第二节真核基因转录水平的调控(精)
第二章 真核基因表达与调控

列开始
⑵起始子(initiator, inr)
• 以转录点为中心的的保守序列,
• 共有序列为PyPyAN(A/T) PyPy,Py-嘧啶, • 功能:是某些基因最佳转录所需的
⑶下游启动子元件
(down stream promotor element, DPE)
第五章 真核基因表达调控
真核基因表达调控的基本特点
• (管家基因)组成性表达;(奢侈基因) 可变性表达(诱导和阻遏)
• 时间(发育相关性)和空间特异性(组 织细胞相关性)
• 正性调控为主
第二节 真核基因表达调控的分子机制
一、顺式作用元件(cis-acting element)
概念: 真核生物中能够被基因调控蛋白特异性识别和结 合,并对自身基因转录起始有调节作用的DNA序列。
• Ⅱ类顺式作用元件包括: 核心启动子( Core promoter),增强子(enhancer),沉 默子(silencer ),及各种反应元件等。
1. 核心启动子( Core promoter)
• Ⅱ类启动子的核心启动子常由TATA盒、位于 TATA盒上游的的上游启动子元件、以转录点 为中心的起始子和下游启动子元件,4个元件 组合而成。
还需TF IIIA
A box (+10~+20) A box (+10~+20) TF III C
TF III C TF III B TF III B TF III C RNA pol III TF RIINI BA polTIIFIIII C
RNA聚合酶III的转录起始
(三) RNA pol II 转录体系的调节
真核生物转录水平的调控机制

真核生物转录水平的调控机制一、转录因子转录因子是真核生物转录水平调控的重要环节。
它们可以识别和结合DNA上的特异序列,从而调控基因的表达。
根据结合位点的不同,转录因子可以分为上游启动子元件和增强子元件两类。
上游启动子元件主要包括TATA box和CAAT box等,而增强子元件则是一种具有增强基因转录功能的DNA序列。
二、染色质重塑染色质重塑是真核生物基因表达调控的重要机制之一。
染色质重塑可以改变染色质的结构,从而影响基因的表达。
染色质重塑过程中,染色质重塑复合物可以将核小体从DNA上移除或重新排列,从而改变染色质的可及性。
此外,染色质重塑还可以影响DNA的甲基化水平,进一步调控基因的表达。
三、miRNA和siRNAmiRNA和siRNA是真核生物中的非编码RNA,它们可以通过与mRNA的特异性结合来调控基因的表达。
miRNA和siRNA可以与mRNA 的3'UTR结合,导致mRNA的降解或翻译抑制,从而调控基因的表达。
此外,miRNA和siRNA还可以通过与转录因子或染色质重塑复合物等相互作用,影响基因的转录和表达。
四、转录起始和延伸转录起始和延伸是真核生物转录水平调控的重要环节。
转录起始和延伸过程中,RNA聚合酶可以识别启动子元件并开始转录,然后沿着DNA序列向下游移动并合成RNA。
在这个过程中,转录起始和延伸复合物可以与RNA聚合酶相互作用,从而影响转录的效率和方向。
此外,一些转录因子也可以与RNA聚合酶相互作用,进一步影响基因的表达。
五、转录后修饰真核生物中的RNA聚合酶可以使用各种转录后修饰来修饰其转录产物。
