具有多种结构和功能。
抗体具有重链和轻链组成,每个抗体分子由两个重链和两个轻链组成,形成Y形状。
抗体的基本结构包括可变区和恒定区,可变区决定了抗体的特异性,恒定区则决定了抗体的生物学功能。
抗体的结构可以分为四个区域:两个抗原结合部位,一个球部和一个棒部。
抗原结合部位位于抗体的顶端,并与抗原结合形成特异性复合物。
抗原结合部位的可变区域由重链和轻链的V区域共同决定,具有高度多样性,可以识别并结合多种抗原。
抗体的球部由重链和轻链的C区域组成,决定了抗体的种类和亚类。
棒部由抗体的重链的C区域组成,可与机体免疫细胞相互作用。
抗体的生物学功能包括中和病原微生物、沉淀抗原、激活补体系统、识别和标记异物、调节免疫应答等。
抗体可以通过与病原微生物的抗原结合来中和病原微生物,阻止其侵入机体细胞。
抗体还可以与抗原结合形成沉淀复合物,促使病原微生物和抗原沉淀而不再对机体产生损害。
抗体还可以与补体系统相互作用,激活补体系统来清除病原微生物。
此外,抗体还可以识别和标记异物,使其易于被机体免疫细胞识别和清除。
此外,抗体还可以调节免疫应答,通过与抗原结合来激活或抑制其他免疫细胞的功能,调节免疫应答的强度和方向。
抗体的生物学功能还可以通过其结构的多样性和可选择性来实现。
抗体的可变区域具有高度多样性,可以识别和结合多种抗原,因此可以用于特异性识别和治疗多种疾病。
抗体还可以通过亲和力成对的方式结合抗原,形成二聚体或多聚体,增强抗体的结合力和生物学功能。
总之,抗体具有重链和轻链组成的Y形结构,包括可变区和恒定区。
抗体的主要生物学功能包括中和病原微生物、沉淀抗原、激活补体系统、识别和标记异物、调节免疫应答等。
抗体的多样性和可选择性使其成为免疫系统中重要的分子,具有广泛的应用前景。
抗体的c区名词解释

抗体的c区名词解释抗体,也被称为免疫球蛋白,是一种由免疫细胞合成并在免疫系统中起关键作用的蛋白质。
它们被广泛应用于医学诊断、疾病治疗和生物科学研究。
抗体的结构复杂多样,其中一个重要的组成部分是C区。
一、抗体的基本结构抗体由四个亚基组成,包括两个重链和两个轻链。
每个重链和轻链都包含一个可变区(V区)和一个常规区(C区)。
C区位于重链和轻链的非抗原特异性区域,也称为常规亚基。
C区在抗体的功能和稳定性中起到至关重要的作用。
二、C区的功能C区在抗体中具有多种功能。
首先,它是抗体和其他免疫细胞(如巨噬细胞和自然杀伤细胞)相互作用的关键部位。
C区参与了抗体与这些细胞表面受体的结合,从而介导了免疫应答过程中的信号传导和效应。
这种相互作用可以激活免疫细胞,引发炎症反应并促进病原体的清除。
此外,C区还参与了抗体的结构稳定性和半衰期调控。
具有较长半衰期的抗体对抗原的清除能力更强,因此对于抗体的疗效至关重要。
C区通过与其他免疫分子的相互作用,影响了抗体的稳定性和其与抗原结合的力度。
三、C区的亚型和功能分化在不同的抗体亚型中,C区的结构和功能可能有所不同。
人体中主要存在着五种抗体亚型,即IgM、IgG、IgA、IgD和IgE。
每种亚型的C区都有独特的结构特征,决定了它们在免疫系统中的功能。
例如,IgM的C区结构较长,形成了一种“五座桥梁”的结构,使其能够更有效地激活免疫细胞。
IgG则具有较长的半衰期,能够在循环系统中稳定存在较长时间,并在炎症反应中发挥重要的调节作用。
IgA主要存在于黏膜表面,提供保护,并参与呼吸道和消化道的免疫防御。
由于C区的亚型差异,抗体可以执行不同的功能。
这为科学家研究抗体的功能和开发相关治疗提供了重要的线索。
四、抗体工程和C区的关系近年来,抗体工程的研究和应用取得了突破性进展。
科学家们能够通过改变抗体的C区结构和序列,实现抗体的改进和优化。
例如,通过工程改变抗体C区的氨基酸序列,可以增强抗体与免疫细胞受体的结合亲和力,提高抗体的活性。
免疫学第三章抗体
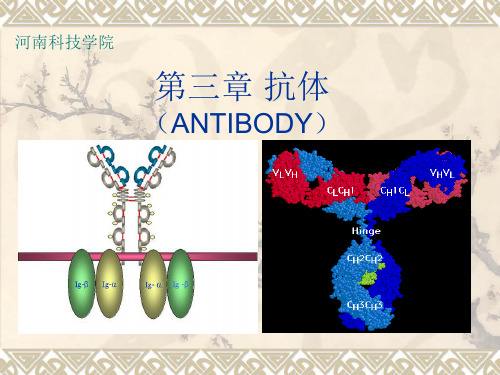
IgG1 (kappa)
IgM (kappa)
(2)同种异型(allotype)
❖ 同种系的不同个体之 间,同一类、亚类、型、 亚型的免疫球蛋白, 在结构上可能会有一 个或数个氨基酸的差 异, 称为同种异型。
❖ 由同一基因座位上不 同等位基因编码。
排列顺序以及结构是随 抗体分子的特异性不同而 有所变化,决定了抗体的 特异性。
重链的可变区和恒定区
❖ 恒定区(constant region, C区):可变区外其余 的氨基酸分子。(占重 链的3/4)。
数量、种类、排列 顺序及含糖量都比较 稳定,称为稳(恒)定 区。
轻链的可变区和恒定区
❖ 可变区(V区):氨基
❖ 免疫球蛋白的存在形式:
免疫球蛋白除分布于体液中之外,还可 存在于B细胞膜上。
❖ 膜型(membrane immunoglobulin, mIg)
--- B细胞膜上的抗原受体;
❖ 分泌型(secreted immunoglobulin, sIg)
---分泌进入体液, 介导体液免疫应答。
2、免疫球蛋白的基本结构
即抗原结合片段。
Fc 片段(fragment crystallizable,Fc):在
低温或低离子强度下可形成结晶,故称为可 结晶片段。
