5基于途径分析的L_异亮氨酸发酵溶氧控制研究
L-异亮氨酸发酵工艺条件的研究

L-异亮氨酸发酵工艺条件的研究
浦军平;陈刚;王开国
【期刊名称】《食品与发酵工业》
【年(卷),期】2003(029)002
【摘要】研究了L-异亮氨酸产生菌Apsm533128(Brevibacterium flavum)菌体的生长规律.讨论了葡萄糖、玉米浆的浓度对L-异亮氨酸的积累的影响,并且在30t 发酵罐上进行发酵工艺条件的研究,通过对通气量、温度、pH值等外环境发酵条件的控制,在30t发酵罐工业化大生产中,L-异亮氨酸发酵产酸率达2.3~2.5%.并对其发酵动力学进行了初步的研究.
【总页数】4页(P93-96)
【作者】浦军平;陈刚;王开国
【作者单位】江苏亚太氨基酸有限公司,张家港市,215600;江苏亚太氨基酸有限公司,张家港市,215600;江苏亚太氨基酸有限公司,张家港市,215600
【正文语种】中文
【中图分类】TQ92
【相关文献】
1.L-异亮氨酸发酵工艺条件的研究 [J],
2.基因敲除降低L-缬氨酸发酵中亮氨酸和异亮氨酸含量的研究 [J], 胡炎华;朱亚然;宫卫波
3.L-异亮氨酸生产菌发酵水平提高的研究 [J], 黄平
4.微生物发酵法生产L-异亮氨酸的研究进展 [J], 孔帅;方应浩;周航;许鹏飞;杨潇;任
立伟;龚大春
5.代谢工程改造谷氨酸棒状杆菌促进L-异亮氨酸发酵合成的研究进展 [J], 谭海;顾阳;卢南巡;常景玲;李志刚
因版权原因,仅展示原文概要,查看原文内容请购买。
L-异亮氨酸发酵的研究

L-异亮氨酸发酵的研究何家骏1材料及设备斜面培养基、摇瓶种子培养基(略)。
1.1一级种子培养基(%)葡萄糖6.5,碳酸钙5.0,玉米油0.4,硫酸铵2.5,硫酸亚铁0.004g/100ml,硫酸锰0.004g/100ml,磷酸二氢钾0.35,灭菌后pH值为6.5~6.8。
1.2二级发酵培养基(%)葡萄糖15,硫酸铵5.0,碳酸钙5.0,硫酸锰0.008g/100ml,硫酸亚铁0.004g/100ml,硫胺素240μg/100ml,生物素30μg/10ml灭菌后pH值6.0至6.5。
1.3设备摇瓶机2台,一级种子罐600L2台,补糖罐2000吨2台,油罐150L1台,发酵罐2000L2台,氨水计量罐200L1台。
721分光光度计1台,旋光计1台,分析天平1台,25型酸度计2台。
2种子罐代谢考察在无菌室内,取少许菌种接入摇瓶中,放在摇瓶机上,培养20~24h,并瓶,接入种子罐。
步骤如下:2.1罐温33~35℃。
2.2通气通入40~50℃无菌空气,流量用进气阀门调节,开始稍为小开,8h后全开。
又可以用进气与排气阀门调节,(因没装流量计)。
2.3培养周期种子罐培养周期20~24h。
2.3.1种子罐内菌体代谢变化(1)pH值培养24h后,pH稍有下降,然后再稍有回升。
(2)糖代谢培养过程中,糖逐渐消耗,24h稍有回升。
(3)氨基氮培养24h后,氨基氮稍有回升。
2.3.2菌体外观变化摇瓶培养液接入种子罐后进行培养,开始种子液为乳白色,培养20h,转乳黄色,24h后逐渐转为浅黄色。
即可移入发酵罐。
3发酵的代谢变化利用压差法,将种子液无菌移入发酵罐,发酵按下列条件掌握。
3.1罐温32~33℃。
3.2通气通入40~50℃无菌空气,用进、排气阀门根据菌种生长情况,随时调节。
3.3发酵周期44~48h。
3.4补糖培养12h后,开始第1次补糖,全发酵过程补7~8次糖,每次补入100L。
3.5pH值发酵过程中,用氨水调解,12h,开始通入氨水,每8h通入15~25L,培养30h后,氨水用量下降,36h后,停止通入氨水,L-异亮氨酸发酵代谢曲线如附图:附图L-异亮氨酸发酵代谢曲线4小结从代谢曲线可知,全程用氨水调解,掌握pH值与氨基氮变化,开始通入氨水时,发酵液pH值即上升,但很快被菌体吸收及利用,pH值迅速下降。
