反转录转座子PPT课件
合集下载
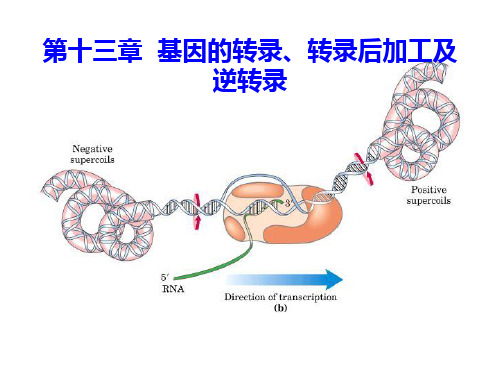
第十三章基因的转录、转录后加工及逆转录ppt课件
◆真核生物启动子 (1)DNA序列在转录起始点的5’端区(上游 区)(2)-25bp :TATA盒(Hogness box) (3)-90bp :GC盒 (4)-70bp :CAAT盒
-90
-70
GC
CAAT
RNA聚合酶Ⅱ催化的转录起始
RNA聚合酶Ⅱ催化各种前体mRNA的合 成
需要多种TF参与:TFⅡA-J
第一节 参与转录的酶
RNA聚合酶——依赖DNA的RNA聚合酶 (DNA-dependent RNA polymerase,DDRP)
以DNA为模板,催化2个游离的NTP 形成3’,5’-磷酸二酯键
一、原核生物RNA聚合酶
1、大肠埃希菌RNA聚合酶的组成 (1)全酶(holoenzyme)
由4种(5个)亚基α2ββ’σ组成 (2)核心酶(core enzyme)
作用位置
步骤1 步骤2
200~250
(3)真核生物mRNA转录后加工—剪接
内含子
外显子
DNA
hnRNA
●剪接所需条件
snRNA (U1-U6) + 蛋白质 (核内小分子核酸)
多种snRNP (核内小核蛋白颗粒)
多种snRNPs装配成
剪接体 (参与剪接过程)
4、RNA编辑(RNA editing)
二.真核生物RNA转录后的加工 1、rRNA转录后的加工
真核生物rRNA 的基因
(rDNA)
转录产物
成簇纵列串联排列
高度重复序列DNA
核质:(Ⅲ)--不需加 工
5s rRNA
核仁:(Ⅰ)--加工
5.8s rRNA 28s rRNA 18s rRNA
rDNA 内含子
基因间隔
转座因子介绍

宿主 DNA
过量的 OUT-RNA 与其配对,抑制 IN-RNA 的转录
甲基化阻止转录酶和 DNA 结合
甲基化阻止转录酶合成 图 23- 46 几种抑制 Tn10 专座的机制,主要是通过控专座酶的合成来调节
主要类型转座子
类型
细菌插入序 列(IS)
结构特征
TIRs;转座酶和或拆 分酶编码基因 IS组份;抗药基因 TIRs;蛋白质编码 基因(有内含子)
表 复合转座子的结构和功能
转座因 子 Tn903 Tn9 Tn10 长 度 (bp) 3100 2500 9300 遗传标 记 KanR CamR TetR 末端组 件 IS903 IS 1 IS 10R IS 10L Tn5 5700 KanR IS 50R IS 50L 反向 1 bp的改变 反向 有2.5%的 差异 方向 反向 正向 二组件的关 系 相同 推测相同 组件的功能 二者皆有功 能 预计有功能 有功能 无功能 有功能 无功能
① 如IS1因子不论以什么方向插入都会降低基因的表达,
②而IS2因子以同一方向插入到染色体中,则会减少基因的表 达,但以相反方向整合,则会增加基因表达。 ③不同转座元件的转座频率不同。一般为10-3-10-4 次/世代。 ④恢复频率则很低(通过精确切除IS元件而产生),为10-6-10-10/ 代。比插入频率率低 10 -3 倍。
• 3.复合转座子在两个末端有IS序列 • 4.首先,在靶位点处产生一个交错切口,切 出转座子。接着,转座子与靶位点连接。 最后,填补插入位点两侧的单链区。 • 5.转座子转座时能够导致宿主序列的缺失、 重复或插入。另外,转座子通过宿主重组 系统导致基因组重排。
2 复合转座子(composite
transposon)
反转录转座子

R U5
gag
缺陷型病毒 v-onc 翻译
U3
R
R U5
gag
助病毒 pol 翻译
env
U3
R
助病毒的蛋 白能使缺陷 型 图 23-65 助病毒的产物可供缺陷型病毒复制 (仿 B.Lewin:《GENES》 Ⅵ,1997, Fig.19.9)
前病毒
c-onc 基因
前病毒 LTR 外显子
LTR gag pol env LTR 外显子 内含子 外显子 LTR gag pol env
反转录病毒 整合入宿主 DNA中的分 子机制,其 本质是转座
整合酶在 LTR 的 3’端 产生 2b 的缺口
整合酶在靶 DNA 上进行交错切割
5’ 3’ 3’ 3’
3 ’5 ’
3’ 5’3’ 5’
整合酶将 LTR 缺口的 3’端和靶 DNA 交错切口的 5’端连接起
图 23-64 整合酶催化反转录病毒的 DNA 整合到宿主的基因组中。