❖ Fab 片段:
每一个含有一条 完整的 L 链和 H 链近 N 端侧 的 1/2 。
❖ Fc 片段:
含有两条 H 链 羧基端(C 端) 的一半。
❖ Fab 片段:
❖ 五 类 免 疫 球 蛋 白 IgG , IgM , IgA , IgE , IgD的单体分子结构都是相似的,即是由 两条相同 H 链和两条相同 L 链共四条肽 链构成的“Y”字形的分子。
抗体的结构

氨基端
抗
原
结
合 部 位
VL
生 物
铰链区
学
活ቤተ መጻሕፍቲ ባይዱ
性
区
VH
CH 2 CH 3 羧基端
重链 轻链
二硫键 补体结合部位 Fc受体结合部位
抗体的结构
抗体的概念
抗体(antibody,Ab)是由B淋 巴细胞在抗原刺激下增殖分化为 浆细胞所产生,是介导体液免疫 的重要效应分子,能与相应抗原 发生特异性结合的免疫球蛋白。
主要分布在血液中,也分布于 组织液及外分泌液中。
免疫球蛋白(immunoglobulin,Ig): 将具有抗体活性或化学结构与抗体相似的球蛋白统称为免 疫球蛋白。
恒定区(constant region, C区):
氨基酸的组成和顺序 相对稳定。
可变区:
① 高变区 (hypervariable region, HVR):氨基酸组成和
顺序变化大。又称为互补性决定 区(complementarity determining region,CDR)。
② 骨架区(framework region, FR):氨基酸组成和
lg为糖蛋白,糖基存在于重链上。
重链(H链):
分子量约 50-75 kD,由450~570个氨基酸残基 组成
五种不同种类的重链:分别为γ链、 μ链、 α链、 δ链和ε
链,其氨基酸种类和排列顺序不同,抗原特异性不同。
免疫球蛋白分为五类:IgG、IgM、IgA、IgD和IgE。
轻链(L链):
功能区的作用: ① VH 和VL 是结合抗原的部位。 ② CL 和CH上具部分遗传标记(同种异
型)。 ③补体结合位点( CH2 或CH3 )。 ④ CH3 或CH4与免疫细胞表面相应受体
简述抗体的基本结构

简述抗体的基本结构抗体是一种由蛋白质组成的分子,也称为免疫球蛋白。
它在免疫系统中起着至关重要的作用,能够识别并结合到体内外的抗原上,从而参与免疫应答的调节和效应。
抗体的基本结构可以分为四个区域:两个相同的轻链(light chain)和两个相同的重链(heavy chain)。
轻链和重链通过二硫键连接在一起,形成了Y形结构。
每个抗体分子都有两个臂和一个柄。
臂部位于抗体的顶端,具有抗原识别和结合的功能;柄部位于抗体的底部,具有效应分子结合的功能。
轻链是由单个多肽链组成的,重链则由两个多肽链组成。
轻链和重链都由一系列的可变区(variable region)和恒定区(constant region)组成。
可变区的序列在不同的抗体中具有高度的变异性,决定了抗体的特异性和亲和力。
恒定区的序列相对保守,决定了抗体的功能和效应。
抗体的可变区包括特异性决定区(hypervariable region),也称为CDR。
CDR位于可变区的末端,由三个短序列组成,分别命名为CDR1、CDR2和CDR3。
CDR通过氢键、疏水作用和范德华力等相互作用与抗原结合,形成抗原-抗体复合物。
抗体的恒定区在不同的抗体类别中有所不同,包括IgG、IgM、IgA、IgD和IgE等。
每种抗体类别具有不同的功能和在免疫反应中的作用。
例如,IgG是最常见的抗体类别,具有长时间的循环寿命和多种效应分子结合的能力;IgM是第一次免疫应答产生的主要抗体类别,具有较高的亲和力和多价结合能力。
在人体内,抗体由B淋巴细胞(B lymphocyte)产生。
当机体遇到外来的抗原时,B淋巴细胞会受到激活,开始分泌抗体。
这个过程称为免疫应答。
一旦抗体与抗原结合,它们可以通过多种机制来识别和清除抗原,包括中和病原体、激活补体系统、调节免疫细胞的活化等。
抗体的结构和功能使其成为疾病诊断和治疗的重要工具。
通过检测体液中的抗体水平,可以确定感染病原体的存在,并评估免疫系统对其的应答情况。
抗体的名词解释微生物学

抗体的名词解释微生物学在微生物学领域,抗体是研究免疫系统的关键组成部分。
抗体,也称免疫球蛋白,是机体免疫系统中一类具有高度特异性的蛋白质。
它们由免疫细胞分泌,并在机体中起到识别、结合和中和外来病原体的作用。
一、抗体的基本结构抗体是由两个基本结构单元组成的,即重链和轻链。
其中,重链分为四种类型:IgG、IgM、IgA和IgE,轻链分为两种类型:κ链和λ链,这些基本结构单元将不同组合形成丰富多样的抗体。
抗体还具有“Y”字形的三维结构,其中“Y”字的两端为抗原结合位点,而“Y”字的顶端则是抗体的效应位点。
二、抗体的功能抗体具有多种功能,对于攻击和保护机体免受外来微生物的侵害起到了重要作用。
首先,抗体能够通过识别外来微生物表面的抗原决定簇结合到病原体上,从而标记该病原体以便免疫系统进一步处理。
其次,抗体还能激活免疫系统中的巨噬细胞、自然杀伤细胞和补体系统等,引发炎症反应和细胞毒性作用,以消灭病原体。
此外,抗体还能参与免疫记忆和免疫调节等免疫过程,对于维持机体内部的免疫平衡起到重要作用。