溶氧对发酵的影响及控制

溶氧对发酵的影响及其控制The dissolved oxygen concentration in the fermentation broth (Dissolved Oxygen, referred to as DO) is the key factor to influence the fermentation, has an important influence on microbial growth and product formation. According to the demand of dissolution characteristics and microbial oxygen on oxygen, analysis of the effects of dissolved oxygen on the fermentation and the effect on fermentation, and then determine the control of dissolved oxygen in the fermentation broth and transfer, the maximum production efficiency.Compared with normal PID controller, the new controller is of small overshoot and quick response, improved stability of the system andincrease the yield of products. Study the influence of dissolved oxygen and controlling the fermentation to improve production efficiency, improve product quality, etc. are important.溶氧浓度(DO)作为发酵控制中的一个关键参数,直接影响着发酵生产的稳定性和生产成本,受到工业生产和实验室研究的重视,无论是厌氧还是需氧发酵,研究发酵液中溶氧对发酵的影响都有重要意义。
产生L_异亮氨酸的黄色短杆菌的代谢途径分析

!"#$%"& ’(")&*+* ,-. !.-/01#+-( -, 234*-)501+(5 6& !"#$%&’()#"%*+ ,-’$*+
! " #$ %&’(!2) ! " #$ *+&,-+.(/!3) 01" #,21" #$ 2&,3&’!2) 456# #&(/%
!. 45657 8759:;<5=>? 2. @A66:B: AC D5A6AB> 27E 1B;5FG6=G;: H7B57::;57B? @I27BFIG7 !+""%%J 3. @I572$42K27 875A7 LA<K5=26? @I27BFIG7 !+""+%J %. @A66:B: AC D5A=:FI7A6AB>? M527N57 8759:;<5=> AC OF5:7F: 27E M:FI7A6AB>? M527N57 +""%%%? @I572
,$ 异亮氨酸是人体 - 种必需氨基酸之一,在临床领域 主 要
作为复合氨基酸输液、 三支链氨基酸输液、 氨基酸口服液等的成 分之一, 用于治疗肝病、 肝昏迷、 体弱乏力等疾 病 , 是比较昂贵的 氨基酸原料药之一。 目前国内 ,$异亮氨酸的市场需求急剧上升, 因而对微生物发酵法生产 ,$异亮氨酸的研究也日益加强 V!W。 工业微生物学家试图通过对细胞 进 行 遗 传 修 饰 来 改 变 生 物 过程的运行, 以实现目的产物的过量合成, 这就需要对代谢网络 进行理性的分析。代谢途径的确定对于 提 供 一 种 以 整 个 代 谢 功 能和表型为出发点进行途径定位的方法 是 必 不 可 少 的 。 