艾 兹 病 病 毒
艾滋病毒的基因结构
vpr rev gag vif tat vpu tat rev nef
LTR
pol
env
LTR
LTR 长末端重复序列 gag 核心蛋白,反转录酶 pol 蛋白酶 vif 感染因子 vpr,vpu 复制因子 tat 反式激活因子 rev(art抗阻遏翻译基因活化, trs trans regulator of splicing ) 调节因子 env 衣壳蛋白 nef(3’orf) negative factor 抑制复制模板
P ♂×M♀ 雌性染色体
P 细胞型
87K
杂种不育 M ♂×P♀ 雌性染色体 P 因子
ORF0 ORF1 ORF2 ORF3
第四章DNA的转座分子生物学2

Ds因子长0.4-4kb,它的中间(在转座酶基因中)有许多种长度不等的缺失, 如Ds9只缺失194bp,而Ds6则缺失2.5kb,Ds的两端也都有11bp的反向重复 序列。
Ac和Ds的末端反向重复几乎是一样的,只有一个不同之处:Ac两端最外边的核苷酸
是彼此不互补的T:G,而Ds是互补的T:A(图)。
(免疫性)
e) 靶序列在转座因子两侧会形成正向重复
f) 转座因子的切除与转座将产生复杂的遗传学效应
当前第31页\共有60页\编于星期六\8点
三、转座子转座频率的调控
♥ 每个转座子控制自身转座的核心----控制转座酶的水平
不到一个转座酶分子/世代/细胞
♥ 自发转座频率---10-7
1)、Tn10转座机制
当前第29页\共有60页\编于星期六\8点
当前第30页\共有60页\编于星期六\8点
● 转座的特点
a) 不依赖供体序列与靶位点间序列的同源性
b) 转座不是简单的转移,涉及转座子的复制
c) 转座插入的靶位点并非完全随机(插入专一型)
Hotspots (热点)
Regional preference ( 在3kb区域内的随机插入) d) 某些转座因子(Tn3)对同类转座因子的插入具有排他性
当前第35页\共有60页\编于星期六\8点
指减低蛋白质合成速度的基因突变
当前第36页\共有60页\编于星期六\8点
2. 造成插入位点靶DNA的少量碱基对重复
IS1、Tn10: 造成9bp的重复。 IS3: 造成3或4bp的重复。 IS4: 造成11bp的重复。
3. 插入位点出现新基因 复合转座子带有抗性基因(如抗药性基因ampc),可
♥ POUT:强启动子
向右转录宿主DNA ♥ INRNA和OUTRNA
Ac和Ds的末端反向重复几乎是一样的,只有一个不同之处:Ac两端最外边的核苷酸
是彼此不互补的T:G,而Ds是互补的T:A(图)。
(免疫性)
e) 靶序列在转座因子两侧会形成正向重复
f) 转座因子的切除与转座将产生复杂的遗传学效应
当前第31页\共有60页\编于星期六\8点
三、转座子转座频率的调控
♥ 每个转座子控制自身转座的核心----控制转座酶的水平
不到一个转座酶分子/世代/细胞
♥ 自发转座频率---10-7
1)、Tn10转座机制
当前第29页\共有60页\编于星期六\8点
当前第30页\共有60页\编于星期六\8点
● 转座的特点
a) 不依赖供体序列与靶位点间序列的同源性
b) 转座不是简单的转移,涉及转座子的复制
c) 转座插入的靶位点并非完全随机(插入专一型)
Hotspots (热点)
Regional preference ( 在3kb区域内的随机插入) d) 某些转座因子(Tn3)对同类转座因子的插入具有排他性
当前第35页\共有60页\编于星期六\8点
指减低蛋白质合成速度的基因突变
当前第36页\共有60页\编于星期六\8点
2. 造成插入位点靶DNA的少量碱基对重复
IS1、Tn10: 造成9bp的重复。 IS3: 造成3或4bp的重复。 IS4: 造成11bp的重复。
3. 插入位点出现新基因 复合转座子带有抗性基因(如抗药性基因ampc),可
♥ POUT:强启动子
向右转录宿主DNA ♥ INRNA和OUTRNA
第十三章 转座子PPT课件

进一步的断裂和融合循环,造成斑驳效应。
cl cl cl
Ds provides a site to initiate the chromatid fusion-bridge-breakage cycle. The products can be followed by clonal analysis.