三、抗体的产生抗体的产生主要依赖于机体的免疫细胞,特别是淋巴细胞。
当机体受到外来病原体感染时,抗原会被呈递到淋巴组织中的抗原呈递细胞上,这些抗原呈递细胞会将抗原信息传递给免疫细胞。
免疫细胞中的B淋巴细胞随即被激活,并开始产生和分泌抗体。
这一过程被称为体液免疫应答,产生的抗体会迅速进入血液循环,通过对病原体进行中和,起到阻止病原体侵入和扩散的作用。
四、抗体的应用抗体不仅在免疫系统中起到重要作用,在医学和生物工程等领域也具有广泛应用。
例如,抗体可以用于诊断某些病原体感染,通过检测抗体水平来判断是否感染。
此外,抗体还可以用于治疗某些疾病,如恶性肿瘤。
在生物工程领域,科学家们利用重组技术生产抗体药物,以满足临床上的需求。
五、抗体研究的发展随着科学技术的不断发展,抗体研究也在不断深入。
科学家们通过对抗体结构和功能的深入了解,不断探索新的抗体工程方法,以提高抗体的亲和力和特异性。
抗体的基本结构

抗体的基本结构抗体是一种特殊的抗原分子,它与外界的其他微生物、抗原或毒素发生反应,对其进行抵抗和清除。
抗体的基本结构是指它的分子组成,即它的结构与功能,以及它们与其环境的相互作用。
抗体是分子组成的复杂结构,其中包含了大量的亚基(subunit)。
抗体由两个不同的抗原结合起来,形成一个双聚体,称为抗体双聚体(antibody dimer)。
每个单元都有其自身的特殊功能,它们在体内结合起来,共同发挥抗体的功能。
抗体双聚体由两个亚基(heavy chain subunit和light chain subunit)组成,它们互相错开,形成一个带有特殊形状的V字型结构。
重亚基包含了四个结构域:FCR、VH、VL和CDR,这四个域分别负责了抗体的活性、特异性、与受体的结合等功能。
轻亚基也由 FCR、VL和CDR三个域组成,分别负责多样性、受体结合和活性等功能。
抗体双聚体的结构非常稳定,而且它能够结合其他抗原,从而实现免疫反应。
在体内,抗体可以与抗原结合,并发生特异性的免疫反应。
该反应的特异性对于抗体的结构也有很大的影响,包括FV结构的稳定性、VH和VL的位置变化、CDR的变形以及FCR的连接方式等。
当抗体双聚体与抗原结合时,尤其是在FCR中,会发生位置变化,从而使抗体双聚体发生折叠改变,从而改变抗原抗体复合物的稳定性。
此外,抗体双聚体还能够与特定的受体结合,该受体可以有助于结合其他分子,从而发挥免疫反应的功能。
受体与抗体双聚体之间的结合依赖于受体上的特定序列,以及其与抗体双聚体之间的相互作用力。
该结合可以通过特异性结合(特异性结合)和非特异性结合(交联结合)两种方式发生,这种结合对于抗原抗体反应有着重要的作用。
抗体的基本结构功能在于它们与其他抗原之间协同发挥的功能,它们的特殊结构可以结合抗原,促进抗原与受体之间的相互作用,而这种相互作用可以促进抗体与抗原之间的免疫反应。
它们可以与受体结合,从而实现对外界抗原的识别,并发挥免疫功能。
抗体的结构组成

抗体的结构组成抗体是人体免疫系统中的重要组成部分,它可以识别和结合抗原,从而参与免疫应答。
抗体的结构组成复杂多样,包括基本的单体结构、多样性区和亚类等。
本文将详细介绍抗体的结构组成。
1. 抗体的基本结构抗体分为五个类别,即IgM、IgD、IgG、IgA和IgE。
它们都由相似的基本结构单元组成,包括两个重链和两个轻链。
这两种链均具有一系列的区域,包括甲胎蛋白区(variable region,V区)和常规区(constant region,C区)。
重链有两类:“μ链”和“δ链”是IgM和IgD的特有重链;“γ链”是IgG的特有重链;“α链”是IgA的特有重链;“ε链”是IgE的特有重链。
轻链有两类:“κ链”和“λ链”。
2. 抗体的多样性区抗体的多样性区域是决定其特异性的关键部分。
抗体中的多样性区域包括重链和轻链的V区域。
抗体的V区域由多个互补决定区(complementarity-determining region,CDR)组成。
CDR是抗体中最具变异性的区域,决定了抗体与特定抗原结合的能力。
抗原结合位点通常位于抗体的VH和VL区域之间,也被称为互补决定区。
3. 抗体的细节结构(1)抗体的Fc区:抗体的Fc区是指抗体中的常规区(C区)和C区之间的一段区域。
Fc 区的结构差异决定了抗体亚类的差异。
例如,IgM和IgE的Fc 区有一个长的C 区段,而IgA的Fc 区有一个有四个C 区段的结构。
(2)抗体的Fab区:抗体的Fab 区是指抗体中的V区和C区的连接部分。
Fab 区包括两个抗原结合片段(F(ab’)2)。
每个Fab片段有一个抗原结合区和一个连接区。
4. 抗体的亚类抗体的重链的C区决定了抗体的亚类。
不同的抗体亚类在结构和功能上有所差异。
IgM是体液中第一次接触到抗原时产生的抗体。
它是由5个单体抗原片段(F(ab)2)加一个乳糖片段(Fc)组成的。
IgG是体液中最主要的抗体,也是体外免疫应答中最重要的抗体。
抗体的基本结构、功能区划分与各区功能

抗体的基本结构、功能区划分与各区功能下载提示:该文档是本店铺精心编制而成的,希望大家下载后,能够帮助大家解决实际问题。