途 径 分 是代谢工程中用以指导遗传操作的理论基 析( K2=IR2> 2726><5<) 础, 是代谢网络分析的基本方法 V%W。对于这种鉴别和分析的方法, 最有发展前途的是利用凸分析原理,这 些 原 理 包 括 最 近 提 出 的 元素模型概念和极端途径模型,两者都 能 用 于 确 定 代 谢 网 络 的 生产能力的界限。一些反应的不可逆性 使 得 代 谢 网 络 中 的 相 当 多的通量存在于一个凸圆锥中,该圆锥 可 由 用 矩 阵 方 法 计 算 出 来的向量确定。代谢途径的结构分析需 要 有 关 计 量 学 模 型 和 可 逆性或非可逆性反应的知识,然后通过 计 算 就 能 得 到 最 优 途 径 这已通过计算机程序得到实现 V’W。王健、 张蓓等对 的通量分布 V+$*W,
发酵工艺控制(溶氧)

(2)、降低发酵液中的CL
降低发酵液中的CL,可采取减少通气量或降低搅拌转速等方式来降低KLa,使发酵液中的CL降低。但是,发酵过程中发酵液中的CL不能低于C临界,否则就会影响微生物的呼吸。
目前发酵所采用的设备,其供氧能力已成为限制许多产物合成的主要因素之一,故此种方法亦不理想。
(一)影响氧传质推动力的因素
要想增加氧传递的推动力(C*一CL),就必须设法提高C*或降低CL。
1、提高饱和溶氧浓度C*的方法
A、温度:降低温度
B、溶液的性质:一般来说,发酵液中溶质含量越高,氧的溶解度越小。
C、氧分压:在系统总压力小于0.5MPa时,氧在溶液中的溶解度只与氧的分压成直线关系。气相中氧浓度增加,溶液中氧浓度也增加。
氨基酸合成的需氧程度产生上述差别的原因,是由它们的生物合成途径不同所引起的,不同的代谢途径产生不同数量的NAD(P)H,当然再氧化所需要的溶氧量也不同。第一类氨基酸是经过乙醛酸循环和磷酸烯醇式丙酮酸羧化系统两个途径形成的,产生的NADH量最多。因此NADH氧化再生的需氧量为最多,供氧愈多,合成氨基酸当然亦愈顺利。第二类的合成途径是产生NADH的乙醛酸循环或消耗NADH的磷酸烯醇式丙酮酸羧化系统,产生的NADH量不多,因而与供氧量关系不明显。第三类,如苯丙氨酸的合成,并不经TCA循环,NADH产量很少,过量供氧,反而起到抑制作用。肌苷发酵也有类似的结果。由此可知,供氧大小是与产物的生物合成途径有关
这个理论假定在气泡和包围着气泡的液体之间存在着界面,在界面的气泡一侧存在着一层气膜,在界面液体一侧存在着一层液膜,气膜内的气体分子与液膜中的液体分子都处于层流状态,分子之间无对流运动,因此氧分子只能以扩散方式,即借助于浓度差而透过双膜,另外,气泡内除气膜以外的气体分子处于对流状态,称为气流主体,在空气主流空间的任一点氧分子的浓度相同,液流主体亦如此。
发酵液中L-异亮氨酸提取工艺研究

lm n 活性 炭 添加量 为 2 脱 色温度 为 6 ℃ 、 色时间 为 2 ri、 色 时的最佳 p 为 47 在此 工 艺条 O i、 %、 0 脱 5 n脱 a H .。
件 下 , 一 亮氨酸提 取 收率 为 9 . , L 异 43 产品 纯度 高达 9 .% 。 % 65 关键 词 : 一 亮氨 酸 L 异 分 离提 取 工 艺 活性 炭 脱 色
了初 步研 究。 别考察 了蛋 白去 除温 度及 时 间、 分 活性 炭 用量 、 色时 间、 色温度 、 脱 脱 发酵 液 p 值 对 L 异 H 一
亮氨 酸 分 离提 取 效 果 的 影 响 。 最 终确 定 了提 取 的 工 艺条 件 . 即蛋 白去 除 温度 及 时 间 分 别 为 9 ℃和 0
1 % 00
酸 , 导致 食 欲不 振 、 质 下 降 、 将 体 贫血 及 其 它功 能 障碍 。因此 , 其用 量逐 年增 长【, 11 , 。目前 , 2 3 在工业 生