结构特征 和 转座特点
插入序列的结构特征
(1)含短的末端反向重复序列; (2)含编码转座酶的基因; (3)靶位点存在 5-9 bp 的短正向重复序列。
In this example, the target is a 5 bp sequence. The ends of the transposon consist of inverted repeats of 9 bp.
• 转座子多个拷贝之间的同源重组引起宿 主DNA的重排;
• 转座子重复序列间的同源重组导致精确 或不精确切除。
A major cause of sequence change within a genome is the movement of a transposon to a new site. This may have direct consequences on gene expression. Unequal crossing-over between related sequences causes rearrangements. Copies of transposons can provide targets for such events.
◆可以作为一个整体进行转座; ◆含有的一个或两个I座机制
所有转座子的共同机制: 在靶DNA上造成交错切口,转座子 与突出的末端相连,填补缺口。
cl cl cl
Ds provides a site to initiate the chromatid fusion-bridge-breakage cycle. The products can be followed by clonal analysis.
结构特征 和 转座特点
插入序列的结构特征
(1)含短的末端反向重复序列; (2)含编码转座酶的基因; (3)靶位点存在 5-9 bp 的短正向重复序列。
In this example, the target is a 5 bp sequence. The ends of the transposon consist of inverted repeats of 9 bp.
• 转座子多个拷贝之间的同源重组引起宿 主DNA的重排;
• 转座子重复序列间的同源重组导致精确 或不精确切除。
A major cause of sequence change within a genome is the movement of a transposon to a new site. This may have direct consequences on gene expression. Unequal crossing-over between related sequences causes rearrangements. Copies of transposons can provide targets for such events.
◆可以作为一个整体进行转座; ◆含有的一个或两个I座机制
所有转座子的共同机制: 在靶DNA上造成交错切口,转座子 与突出的末端相连,填补缺口。
DNA的转座(DNA transposition)【精选-PPT】

2)特点: ♣ 不必借助同源序列就可移动的DNA片段,即转座作
用与供体和受体之间的序列无关。 ♣ 原核生物和真核生物均有转座子。 ♣ 转座序列可沿染色体移动,甚至在不同染色体间跳
跃。
3)种类与特点
(1) 两种类型: A 简单转座子(simple transposon) 或(插入序列 insertion sequence IS ) B 复合转座子(composite transposon)
转座酶
38bp regulator β- 内酰胺酶
b)两端重复序列为IS的复合转座子
e.g. IS插入到功能基因两端,可能形成复合转座因子
其两翼往往是两个相同或高度同源的IS序列, 表明IS序列插入到某个功能基因两端时就可能产生 复合转座子。一旦形成复合转座子,IS序列就不能 再单独移动,因为它们的功能被修饰了,只能作为 复合体移动。