文档下载后可定制修改,请根据实际需要进行调整和使用,谢谢!本店铺为大家提供各种类型的实用资料,如教育随笔、日记赏析、句子摘抄、古诗大全、经典美文、话题作文、工作总结、词语解析、文案摘录、其他资料等等,想了解不同资料格式和写法,敬请关注!Download tips: This document is carefully compiled by this editor. I hope that after you download it, it can help you solve practical problems. The document can be customized and modified after downloading, please adjust and use it according to actual needs, thank you! In addition, this shop provides you with various types of practical materials, such as educational essays, diary appreciation, sentence excerpts, ancient poems, classic articles, topic composition, work summary, word parsing, copy excerpts, other materials and so on, want to know different data formats and writing methods, please pay attention!一、引言抗体作为免疫系统中极为重要的一部分,是机体对抗外源性病原体的关键保护因子。
抗体的化学本质什么

抗体的化学本质什么一、前言抗体具有非常重要的生物学意义和医学价值,是生命科学中的重要研究对象。
抗体涉及到多个学科领域,其中最为核心的是生物化学。
抗体是由生物化学反应生成的一类大分子,具有很强的特异性和亲和力,广泛应用于生物研究、药物研发和临床诊断等领域。
本文将从抗体的基本结构出发,介绍抗体的化学本质及其相关原理。
二、抗体的基本结构(一)抗体的分类抗体分为五种不同类型,即IgG、IgM、IgA、IgD和IgE。
它们在结构和功能上有所不同,但都由重链和轻链组成。
(二)重链和轻链重链和轻链分别是抗体的两个组成部分,重链其C末端有一个糖基化的尾部,即酵母菌结构域,轻链没有。
重链和轻链分别由N端变异率区间(VH和VL)和四个保守区域(CH1、CH2、CH3和CL)组成。
(三)抗体的基本结构抗体的基本结构由两个抗原结合面、两个轻链和两个重链组成。
重链和轻链通过二硫键连接,形成一个Y形结构。
其中两个轻链连接在Y型结构的底部,形成一个小臂,而两个重链连接在Y型结构的顶端,形成一个重臂,重臂和小臂都有一个抗原结合面。
三、抗体的化学组成抗体是一种免疫球蛋白,其化学组成非常复杂,包括氨基酸、糖、核酸等多种分子。
下面分别介绍抗体的化学组成。
(一)氨基酸氨基酸是组成抗体的基本单位,抗体总共包含20种氨基酸。
其中,一些特殊的氨基酸如半胱氨酸、天冬氨酸、缬氨酸等在抗体的结构和功能中起着重要作用。
氨基酸序列的变化使得不同抗体具有不同的结构和特异性。
(二)糖糖是抗体中另一个重要的结构组分。
其中,N-糖基化是一种常见的修饰方式,包括尾糖基、复杂糖基和高岭糖基等。
这些糖基通过与细胞表面上的受体结合或与其他蛋白质相互作用来调节抗体的生物学功能。
(三)核酸在抗体基因的重排和突变过程中,DNA和RNA扮演着重要角色。
这些过程使得不同的抗体分子可以产生,具有不同的结构和特异性。
四、抗体的化学本质抗体具有很高的特异性和亲和力,这取决于其化学本质。
抗体知识点总结

抗体知识点总结一、抗体的基本概念抗体(antibody),也称免疫球蛋白,是机体免疫系统中的一种重要蛋白质,由免疫球蛋白和其他蛋白质组成。
抗体主要由B细胞产生,在免疫系统中起着重要的作用,可以识别并结合到抗原分子,并进行中和、沉淀、激活补体等免疫反应。
抗体的结构复杂,可以分为五个类别(IgM、IgG、IgA、IgD、IgE),每种类别具有不同的功能和特点。
二、抗体的结构1. 抗体的总体结构抗体的基本结构由两条重链和两条轻链组成,重链和轻链通过二硫键连接在一起,形成一条“Y”形的结构。
每个抗体分子上有两个抗原结合位点,可以与抗原特异性结合。
2. 抗体的免疫球蛋白结构每个抗体分子由一个具有特异性的抗原结合区域和一个常规结构的Fc区域组成。
抗原结合区域由重链和轻链上的可变区域共同组成,具有高度的多样性,可以与不同的抗原结合。
Fc区域由重链上的常规区域组成,具有一定的生物学功能。
3. 抗体的多样性抗体的多样性主要来源于其抗原结合区域的可变区域,每个抗体分子可以结合不同的抗原。
三、抗体的功能1. 中和作用抗体可以结合到细菌、病毒等病原微生物上,阻止其进入宿主细胞,从而起到中和病原微生物的作用。
2. 激活补体抗体结合到抗原上可以激活补体系统,引发细胞溶解、炎症反应等生物学效应。