产上 主要应 用发 酵法 生产 L 异亮 氨 酸 。 一 生产 上 分 离提 取 氨基 酸 的方法 有 很 多种 , 如 离 子交换 法 、膜分 离技 术等 。本 文采 用加 热除 蛋 白 、 电点 有机 溶剂 法从 发 酵液 中提 取 L 异亮 氨 等 一 酸 。确立 了最 适蛋 白去 除温 度及 时 间 、活性 炭 用 量 、 色 时间 、 色温 度 、 酵液 p 脱 脱 发 H值 对 L 异 亮 一 氨 酸分离 提取 的影 响 。
的发 酵液 ,分别在 6 ℃、0 、0 9 ℃、O  ̄下 O 7 ℃ 8 ℃、O I0C 加热 l mi O n除蛋 白, 比较结 果如 表 1 所示
表 1 加 热 温 度 对 去 除 蛋 白量 的 影 响
发酵液溶解氧浓度控制及溶氧系数的测定
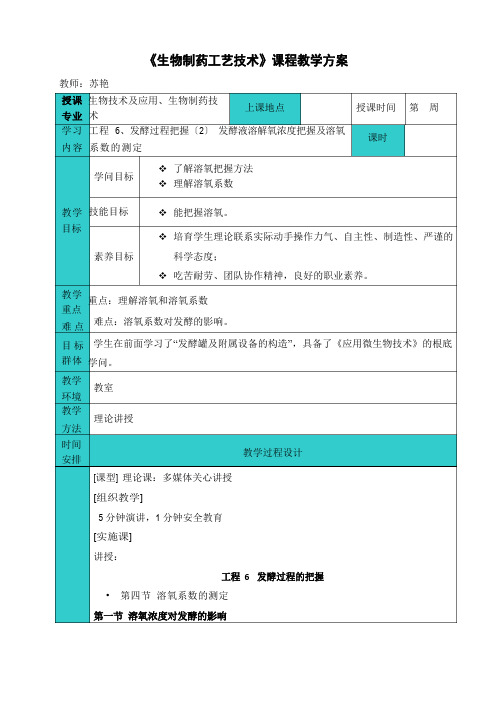
《生物制药工艺技术》课程教学方案教师:苏艳授课生物技术及应用、生物制药技专业术上课地点授课时间第周学习工程6、发酵过程把握〔2〕发酵液溶解氧浓度把握及溶氧课时内容系数的测定学问目标教学技能目标目标❖了解溶氧把握方法❖理解溶氧系数❖能把握溶氧。
❖培育学生理论联系实际动手操作力气、自主性、制造性、严谨的素养目标科学态度;❖吃苦耐劳、团队协作精神,良好的职业素养。
教学重点:理解溶氧和溶氧系数重点难点难点:溶氧系数对发酵的影响。
目标学生在前面学习了“发酵罐及附属设备的构造”,具备了《应用微生物技术》的根底群体学问。
教学教室环境教学理论讲授方法时间安排[课型] 理论课:多媒体关心讲授[组织教学]5 分钟演讲,1 分钟安全教育[实施课]讲授:教学过程设计工程6发酵过程的把握•第四节溶氧系数的测定第一节溶氧浓度对发酵的影响一、供氧与微生物的关系•发酵液中溶解氧的多少,一般以溶氧系数Kd值表示。
•各种微生物的氧化酶体系〔如过氧化氢酶、细胞色素氧化酶、黄素脱氢酶、多酚氧化酶等〕不同,在不同条件下,其呼吸程度是不同的。
微生物的吸氧量常用呼吸强度和耗氧速率两种方法来表示。
◆呼吸强度:指单位质量干菌体在单位时间内所吸取的氧量,以QO2 表示,单位为mmol/(g.h)。
◆呼吸强度可以表示微生物的相对需氧量,但当培育液中有固定成分存在时,测定就有困难,这时可用耗氧速率来表示。
◆耗氧速率:指单位体积培育液在单位时间内的吸氧量,以r 表示,单位为mmol/(L.h)。
◆耗氧速率取决于呼吸强度和菌体浓度。
式中:•γ为微生物耗氧速率,mmol/(L.h);•Q O2为呼吸强度,单位为mmol/(g.h);•ρ为菌体的质量浓度,g/L。
•发酵的不同阶段对氧的要求不同:•举例:一般菌体生长生殖期比谷氨酸生成期对溶氧要求低,长菌阶段供氧为菌体需氧量的“亚适量”,要求溶氧系数K d 为 4.0×l0-6 ——5.9×l0-6mol/(ml.min.mPa) ,形成谷氨酸阶段要求溶氧系数K d 为1.5×l0-5——1.8×l0-5 mol/(ml.min.mPa)。
L_异亮氨酸菌种选育及发酵条件优化(1)

21 1 L2异亮氨酸产生菌选育谱系 从野 生 型 谷 氨 酸 产 生 菌 栖 糖 蜜 棒 杆 菌
( Cory nebacteri um mel assecol a ) A TCC17965 为 出 发菌株 ,经硫酸二乙酯和60 Co 诱变处理 ,依次赋予 S Gr 、L eu2M Er 、L eu2 、A HVr 、Sucg 、Et hr 遗传标记 ,得 到一株 L2异亮氨酸产生菌 I3125 ,在适宜的条件下 摇瓶产酸平均为 14~15 g/ L . 选育过程见图 1. 21 2 发酵培养基优化
M Er + L eu2 + A HVr + Sucg + Et hr ) ,在培养基未经优化的情况下产 L2异亮氨酸 14~的影响 ,在优化的培养基和发酵条件下积累 L2异亮氨酸 191 2
g/ L ,最高时可达 211 3 g/ L .