Ac-Ds转座元件结构示意图。右边示Ac及Ds元件的单链DNA末端反向重复 配对所形成的茎环结构,这种结构可能对转座有意义
由于缺失转座酶,Ds因子不能自主移动,因此Ds因 子是非自主移动的受体因子(dissociator),而Ac则为自主 移动的调节因子(activator ),Ds的转座依赖于Ac元件的存 在。
IS
IS
IS
transposition
IS L
IS R
臂
中心区
臂
♣ 两侧的IS既可以是 IR,又可以是DR状态 (IR多)
♣ 当两个IS组件相同 时,其中任一个都可 行使转座功能
♣ 不同时,主要依 靠一个
2、转座噬菌体 Mu phage (巨型转座子 ) 以E.coli为寄主的温和型噬菌体(溶源、裂解)
b) 进入裂解生长后, 复制产生后代Mu DNA 几乎全部插入寄主DNA 中,并可继续转座(形 成寄主DNA和Mu的共 合体),噬菌体成熟时, 切段共合体包装
反转录的原理及应用PPT

VS
反转录还可以用于检测基因变异和突 变。通过将特定DNA片段反转录为 cDNA并进行测序,可以检测基因序 列的差异,为遗传病和癌症等疾病的 诊断和治疗提供依据。
基因组学和生物信息学
在基因组学和生物信息学领域,反转录也发挥着重要作用。通过反转录和扩增特定基因组区域,可以 生成足够量的DNA进行高通量测序,推动全基因组测序和表观遗传学研究。
的特异性。
反转录的前景
广泛应用
反转录技术是基因表达研究、基因克隆、基因治疗等领域的重要手 段,随着相关领域的发展,反转录技术的应用将更加广泛。
新技术发展
随着测序技术的发展,单细胞测序、纳米孔测序等新技术为反转录 提供了新的应用场景和研究方向。
疾病诊断和治疗
反转录技术在疾病诊断和治疗方面具有重要价值,如针对某些遗传性 疾病、肿瘤等疾病的基因治疗。
要点二
详细描述
反转录酶链反应的基本步骤包括RNA提取、逆转录、链式反 应扩增和产物检测。在逆转录步骤中,反转录酶催化RNA模 板逆转录为cDNA,这一过程中需要使用特异性引物来确保 逆转录的特异性。链式反应扩增后,可以通过凝胶电泳或荧 光检测等方法对产物进行检测。与传统的反转录PCR相比, 反转录酶链反应具有更高的灵敏度和特异性,能够更好地应 用于临床诊断和科学研究。
详细描述
实时反转录PCR使用荧光标记的探针或染料, 在扩增过程中能够实时监测荧光信号。通过 荧光信号的变化,可以实时跟踪反应过程, 获取扩增的实时数据。与传统的反转录PCR 相比,实时反转录PCR具有更高的灵敏度和 特异性,能够更好地应用于临床诊断和科学 研究。
反转录酶链反应
要点一
总结词
反转录酶链反应是一种将RNA逆转录为cDNA并进行链式反 应的实验技术,通过使用反转录酶和特异性引物,将RNA模 板逆转录为DNA,再进行链式反应扩增。
DNA的重组与转座PPT课件

DNA的重组与转座
7
➢细菌的接合作用(Bacterial Conjugation) Sex pillus
Extra genetic material
Genome of the recipient
F-factor for cleavage the circular DNA
DNA的重组与转座
8
▪ Transformation
Ruv A (DNA 重组与修复蛋 白)
RuvB
rec F
与双链DNA和单链DNA结合
rec O
促进互补的单链DNA的复性
rec R
参与DNA的修复与重组
recB, rec C, 1.依赖于ATP的外切酶的活性;
rec D
2.可被ATP增强的内切酶的活性;
3. ATP依赖的解旋酶的活性。
ruv A
DNA解旋酶。特异的作用于Holliday衔接点,并与RuvB 相互作用,促进Ruv A 及RuvB沿着DNA链移动及双链 DNA的解旋。
DNA的重组与转座
26
➢转座引起的遗传学效应
1.引起插入突变 2.在插入位置染色体DNA重排而出现新基因 3.影响插入邻近基因的表达,使宿主表型改变 4.转座子插入染色体后引起两侧染色体畸形
DNA的重组与转座
27
➢真核生物的转座因子(The Transposable element in Eukaryote)
DNA的重组与转座
1
同源重组(Homologous recombination)