这些修饰可以包括mRNA的加尾、编辑、剪接和稳定性等。
这些过程可以影响mRNA的翻译效率和稳定性,从而影响基因的表达。
此外,一些蛋白质也可以通过磷酸化、乙酰化或甲基化等修饰来影响基因的表达。
六、细胞周期与细胞分化细胞周期和细胞分化是真核生物细胞生命活动中的重要过程,也是转录水平调控的重要方面。
真核生物DNA水平上的基因表达调控

复杂多基因家族 复杂多基因家族一般由几个相关基因家族构成,基因家族之
间由间隔序列隔开,并作为独立的转录单位。现已发现存在不 同形式的复杂多基因家族。
海胆的组蛋白基因家族 串联单位中的每一个基因分别被转录成单顺反子 RNA,这些RNA都没有内含子,而且各基因在同一条DNA链上按同一方 向转录,每个基因的转录与翻译速度都受到调节。 研究还表明,在一个特定的细胞中,并不是所有串联的单位都得到转录。 胚胎发育的不同阶段或不同组织中,有不同的串联单位被转录,暗示可 能存在具有不同专一性的组蛋白亚类和发育调控机制。
一、基因家族 二、真核基因的断裂结构 三、真核生物DNA水平上的基因表达调控 四、DNA 甲基化与基因活性的调控
前述:真核基因组的一般构造特点
① 在真核细胞中,一条成熟的mRNA链只能翻译出一条多肽 链,不存在原核生物中常见的多基因操纵子形式。
② 真核细胞DNA都与组蛋白和大量非组蛋白相结合,只有一 小部分DNA是裸露的。
真核生物基因调控,根据其性质可分为两 大类:
第一类:瞬时调控或称可逆性调控
它相当于原核细胞对环境条件变化所做出的反应,包 括某种底物或激素水平升降及细胞周期不同阶段中酶 活性和浓度的调节。
第二类:发育调控或称不可逆调控
是真核基因调控的精髓部分,它决定了真核细胞生长、 分化、发育的全部进程。
在真核生物中基因表达的调节其特点是:
(1)多层次; (2)无操纵子和衰减子; (3)个体发育复杂; (4)受环境影响较小;
研究基因调控主要应回答3个问题:
① 什么是诱发基因转录的信号? ② 基因调控主要是在哪一步(模板DNA的转录、mRNA
的成熟或蛋白质合成)实现的? ③ 不同水平基因调控的分子机制是什么?
分子生物学:真核基因表达调控

真核基因表达的多级调控
在真核生物中基因表达的调节其特是
(1)多层次; (2)无操纵子和弱化子; (3)个体发育复杂; (4)受环境影响较小;
研究基因调控3个问题:
① 什么是诱发基因转录的信号?
基因扩增是指某些基因的拷贝数专一性大量增加的现象,它 使细胞在短期内产生大量的基因产物以满足生长发育的需要,是 基因活性调控的一种方式。
实例: 非洲爪蟾的卵母细胞中原有rRNA基因(rDNA)约500个拷
贝,在减数分裂I的粗线期,这个基因开始迅速复制,到双线 期它的拷贝数约为200万个,扩增近4000倍,可用于合成1012个 核糖体,以满足卵裂期和胚胎期合成大量蛋白质的需要。
二、基因扩增、基因重排和基因丢失
三、DNA甲基化与基因活性的调控
一、 染色质结构对转录的影响
按功能状态的不同可将染色质分为: (1)活性染色质(有转录活性) (2)非活性染色质(没有转录活性)
染色质的核小体发生构象改变,松散的染色质结构,便 于转录调控因子和顺式用元件结合和RNA聚合酶在转录模板上 滑动。
真核基因调控中虽然也发现有负性调控元件,但其存在并不 普遍;
顺式作用元件: 由若干可以区分的DNA序列组成,并与特定的功能