完全培养基 、基本培养基和种子培养基均以 20 %的 NaO H 调至 p H 71 0 , 01 1 M Pa 下 灭菌 20 min ; 发酵培养基以 20 %的 NaO H 调 p H 至 71 0 , 01 07 M Pa 压力灭菌 10 min. 11 3 培养方法 11 31 1 种子培养 接一环生长良好的斜面种子于 装有 40 mL 种子培养基的 250 mL 三角瓶中 ,30 ℃ 于往复式摇床上培养 11 h , 85 r/ min ,振幅 90 mm. 11 31 2 发酵培养 接 1 mL 种子液于装有 15 mL 发酵培养基的 250 mL 三角瓶中 ,30 ℃于往复式摇 床上培养 72 h , 100 r/ min ,振幅 90 mm. 11 4 诱变筛选方法
要[11] . 为此作者考察了在发酵培养基中分别添加不 同氮源 (含氮量以硫酸铵 40 g/ L 的用量的含氮量来 计算其它氮源的添加量) 对 I3125 合成 L2Ile 影响 , 结果见图 3.
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
收稿日期: 2006 08 09 修回日期: 2006 12 28 * 天津市科技攻关项目 ( 05Y FG PGX 07000 ) ** 通讯作者, 电子信箱: w en ty@ im. ac. cn
色氨酸产量达到 9. 03g /L。 现阶段国内外对 L 异亮氨酸途径分析的研究还比
较欠缺, 因而本文以途径分析理论为指导, 确定了黄色 短杆菌由葡萄糖生产 L 异亮氨酸的最佳途径。根据途 径分析的结果利用改变溶氧的方法来改变细胞内部的 代谢流 分 布, 使 其在 特 定环 境下 生成 更 多的 L 异 亮 氨酸。
经途径分析 后, 一个特殊的 代谢路径的 所有酶 被 过量表达时, 编码不必要的酶的基因被敲除, 就会构建 出一个高产的路径。为了避免通过 TCA 循环和乙醛酸 支路生成 OAA 而造成碳源的浪费, 可以通过过量表达 PEP羧化 酶的基因直接使 PEP 生成 OAA 或过量表 达 丙酮酸激酶的基因和丙酮酸 羧化酶的基因通过 Pyr生
通过途径分析可知, 葡 萄糖进入 L 异亮氨酸生 成 途径有两种方式, 一是 PEP Pyr A c C oA OAA, 另一 种是 CO2 的回补反应, 即 PE P直接生成 OAA。若通过 TCA 循环生成 OAA, 则会生成 CO2 造成 碳源损失。在 模型 1和模型 2 中, 由 1m ol葡 萄糖生成 1m olL 异亮 氨 酸, 产率高达 100% , 见图 1。其 原因就是 PEP或 PYR 直接生成 OAA, 且葡萄糖由 EM P途径进入代谢网络而 不通过 HM P 途 径进 入代谢 网络, 从而避 免了 碳源 的 浪费。
Vo .l 27 No. 2 2007
Hale Waihona Puke 但谷氨酸代谢流却 明显降低, 在 发酵中后期 控制较 低 溶氧, 谷氨酸合成代谢流较大。 X iao等 [13] 研究了不 同 溶氧对谷氨酸发酵中两个关键酶 ( 谷氨酸脱氢酶 GDH 和乳酸脱氢酶 LDH ) 和代谢流的影响, 研究表明, 在 过 低溶氧条件下, TCA 循环代谢流量减小, 不足以平衡葡 萄糖酵解速率, 从而刺激了 LDH 的酶活, 使代谢流转向 乳酸生成, 造成乳酸积累; 而过高溶氧, GDH 酶活明 显 降低, 且 TCA 循环流 量加大, 生成 大量 CO2, 造成碳 源 损失, 两种情况 均不利于谷 氨酸生成。刘 勇等 [ 2] 的 研 究表明, 在 L 异亮氨酸发酵中, 转速不同对各种氨基酸 产量有明 显 影 响, 低 转 速时 产 生 杂 酸 较 多。 