同源重组:在有相同或近于相同碱基序列(75bp以上) 的DNA分子之间的遗传交换,发生在减数分裂过程。
同源重组具备的条件: 1)两条同源双链DNA分子紧密结合; 2)两个DNA链上同一个部位的单链断裂
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
P 因子
ORF0 ORF1 ORF2 ORF3
杂种不育
P 细胞型 66KD 阻遏物
66K
阻遏物 抑制所
有P因 子转座
图 23-57 杂种不育取决于基因组中 P 因子和不同类型细胞中 66KD 阻遏物的相互作用。 (仿 B.Lewin:《GENES 》Ⅵ,1997, Fig.18.27)
第八节 反转录病毒和反转录
splicing )
SUCCESS
THANK YOU
2019/8/22
10-80 80-100 终止密码子
170-1350
R U5 gag
~2000
pol
env
~2900 ~1800
转录
U3 R
雄性染色体 P 因子
ORF0 ORF1 ORF2 ORF3
P 品系 (P♂×P♀) 雌性染色体 P 因子
ORF0 ORF1 ORF2 ORF3
P♂×M♀ 雌性染色体
P 细胞型 66KD 阻遏物
66K
阻遏物
抑制所 有P因 子转 座
P 细胞型
P因子 合成转 座酶
87K
雄性染色体 无 P 因子
M♂×P♀ 雌性染色体
缺陷型的转座因子的插入而产生也就顺理
成章了(缺陷的转座因子自己并不能移
动)。Dt的缺乏使得表型保持稳定。
Phoades将基因 型a1/a1;Dt/的玉米发芽, 检测它们是否 带有在各组织 中产生色素斑 的基因
叶片的花 斑表型
CACTACCAAGAAAA 1 ORF1
TTTTCTTGTAGTG 2 3 4 5 6 7 8 9 10 11 ORF2
测交, 结果有的后代完全是有颜色的。表明在亲本中每
个斑点实际上是回复突变的。
这样a1成为首次发现的不稳定突变等
位 基 因 ( unstable mutant allele ) 的 例
子。即一种回复突变率很高的等位基因。
然而这种等位基因的不稳定是取决于不连
锁的Dt基因的存在。一旦回复突变的发生,
它们就变得稳定了;即Dt基因能离开A1基因, 这时A1表型不再改变。这样a1表型是由一个
2. 顺式优先(Cis-Preference)
3dam甲基化
减少转座频率 把转座和复制叉的途径连系起来。
转座酶
Tn10
>>>>>>> IS10R >>>>>>>> 宿主 DNA
OUT IN
过量的 OUT-RNA 与其配对,抑制 甲基化阻止转录酶和 DNA 结合 IN-RNA 的转录
甲基化阻止转录酶合成
艾 兹 病 病 毒
艾滋病毒的基因结构
vpr
rev
rev
tat vpu
gag tat
vif nef
LTR env
pol LTR
LTR
长末端重复序列
gag
核心蛋白,反转录酶
pol
蛋白酶
vif
感染因子
vpr,vpu
复制因子
tat
反式激活因子
rev(art抗阻遏翻译基因活化, trs trans regulator of
二.果蝇中的P因子
“ 杂种不育”(hybrid dysgenesis)。 P型(父本贡献的, paternal contributing) M型(母本贡献的, maternal contributing) M(♂)×P(♀)
后代不育 P(♂)×M(♀)
后代可育。
雄性染色体 P 因子
ORF0 ORF1 ORF2 ORF3
A1a1Dtd原t的来植的物品,系自可交能产为生A1了A1上dt述dt比,例突。变后产生
产生斑点的一种可能是在体细胞中产生了回 复突变a1→A1,但大量的斑点需要很高频率的回复
突变。Rhoades能在a1a1Dt_(花斑)特殊无性种植
物的花中找到相应的花药,其花粉应携带回复突变
产生色素的基因型,而他用这些花粉与a1a1的植株
有的Tn; (2) 在新位置上转座子的两端出现正向重复靶
序列; (3)转座过程中出现共合体。
六.非复制型转座 七.Tn10的转座具有多项控制 1.“ 多 拷 贝 抑 制 ” ( multicopy