3. 免疫沉淀抗体与抗原结合形成免疫复合物,可以沉淀在组织中,起到清除抗原的作用。
4. 刺激B细胞抗体与抗原结合后可以刺激B细胞产生更多的抗体,从而增强免疫反应。
5. 细胞毒作用某些抗体可以结合到靶细胞表面,引发细胞毒作用,促使细胞凋亡或溶解。
四、抗体的生成过程1. 抗原识别当机体内部或外部出现抗原刺激时,B细胞中的抗原受体可以识别并结合到抗原,激活B 细胞。
2. B细胞激活被激活的B细胞会开始增殖并分化成浆细胞和记忆B细胞。
3. 浆细胞产生抗体浆细胞是产生抗体的细胞,它可以大量合成和分泌特异性抗体。
4. 记忆B细胞记忆B细胞可以长期存留在机体内,当再次遇到相同的抗原时,可以迅速产生抗体,加强免疫反应。
简述抗体的基本结构

简述抗体的基本结构抗体,也称为免疫球蛋白,是一种由免疫系统产生的蛋白质分子,主要功能是识别和结合外来物质,以便将其标记并引发免疫反应。
抗体的基本结构由四个多肽链组成,包括两条重链和两条轻链。
本文将从整体结构、重链和轻链的组成以及可变区和恒定区的功能等方面简述抗体的基本结构。
抗体的整体结构可分为两个基本部分,即Fab区和Fc区。
Fab区是抗体的抗原结合部位,由两个可变区域(VH和VL)组成,分别位于重链和轻链的N端。
Fab区的可变区域能够与抗原结合,因此每个抗体能够结合特定的抗原。
Fc区则是抗体的结构稳定区域,由两条重链的C端组成,是抗体的效应部位,能够与免疫细胞或其他分子相互作用,引发免疫反应。
重链是抗体的一条多肽链,分为五个类别:IgM、IgG、IgA、IgD 和IgE。
每个类别的重链都有其特定的功能和分布。
重链的C端由恒定区组成,决定了抗体类别的特性。
重链的N端则由可变区组成,决定了抗体的抗原结合特异性。
重链可通过二硫键与轻链相连,共同形成抗体的整体结构。
轻链是抗体的另一条多肽链,分为两个类别:κ链和λ链。
轻链与重链相连的部分也包括恒定区和可变区。
轻链的可变区与重链的可变区共同形成抗原结合位点,决定了抗体的抗原结合特异性。
轻链的恒定区则决定了抗体的类别。
抗体的可变区是抗体结构中最为关键的部分,也是抗体的抗原结合位点。
可变区的序列和结构高度多样,能够与各种不同的抗原结合。
可变区的多样性是由基因重组和突变引起的,使得免疫系统能够识别和结合各种外来物质。
而恒定区则决定了抗体的类别和功能,包括了促进免疫细胞的吞噬作用、激活补体系统等功能。
抗体的基本结构由四个多肽链组成,包括两条重链和两条轻链。
重链和轻链通过二硫键相连,形成抗体的整体结构。
重链的C端决定了抗体的类别和功能,N端的可变区决定了抗体的抗原结合特异性。
轻链的恒定区决定了抗体的类别,可变区与重链的可变区共同形成抗原结合位点。
抗体的基本结构为其在免疫系统中发挥重要功能提供了基础,也为我们理解抗体的作用机制提供了重要线索。
抗体的结构与功能

抗体的产生:由B细胞在抗原刺激下产生 抗体的种类:IgG、IgM、Ig、IgE、IgD等 抗体的调节:由T细胞和B细胞相互作用调节 抗体的寿命:短寿命抗体和长寿命抗体 抗体的效应:中和作用、调理作用、补体结合作用等
自身免疫性疾病: 如系统性红斑狼疮、 类风湿性关节炎等
过敏性疾病:如 哮喘、过敏性鼻 炎等
抗体在生物技术中的应用:用于基因工程、细胞工程、酶工程等生物技术的研究与开发
抗体药物的研发:通过基因工程技术 将抗体基因导入到细菌、酵母或哺乳 动物细胞中进行大规模生产
抗体药物的前景:随着基因工程技 术的不断发展抗体药物的研发和应 用将更加广泛和深入
添加标题
添加标题
添加标题
添加标题
抗体药物的应用:用于治疗癌症、 自身免疫性疾病、感染性疾病等
抗体与自身免疫疾病相关可能 导致自身免疫性疾病
参与免疫调节:抗体可以调节免疫 反应防止免疫系统过度反应
参与细胞分化和增殖:抗体可以影 响细胞的分化和增殖影响细胞的生 长和发育
添加标题
添加标题
添加标题
添加标题
参与细胞信号传导:抗体可以与细 胞表面的受体结合传递信号影响细 胞功能
参与细胞凋亡:抗体可以诱导细胞 凋亡清除受损或异常细胞维持机体 健康
抗体的产生和调节
抗原刺激:抗原进入机体刺激免疫系统产生抗体 B细胞活化:抗原刺激B细胞使其活化并开始产生抗体 抗体生成:B细胞通过基因重排和表达生成特异性抗体 抗体分泌:抗体通过B细胞的分泌途径释放到血液中 抗体调节:免疫系统通过反馈机制调节抗体的产生和分泌
亲和力成熟:抗体与抗原结合的能力逐渐增强的过程 类别转换:抗体从一种类别转变为另一种类别的过程 免疫球蛋白重链和轻链的基因重排:抗体产生的基础 抗体的亲和力成熟和类别转换的机制:免疫系统对病原体的识别和反应
简述抗体的基本结构和生物学功能

简述抗体的基本结构和生物学功能抗体,也被称为免疫球蛋白(immunoglobulin, Ig),是一种由免疫系统产生的特殊蛋白质,主要作用是识别和中和体内外的病原体,保护机体免受感染。
抗体的基本结构和生物学功能是免疫系统最重要的组成部分之一抗体的基本结构包括四个多肽链,分为两对相同的轻链(light chains)和重链(heavy chains)。
每条轻链和重链由一系列不同的氨基酸组成,通过二硫键链接在一起形成抗体的Y形结构。