宋 文 军 等 [14] 给出了 L 异 亮氨酸发 酵中后 期不同转 速下的 代 谢流分布, 研究表 明, 转速过低, TCA 循环代 谢流量 减 小, 但副产氨基酸流量增加, 差不多每一种副产氨基酸 的代谢流均要比高 转速时要大, 尤其是副产 缬氨酸 明 显过多。随着转速增加, TCA 循环代谢流量增加, 各副 产氨基酸流量减小, 这说明供氧充足有利于抑制杂酸。 但是供氧过高又会造成 TCA 途径流量过大, 导致 L 异 亮氨酸的代谢流减小, 这说 明控制合适 的溶氧对 L 异 亮氨酸发酵至关重要。若能在发酵过程中恰当控制溶 氧, 就可以改变代谢流 量, 从而增大 L 异亮氨酸 产率, 减少杂酸生成。
感分析仪测定。 1. 4. 2 OD 值的 测定 将发酵液稀释 一定倍数后, 在 波长 620nm 处用分光光度计测定。 1. 4. 3 发酵液中氨基酸含量测定 采用 E lite AAA 型 氨基酸分析仪测定。
2 结果和讨论
2. 1 L 异亮氨酸生物合成途径分析 根据 L 异亮氨酸代谢网络以及各步反应的化学计
摘要 利用途径分析方法对黄色短杆菌 ( B revibacterium f lavum ) TC 21 生产 L 异亮氨酸的途径进 行了分析, 确定了黄色短杆菌 TC 21生产 L 异亮氨酸的最佳途径的通量分布, 根据途径分析的结 果, 发酵过程中的代谢流量对 L 异亮氨酸产量有明显影响, 而代谢流量与发酵过程中的溶氧密切 相关, 因此可以通过控制溶氧来提高 L 异亮氨酸产量。在发酵过程的不同阶段, 根据菌体生长和 产酸的需求, 改变合成途径的代谢流量, 可以有效提高产酸率。实验证明, 通过溶氧分阶段控制 发酵生产 L 异亮氨酸, 比溶氧恒定控制方式发酵产率提高了 15. 77% 。实验结果说明, 用途径分 析的结果指导发酵过程中的溶氧可以大幅度提高 L 异亮氨酸的产量。 关键词 L 异亮氨酸 途径分析 溶氧控制 黄色短杆菌 中图分类号 Q815
种子培 养基: 葡萄糖 30, KH2 PO4 1. 5, M gSO4 0. 4, FeSO4 0. 01, M nSO4 0. 01, ( NH 4 ) 2 SO4 3, 尿 素 2, 豆饼水 解液 20m l/L, 生物素 0. 5m g/L, VB1 2. 5m g /L, pH 7. 0~ 7. 2。
量平衡式, 可由计算机计算出该代谢可分为 9 个模型。 产率最高的基本模型代表了最优途径。最优途径和通 量分布见图 1[ 5]。
图 1 由葡萄糖到 L 异亮氨酸的最优途径和 通量分布 F ig. 1 Op tim al pathways and f lux d istr ibu tion for converting glucose to L isoleu cine
图 2 L Ile发酵在不同 转速下 DO 的变化 F ig. 2 Change of DO of d ifferen t stirr ing speed
in L Isoleucine ferm enta tion
图 3 不同转速对生长速率和产酸速率的影响 虚心表示转速对产酸的影响, 实心表示转速对 O D620的影响
种子 培 养: 接 一 环生 长 良好 的 斜 面种 子 至 装 有 25m l种子培养基的 500m l摇瓶中, 置于 旋转式摇床 上 ( 200 r/m in ), 31 振荡培养 38h。
发酵: 按 15%接种量将种子液接入 5L 自动控制发 酵罐中, 装液量 3L, 通风量 1 ~ 3L /m in, 根据 溶氧控 制 搅拌转速, 培养温度 31 , 通过自动流加浓氨水溶液控 制 pH 7. 0, 发酵 88h。 1. 4 分析方法 1. 4. 1 发酵液中残糖测定 采用 SBA 40 系列生物传