inhibition)
(1)Pout 比 Pin强得多; (2) OUT RNA比IN RNA较为稳定。 (3) OUT RNA的功能是作为一种反义RNA,
McClintock。1940~1950描述了大量的控制因 子
A1 A1 dt dt
双突变
A1 a1 Dt dt
自交
9/16 3/16
3/16
1/16
A1-D1- A1- dt dt
a1a1Dt-
a1a1d1d1
A1: 控制色素形 成
Dt: 点
控制产图生2斑3-47 玉米花斑表型的遗传学解释。
Marcus Rhoades分析了一种墨西哥玉米 的穗,此穗来自于一种籽粒有颜色的纯种自花受粉 的玉米,但它在后代中表现出一种意外的双因子杂 种修饰性孟德尔分离比
转座酶结合在 Tn 两端 转座子末端被交错剪切
+
另一条链也被切
受体也被交错切割
图 23-42 交换结构经剪切释放 后导致非复制型转座子插入到靶 DNA 中,DR 包在两侧,供体留 下了一个双链缺口。
供体被释放
Tn 连接到靶上
图 23-43 Tn 的两条链先后被切割,然后转座子与切开 的靶 复制性转座在转座后原来的位置上保留原
图 23- 46 几种抑制 Tn10 专座的机制,主要是通过控专座酶的合成来调节
第七节 真核生物的转座因子
一、玉米中的控制因子
1938年Marcus Rhoades首次发现不稳定突变
等位基因(unstable mutant allele),即 一种回复突变率很高的等位基因。不稳
定是取决于不连锁的Dt基因的存在。
子
一.反转录病毒(retroviruses)
(一) 反转录病毒的生活史
反转录子(retroposons)
反转录转座子(retrotransposons)
1. 反转录病毒基因组的结构与功能
2. 反转录病毒的整合模型
3. 反转录病毒可转录细胞的序列
二.酵母的Ty因子(transponson yeast)
转录
Spm-w-8011 dSpm-7995 dSpm-799 dSpm-8004
图 23-54 Spm/En 有两个基因,tnpA 有 11 个外显子转录成 2500b 拼接 mRNA。tnpB 含有 6kbmRNA,含 2 个读框。(仿 B.Lewin:《GENES
Ⅵ,1997, Fig.18.24)
ORF0 ORF1 ORF2 ORF3
杂种不育
P 细胞型 66KD 阻遏物
66K
阻遏物 抑制所
有P因 子转座
图 23-57 杂种不育取决于基因组中 P 因子和不同类型细胞中 66KD 阻遏物的相互作用。 (仿 B.Lewin:《GENES 》Ⅵ,1997, Fig.18.27)
第八节 反转录病毒和反转录
splicing )
SUCCESS
THANK YOU
2019/8/22
10-80 80-100 终止密码子
170-1350
R U5 gag
~2000
pol
env
~2900 ~1800
转录
U3 R
雄性染色体 P 因子
ORF0 ORF1 ORF2 ORF3
P 品系 (P♂×P♀) 雌性染色体 P 因子
ORF0 ORF1 ORF2 ORF3
P♂×M♀ 雌性染色体
P 细胞型 66KD 阻遏物
66K
阻遏物
抑制所 有P因 子转 座
P 细胞型
P因子 合成转 座酶
87K
雄性染色体 无 P 因子
M♂×P♀ 雌性染色体
缺陷型的转座因子的插入而产生也就顺理
成章了(缺陷的转座因子自己并不能移
动)。Dt的缺乏使得表型保持稳定。
Phoades将基因 型a1/a1;Dt/的玉米发芽, 检测它们是否 带有在各组织 中产生色素斑 的基因
叶片的花 斑表型
CACTACCAAGAAAA 1 ORF1
TTTTCTTGTAGTG 2 3 4 5 6 7 8 9 10 11 ORF2
测交, 结果有的后代完全是有颜色的。表明在亲本中每
个斑点实际上是回复突变的。
这样a1成为首次发现的不稳定突变等
位 基 因 ( unstable mutant allele ) 的 例
子。即一种回复突变率很高的等位基因。
然而这种等位基因的不稳定是取决于不连
锁的Dt基因的存在。一旦回复突变的发生,
它们就变得稳定了;即Dt基因能离开A1基因, 这时A1表型不再改变。这样a1表型是由一个