轻链分为κ(kappa)和λ(lambda)两种类型,重链则分为五个不同的类别:IgG、IgM、IgA、IgD和IgE。
抗体的Y形结构由两个抗原结合部位(Fab)和一个结构相同的晶体抗体组(Fc)组成。
Fab部分是抗体与抗原特异性相互作用的区域,每个Fab部分都包含一个完整的轻链和一半的重链。
Fab与抗原结合后,可以阻止病原体进入机体细胞或标记其以便被免疫系统识别和消除。
Fc区域位于抗体的尾部,包含另一半的重链,不直接与抗原相互作用,但是可以与其他细胞或分子结合,介导一系列生物学效应,如调理细胞介导的毒杀(ADCC)和补体激活。
抗体的生物学功能主要包括以下几个方面:1. 识别和结合抗原:抗体可以通过它的Fab区域与抗原(比如细菌、病毒、肿瘤细胞等)结合。
抗体与抗原之间的结合是高度特异性的,由抗体的可变区决定。
抗体与抗原的结合可以阻止病原体进入机体细胞,中和病原体毒素或标记它们以被免疫系统识别和清除。
2.补体激活:抗体的Fc区域可以识别和结合补体蛋白,进而激活免疫系统中的补体级联反应。
补体激活可以引发炎症反应和细胞溶解,直接杀伤病原体或增强其他免疫细胞的杀伤作用。
3.调理细胞介导的毒杀:抗体的Fc区域可以结合调理细胞上的Fc受体,如自然杀伤细胞(NK细胞)。
这种结合可以激活调理细胞,使其释放细胞毒性因子,直接杀伤抗原表达的细胞。
4.免疫记忆:抗体参与免疫系统的记忆反应。
当机体第一次暴露在特定抗原时,B淋巴细胞会产生和分泌相应的抗体。
抗体的基本结构和功能

抗体的基本结构和功能
一、引言
抗体是一种重要的免疫分子,具有广泛的生物学功能。
本文将介绍抗体的基本结构和功能。
二、抗体的基本结构
1. 抗体的种类
抗体分为五种类别:IgG、IgM、IgA、IgD和IgE。
2. 抗体的基本结构
抗体由两个重链和两个轻链组成,每条链都包含一个可变区域和一个恒定区域。
可变区域决定了抗体与特定抗原结合的能力,恒定区域则决定了抗体的生物学活性。
3. 抗原结合部位
每个抗体分子都有一个特异性决定区(CDR),也称为互补决定区
(CDR)。
CDR是由可变区域组成的,其序列决定了抗体与特定抗原结合的能力。
三、抗体的功能
1. 中和作用
中和作用是指通过与病毒或细菌表面上的特异性受体相互作用,阻止
它们进入宿主细胞并进行复制。
2. 凝集作用
凝集作用是指通过与多个细胞或颗粒相互作用,使它们聚集在一起形
成大块,从而更容易被免疫系统识别和清除。
3. 激活补体系统
抗体可以激活补体系统,引起炎症反应和细胞溶解。
4. 识别和标记作用
抗体可以识别和标记它们所结合的细胞或颗粒,从而被免疫系统清除。
5. 调节免疫反应
抗体可以调节免疫反应的强度和方向,从而保持机体内部环境的稳定性。
四、结论
抗体是一种重要的免疫分子,具有广泛的生物学功能。
通过了解其基本结构和功能,我们可以更好地理解免疫系统如何工作,并为开发新型治疗方法提供指导。
抗体基本结构

抗体基本结构抗体是一种能够识别和结合特定抗原的蛋白质,是人体免疫系统中的重要成分之一。
抗体的基本结构对于人们深入了解其功能、特性及其应用具有非常重要的作用。
从以下几个方面,介绍一下抗体的基本结构。
1. 抗体的区域抗体包含两个区域,一个是变区,另一个是常量区。
变区赋予抗体识别和绑定不同抗原的能力,常量区则规定了抗体的种类和生物学功能。
2. 抗体的种类目前已知的抗体种类有五种(IgM、IgG、IgA、IgD和IgE),它们的常量区结构不同,具有不同的生物学特性与功能。
IgM抗体是活化B细胞的第一响应者,IgG抗体可以穿越胎盘保护胎儿,IgA抗体可以呈现在分泌液中,抗细菌和病毒,IgD与IgM同在B细胞的表面,研究人员对IgE的特性最近甚嚣尘上,因为它与过敏反应有关。
3. 抗体的基本单元抗体基本单元是由四个多肽链组成的。
两条重链和两条轻链形成Y形结构。
重链和轻链中每一条均由变区和常量区组成。
变区可以识别特异性抗原,并使抗体与之紧密结合。
4. 抗体特异性抗体的特异性是由重链和轻链的变区定义的。
变区由大量不同的基因编码,使得抗体在识别抗原时表现出高度的特异性。
当发生呼吸道感染时,抗体变区可以特异性地结合病原体细胞壁上的多糖蛋白。
5. PentamerIgM是唯一的天然五聚体(pentamer)抗体,具有10个重链和10个轻链。
IgM特性为半抗原的,这意味着在结合抗原时需要至少两个IgM分子。
6. 抗体定向抗体定向是指抗体分子相对于悬浮颗粒或细胞的方向。
当抗体结合特异性抗原时,会形成一个非常稳定的结构。
这种结构是通过碳氢键和氢键构成的。
抗体的定向使其可以识别和结合特定的抗原。
综上所述,抗体是一种非常复杂、多样化且重要的物质。
随着生命科学的不断发展,对于抗体的了解和研究也将会更加深入。
对于人类健康、疾病治疗以及医学的发展,抗体的应用前景也十分广泛且重要。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
免疫球蛋白目录1.2.3.4.1.2.3.4.5.6.5.1.2.3.4.6.1.2.3.7.1.2.8.1.2.3.4.5.9.1.2.3.10.11.1.2.12.具有活性的蛋白称为,又称为抗体。