发 酵 培 养 基: 葡 萄 糖 70, ( NH 4 ) 2 SO4 15, FeSO4 0. 015, M gSO4 0. 5, M nSO4 0. 015, KH 2 PO4 1. 5, K2H PO4
2007, 27( 2)
杨 宁 等: 基于途径分析的 L 异亮氨酸发酵溶 氧控制研究
71
3, 生物素 140 g /L, VB1 0. 1m g /L, M et 0. 02, 豆饼水 解 液 20m l/L, 玉米浆 25m l/L, pH 7. 0 ~ 7. 2。 1. 3 培养方法
中国生物工程杂志 Ch ina B iotechnology, 2007, 27( 2): 70~ 75
基于途径分析的 L 异亮氨酸发酵溶氧控制研究*
杨 宁 1 王 健 2 徐庆阳 1 陈 宁 1 温廷益 1, 3**
( 1 天津科技大学生物工程学院 天津 300222 2 吉林大学生物与农业工程学院 长春 130022) ( 3 中国科学院微生物研究所 北京 100080)
1 材料和方法
1. 1 供试菌株 黄色短杆 菌 B rev iba cterium f lavum TC 21 (M et- +
E thr + ABr + AEC r ) , 天津科技大学代谢控制发酵研 究室保藏菌种。 1. 2 培养基 ( g /L )
斜面培养基: 葡萄糖 1, 酵母膏 5, 蛋白胨 10, 氯化 钠 2. 5, 琼脂 20, pH 7. 0~ 7. 2。
成 OAA。为了避 免 因 生成 CO2 而 造 成浪 费, 应该 对 6 磷酸葡萄糖脱 氢酶进 行截 堵。按 照途 径分析 结果, HM P途径、TCA 循环和乙醛酸支路应该减弱, 从而使碳 架流转向异亮氨酸的合成, 因为 TCA 循环 中的酮戊二 酸脱氢酶和柠 檬酸合成酶 催化的反应 是不可逆 的, 能 量和许多中间代谢物需要 TCA 循环来 提供, 所以该循 环中的酶不能被完全去除。现阶段国内外各种研究表 明 [ 2, 3, , 12~ 14] 溶氧对代谢 过程有显 著影响, 故本文 采用 控制溶氧的方法来改变 L 异亮氨酸代谢, 进而提高 L 异亮氨酸产率。
虽然溶氧影响 L 异亮氨酸合成代谢的机理尚未完 全清楚, 但国内 外已有多人 对溶氧与代 谢过程的 关系 进行了研究。 Zhang等 [ 12] 给出了谷氨酸发 酵过程中溶 氧与代谢流量的量化关系, 研究表明, 在其它条件相同 情况下, 高溶氧条件下 TCA 循环代谢流量大于低溶氧,
72
中国生物工程杂志 China B io techno logy
F ig. 3 Rate of grow th and rate of L Iso leuc ine by d ifferent revolution s p er m inu te
由图 2 可知, 在控制不同转速的发酵过程中, 相对 溶氧 ( DO )均表现出相似的变化规律, 但发酵的不同阶 段对氧的需求不同。在发酵初期 ( 0 ~ 32h ) , 菌体耗 氧 速率明显快于供氧速率, 表现为相对溶氧的迅速下降; 而 32h后, 好氧速率 和供氧速 率基本 保持平衡。发 酵 末期 ( 60h 后 ) , 相对溶 氧回 升, 发 酵结 束相 对溶 氧 达 40% 以上。 2. 2. 2 不同转速对 L 异亮 氨酸生产菌 生长和产酸 的 影响 在其它发 酵条件相同 的情况下, 控制 不同转 速 进行 5L 罐的分批发酵, 得到发酵过程不同时刻生长速 率与产酸速率随时间的变化曲线结果, 如图 3所示。