基因相连,组成基因转录的调控区,通过与相应的反 式作用因子结合,实现对基因转录的调控。
反式作用因子: 能直接地或间接地识别或结合在各类顺式作用元
件核心序列上,参与调控靶基因转录效率的蛋白因子, 也被称为转录因子(TF)。
哺乳类基因组中约存在4万个CpG 岛,它们大多位于结构基 因启动子的核心序列和转录起始点,其中有60%~ 90% 的 CpG 被甲基化, CpG 岛在基因表达调控中起重要作用。
普通遗传学第十四章 基因表达的调控
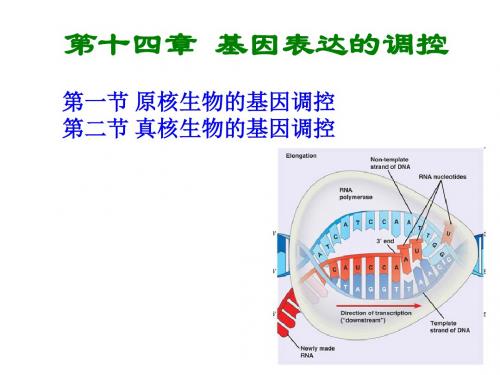
第二节 真核生物的基因调控
一、 DNA水平的调控 二、染色质水平调控 三、转录水平的调控 四、翻译水平的调控
一 DNA水平的调控
1、基因丢失 2、基因扩增 3、基因重排 4、DNA甲基化
一、DNA水平的调控 1、基因丢失
某些原生动物,如线虫、昆虫和甲克类动 物在个体发育过程中,许多体细胞经常丢掉 整个或者部分染色体,只有将要分化形成生 殖细胞的细胞中保留全部染色体。
3、基因重排
基因重排:DNA分子核苷酸序列的重新排 列。重排不仅可以形成新的基因,还可以调 节基因表达。基因组中的DNA序列重排并 不是一种普遍方式,但它是一些基因调控的 重要机制。
① 酵母交配型转换 →a 这种交配型转换的基础是遗传物质的重排。 控制交配型的MAT(mating-type)基因位于酵母菌 第3染色体上,MATa和MAT互为等位基因。
第一节 原核生物的基因调控 一、转录水平的调控 二、翻译水平的调控
二、翻译水平的调控
1、反馈调控机制
如果某种蛋白质过量积累,将与其自身的 mRNA结合,阻止进一步翻译。这种结合位点 通常包括mRNA 5’端非翻译区,也包括启动子 区域的 Shine-Dalgarno (SD) (AGGAGGU) 序 列。
(二)组蛋白质修饰和非组蛋的作用
组蛋白可被修饰,修饰可改变其与DNA的接 合能力。若被组蛋白覆盖的基因要表达,那么 组蛋白必须被修饰,使其和DNA的结合由紧 变松,这样DNA链才能和RNA聚合酶或调节 蛋白相互作用。因此组蛋白的作用本质上是真 核基因调节的负控制因子,即它们是基因表达 的抑制物。 非组蛋白打开特异基因的分子,具有组织特异 性,在基因表达的调节、细胞分化的控制以及 生物的发育中起着很重要的作用。
免疫球蛋白的多样性
生物化学(本科)第十章基因表达调控及其相关细胞信号转导通路随堂练习与参考答案

⽣物化学(本科)第⼗章基因表达调控及其相关细胞信号转导通路随堂练习与参考答案⽣物化学(本科)第⼗章基因表达调控及其相关细胞信号转导通路随堂练习与参考答案第⼀节概述第⼆节原核基因的转录调控第三节真核基因的转录调控第四节相关细胞信号转导通路1. (单选题)基因表达调控的基本控制点是( )A. 基因结构活化B. 转录起始C. 转录后加⼯D. 蛋⽩质翻译及翻译后加⼯E. mRNA从细胞核转运到细胞浆参考答案:B2. (单选题)分解代谢物基因激活蛋⽩(CAP)对乳糖操纵⼦表达的影响是( )A. 正性调控B. 负性调控C. 正性调控、负性调控都可能D. ⽆调控作⽤E. 可有可⽆参考答案:A3. (单选题)阻遏蛋⽩识别操纵⼦中的( )A.启动基因B.结构基因C.操纵基因D.内含于E.