是由机体的在的下分化、分裂而成的一组特殊球蛋白。
人和动物的中的免疫球蛋白极不均一,其组成、、大小、电荷、活性等都有很大差异,约占机体全部血的20~25%。
目前已在人、小鼠等血清中先后分纯得到5类免疫球蛋白,1968年,统一命名为免疫()、()、免疫球蛋白A()、()、()。
Porter等对血清IgG抗体的研究证明,单体分子的基本结构是由四条肽链组成的。
即由二条相同的分子量较小的肽链称为轻链和二条相同的分子量较大的肽链称为重链组成的。
轻链与重链是由二硫键连接形成一个四肽链分子称为Ig分子的,是构成免疫球蛋白分子的基本结构。
Ig单体中四条肽链两端游离的或的方向是一致的,分别命名为氨基端(N端)和羧基端(C端)。
图2-3 免疫球蛋白分子的基本结构示意图轻链和重链由于蛋白(M蛋白)是均一性球蛋白分子,并证明本周蛋白(BJ)是Ig分子的L链,很容易从患者和中纯化这种蛋白,并可对来自不同患者的进行,从而为Ig分子序列分析提供了良好的材料。
1.轻链(lightchain,L)轻链大约由214个氨基酸残基组成,通常不含碳水,分子量约为24kD。
每条轻链含有两个链内二硫键所组成的环肽。
L链共有两型:kappa(κ)与lambda(λ),同一个天然Ig分子上L链的型总是相同的。
正常人血清中的κ:λ约为2:1。
2.重链(heavychain,H链)重链大小约为轻链的2倍,含450~550个氨基酸残基,分子量约为55或75kD。
每条H链含有4~5个链内二硫键所组成的环肽。
不同的H链由于氨基酸的排列顺序、二硫键的数目和们置、含糖的种类和数量不同,其抗原性也不相同,根据H链抗原性的差异可将其分为5类:μ链、γ链、α链、δ链和ε链,不同H链与L链(κ或λ链)组成完整Ig的分子分别称之为IgM、IgG、IgA、IgD和IgE。
γ、α和δ链有4个环肽,μ和ε链含有5个环肽。
重链(heavy chain,H 链)由450~570个氨基酸残基组成,分子量约为50~70kD。
不同的H链因氨基酸的排列顺序、二硫键的数目和位置、含糖的种类和数量不同,其抗原性也不相同,可将其分为μ链、γ链、α链、δ链、ε链五类,这些H链与L链(κ链或λ链)组成的完整Ig分子分别称为IgM(μ)、IgG(γ)、IgA(α)、IgD(δ)和IgE(ε可变区和恒定区通过对不同骨髓蛋白或本周蛋白H链或L链的氨基酸序列比较分析,发现其氨基端(N-末端)氨基酸序列变化很大,称此区为可变区(V),而羧基末端(C-末端)则相对,变化很小,称此区为恒定区(C区)。
1.可变区(variableregion,V区)位于L链靠近N端的1/2(约含108~111个氨基酸残基)和H链靠近N端的1/5或1/4(约含118个氨基酸残基)。
每个V区中均有一个由链内二硫键连接形成的肽环,每个肽环约含67~75个氨基酸残基。
V区氨基酸的组成和排列随抗体结合抗原的特异性不同有较大的。
由于V区中氨基酸的种类、排列顺序千变万化,故可形成许多种具有不同结合抗原特异性的抗体。
L链和H链的V区分别称为VL和VH。
在VL和VH中某些局部区域的氨基酸组成和排列顺序具有更高的变休程度,这些区域称为高变区(hypervariable region,HVR)。
在V区中非HVR部位的氨基酸组面和排列相对比较保守,称为骨架区(framework region)。
VL中的高变区有三个,通常分别位于第24~34、50~65、95~102位氨基酸。
VL和VH的这三个HVR分别称为HVR1、HVR2和HVR3。
经X线衍射的研究分析证明,高变区确实为抗体与抗原结合的位置,因而称为决定簇互补区(complementarity-determining region,CDR)。
VL和VH的HVR1、HVR2和HVR3又可分别称为CDR1、CDR2和CDR3,一般的CDR3具有更高的高变程度。
高变区也是Ig分子独特型决定簇(idiotypic determinants)主要存在的部位。
在大多数情况下H链在与抗原结合中起更重要的。
图2-4 与抗原表位结合高变区(HVR)示意图(G表示相对保守的)2.恒定区(constantregion,C区)位于L链靠近C端的1/2(约含105个氨基酸残基)和H链靠近C端的3/4区域或4/5区域(约从119位氨基酸至C末端)。
H链每个区约含110多个氨基酸残基,含有一个由二锍键连接的50~60个氨基酸残基组成的肽环。
这个区域氨基酸的组成和排列在同一种属动物Ig同型L链和同一类H链比较恒定,如人抗IgG与人抗外的IgG,它们的V区不相同,只能与相应的抗原特异性的结合,但其C区的结构是相同的,即具有相同的抗原性,应用马抗人IgG第二体(或称)均能与这两种抗不同外毒素的抗体(IgG)发生结合。
这是制备第二抗体,应用、酶、同位毒等标记抗体的重要基础。
功能区Ig分子的H链与L链可通过链内二硫键折叠成若干球形功能区,每一功能区(domain)约由110个氨基酸组成。
在功能区中氨基酸序列有高度性。
1.L链功能区分为L链可变区(VL)和L链恒定区()两功能区。
2.H链功能区IgG、IgA和IgD的H链各有一个可变区(VH)和三个恒定区(CH1、CH2和CH3)共四个功能区。