2. 顺式优先(Cis-Preference)
3dam甲基化
减少转座频率 把转座和复制叉的途径连系起来。
转座酶
Tn10
>>>>>>> IS10R >>>>>>>> 宿主 DNA
OUT IN
过量的 OUT-RNA 与其配对,抑制 甲基化阻止转录酶和 DNA 结合 IN-RNA 的转录
甲基化阻止转录酶合成
艾 兹 病 病 毒
艾滋病毒的基因结构
vpr
rev
rev
tat vpu
gag tat
vif nef
LTR env
pol LTR
LTR
长末端重复序列
gag
核心蛋白,反转录酶
pol
蛋白酶
vif
感染因子
vpr,vpu
复制因子
tat
反式激活因子
rev(art抗阻遏翻译基因活化, trs trans regulator of
二.果蝇中的P因子
“ 杂种不育”(hybrid dysgenesis)。 P型(父本贡献的, paternal contributing) M型(母本贡献的, maternal contributing) M(♂)×P(♀)
后代不育 P(♂)×M(♀)
后代可育。
雄性染色体 P 因子
ORF0 ORF1 ORF2 ORF3
A1a1Dtd原t的来植的物品,系自可交能产为生A1了A1上dt述dt比,例突。变后产生
产生斑点的一种可能是在体细胞中产生了回 复突变a1→A1,但大量的斑点需要很高频率的回复
突变。Rhoades能在a1a1Dt_(花斑)特殊无性种植
物的花中找到相应的花药,其花粉应携带回复突变
产生色素的基因型,而他用这些花粉与a1a1的植株
有的Tn; (2) 在新位置上转座子的两端出现正向重复靶
序列; (3)转座过程中出现共合体。
六.非复制型转座 七.Tn10的转座具有多项控制 1.“ 多 拷 贝 抑 制 ” ( multicopy
inhibition)
(1)Pout 比 Pin强得多; (2) OUT RNA比IN RNA较为稳定。 (3) OUT RNA的功能是作为一种反义RNA,
McClintock。1940~1950描述了大量的控制因 子
A1 A1 dt dt
双突变
A1 a1 Dt dt
自交
9/16 3/16
3/16
1/16
A1-D1- A1- dt dt
a1a1Dt-
a1a1d1d1
A1: 控制色素形 成
Dt: 点
控制产图生2斑3-47 玉米花斑表型的遗传学解释。
Marcus Rhoades分析了一种墨西哥玉米 的穗,此穗来自于一种籽粒有颜色的纯种自花受粉 的玉米,但它在后代中表现出一种意外的双因子杂 种修饰性孟德尔分离比
转座酶结合在 Tn 两端 转座子末端被交错剪切
+
另一条链也被切
受体也被交错切割
图 23-42 交换结构经剪切释放 后导致非复制型转座子插入到靶 DNA 中,DR 包在两侧,供体留 下了一个双链缺口。
供体被释放
Tn 连接到靶上
图 23-43 Tn 的两条链先后被切割,然后转座子与切开 的靶 复制性转座在转座后原来的位置上保留原
图 23- 46 几种抑制 Tn10 专座的机制,主要是通过控专座酶的合成来调节
第七节 真核生物的转座因子
一、玉米中的控制因子
1938年Marcus Rhoades首次发现不稳定突变
等位基因(unstable mutant allele),即 一种回复突变率很高的等位基因。不稳
定是取决于不连锁的Dt基因的存在。
子
一.反转录病毒(retroviruses)
(一) 反转录病毒的生活史
反转录子(retroposons)
反转录转座子(retrotransposons)
1. 反转录病毒基因组的结构与功能
2. 反转录病毒的整合模型
3. 反转录病毒可转录细胞的序列
二.酵母的Ty因子(transponson yeast)
转录
Spm-w-8011 dSpm-7995 dSpm-799 dSpm-8004
图 23-54 Spm/En 有两个基因,tnpA 有 11 个外显子转录成 2500b 拼接 mRNA。tnpB 含有 6kbmRNA,含 2 个读框。(仿 B.Lewin:《GENES
Ⅵ,1997, Fig.18.24)