IgM和IgE的H链各有一个可变区(VH)和四个恒定区(CH1、CH2、CH3和CH4)共五个功能区。
如要表示某一类免疫蛋白H链恒定区,可在C(表示恒定区)后加上相应重链名称(希腊字母)和恒定区的位置(阿拉伯数字),例如IgG重链CH1、CH2和CH3可分别用Cγ1、Cγ2和Cγ3来表示。
IgL链和H链中V区或C区每个功能区各形成一个免疫球蛋白折叠(immunoglobulin fold,Ig fold),每个Ig折叠含有两个大致平行、由二硫连接的β片层结构(betapleated sheets),每个β片层结构由3至5股反平行的链组成。
可变区中的高变区在Ig折叠的一侧形成高变区环(hypervariable loops),是与抗原结合的位置。
3.功能区的作用(1)VL和VH是与抗原结合的部位,其中HVR(CDR)是V区中与(或表位)互补结合的部位。
VH和VL通过非共价,组成一个FV区。
单位Ig 分子具有2个抗原结合位点(antigen-bindingsite),二聚体分泌型IgA 具有4个抗原结合位点,五聚体IgM可有10个抗原结合位点。
(2)CL和CH上具有部分同种异型的标记。
(3)CH2:IgGCH具有Clq结合点,能活化补体的经典活化途径。
IgG 借助CH2部分可通过主动到胎体内。
(4)CH3:IgGCH3具有结合、巨噬、粒细胞、和Fc段的功能。
IgMCH3(或CH3因部分CH4)具有补体结合位点。
IgE的Cε2和Cε3功能区与结合和FCεRI有关。
4.铰链区(hingeregion)铰链区不是一个的功能区,但它与其客观存在功能区有关。
铰链区位于CH1和CH2之间。
不同H铰链区含氨基酸数目不等,α1、α2、γ1、γ2和γ4链的铰链区较短,只有10多个氨基酸残基;γ3和δ链的铰链区较长,约含60多个氨基酸残基,其中γ3铰链区含有14个残基。
铰链区包括H链间二硫键,该区富含,不形成,易发生伸展及一定程度的转动,当VL、VH与抗原结合时此氏发生扭曲,使抗体分子上两个抗原结合点更好地与两个抗原决定簇发生互补。
由于CH2和CH3构型变化,显示出活化补体、结合细胞等生物学活性。
铰链区对、敏感,当用这些蛋白酶水解免疫球蛋白分子时常此区发生。
IgM和IgE 缺乏铰链区。
J链和分泌成分1.J链(joining chain)存在于二聚体分泌型IgA和五聚体IgM中。
J链分子量约为15kD,由于124个氨基酸组成的酸性糖蛋白,含有8个半残基,通过二硫键连接到μ链或α链的羧基端的半胱氨酸。
J链可能在Ig二聚体、五聚体或多聚体的组成以及在体内转运中的具有一定的作用。
2.分泌成分(secretorycomponent,SC)又称分泌片(secretory piece),是分泌型IgA上的一个辅助成分,分子量约为75kD,糖蛋白,由上皮细成,以共价形式结合到Ig分子,并一起被分泌到粘膜表面。
SC的存在对于抵抗液中蛋解酶的降解具有重要作用。
单体、双体和五聚体1.单体由一对L链和一对H链组成的基本结构,如IgG、IgD、IgE 血清型IgA。
2.双体由J链连接的两个单体,如分泌型IgA(secretory IgA,SIgA)二聚体(或多聚体)IgA结合抗原的亲合力(avidity)要比单体IgA高。
图2-5 分泌型IgA结构示意图3.五聚体由J链和二硫键连接五个单体,如IgM。
μ链Cys414(Cμ3)和Cys575(C端的尾部)对于IgM的多聚化极为重要。
在J链存在下,通过两个邻近单体IgMμ链Cys之间以及J链与邻μ链Cys575之间形成二硫键组成五聚体。
由粘膜下所合成和分泌的IgM五聚体,与粘膜上皮细胞表面pIgR(poly-Ig receptor,pIgR)结合,穿过粘膜上皮细胞到粘膜表面成为分泌型IgM(secretory IgM)。
酶解片段1.本瓜蛋白酶的水解片段Porter等最早用木瓜蛋白酶(papain)水解兔IgG,从而区划获知了Ig四肽链的基本结构和功能。
(1)裂解部位:IgG铰链区H链链间二硫键近N端侧切断。
(2)裂解片段:共裂解为三个片段:①两个Fab段(抗原结合段,fragmentof antigen binding),每个Fab段由一条完整的L链和一条约为1/2的H链组成,Fab段分子量为54kD。
一个完整的Fab段可与抗原结合,表现为单价,但不能形成凝集或。
Fab中约1/2H链部分称为Fd段,约含225个氨基酸残基,包括VH、CH1和部分铰链区。
②一个Fc段(可结晶段,fragmentcrystallizable),由连接H链二硫键和近羧基端两条约1/2的H链所组成,分子量约50kD。
Ig在异种间免疫所具有的抗原性主要存在于Fc段。
图2-6 人分泌型IgA和分泌型IgM的局部产生示意图图2-7 IgM结构示意图2.胃蛋白酶的水解片段Nisonoff等最早用胃蛋白酶(pepsin)裂解免疫球蛋白。
(1)裂解部位:铰链区H链链间二硫键近C端切断。
(2)裂解片段:1)F(ab')2:包括一对完整的L链和由链间二硫键相连一对略大于Fab中Fd的H链,称为Fd',约含235个氨基酸残基,包括VH、VH1和铰链区。