外显⼦参考答案:C4. (单选题)⽬前认为基因表达调控的主要环节是( )A. 翻译后加⼯B. 转录起始C. 翻译起始D. 转录后加⼯E. 基因活化A. 适应环境B. 调节代谢C. 维持⽣长D. 维持分裂E. 维持分化参考答案:A6. (单选题)⼀个操纵⼦通常含有( )A.⼀个启动序列和⼀个编码基因B.⼀个启动序列和数个编码基因C.数个启动序列和⼀个编码基因D.数个启动序列和数个编码基因E.两个启动序列和数个编码基因参考答案:B7. (单选题)对乳糖操纵⼦来说( )A. CAP是正性调节因素,阻遏蛋⽩是负性调节因素B. CAP是负性调节因素,阻遏蛋⽩是正性调节因素C. CAP和阻遏蛋⽩都是正性调节因素D. CAP和阻遏蛋⽩都是负性调节因素E. 在不同条件下,CAP和阻遏蛋⽩均显⽰正性或负性调节特点参考答案:A8. (单选题)lac阻遏蛋⽩由( )A. Z基因编码B. Y基因编码C. A基因编码D. I基因编码E. 以上都不是参考答案:D9. (单选题)基因表达产物是( )A. RNAB. DNAC. 蛋⽩质D. 酶和DNAE. ⼤多数是蛋⽩质,有些是RNAA. 作为阻遏物结合于操纵基因B. 作为辅阻遏物结合于阻遏物C. 使阻遏物变构⽽失去结合DNA的能⼒D. 抑制阻遏基因的转录E. 使RNA聚合酶变构⽽活性增加参考答案:C11. (单选题)顺式作⽤元件是指( )A. TATA box和CCAAT boxB. 基因的5’-侧翼序列C. 基因的3’-侧翼序列D. 具有转录调节功能的特异DNA序列E. 增强⼦参考答案:D12. (单选题)反式作⽤因⼦是指( )A. 具有激活功能的调节蛋⽩B. 具有抑制功能的调节蛋⽩C. 对另⼀基因具有激活功能的调节蛋⽩D. 对另⼀基因表达具有调节功能的蛋⽩E. 是特异DNA序列参考答案:D13. (单选题)启动⼦是指( )A. DNA分⼦中能转录的序列B. 与RNA聚合酶结合的DNA序列C. 与阻遏蛋⽩结合的DNA序列D. 有转录终⽌信号的DNA序列E. 与反式作⽤因⼦结合的RNA序列参考答案:B14. (单选题)增强⼦的作⽤是( )A. 促进结构基因转录B. 抑制结构基因转录C. 抑制阻遏蛋⽩D. 抑制操纵基因表达E. 抑制启动⼦A. 操纵基因结合B. 启动⼦上游的CAP位点结合C. DNA分⼦中任意⼀段序列结合D. 增强⼦结合E. 沉默⼦结合参考答案:B16. (单选题)通过胞内受体发挥作⽤的信息物质为 A.⼄酰胆碱B.γ-氨基丁酸C.胰岛素D.甲状腺素E.表⽪⽣长因⼦参考答案:D17. (单选题)细胞内传递信息的第⼆信使是A.受体B.载体C.⽆机物D.有机物E.⼩分⼦物质参考答案:E18. (单选题)下列哪项不是受体与配体结合的特点 A.⾼度专⼀性B.⾼度亲和⼒C.可饱和性D.不可逆性E. ⾮共价键结合参考答案:D19. (单选题)通过膜受体起调节作⽤的激素是A.性激素B.糖⽪质激素C.甲状腺素D.肾上腺素E.活性维⽣素D3参考答案:D20. (单选题)胞内受体的化学本质为F.糖脂参考答案:A21. (单选题)IP3与相应受体结合后,可使胞浆内哪种离⼦浓度升⾼A.K+B.Na+C.HCO3-D.Ca2+E. Mg2+参考答案:D22. (单选题)在细胞内传递激素信息的⼩分⼦物质称为 A.递质B.载体C.第⼀信使D.第⼆信使E.第三信使参考答案:D23. (多选题)真核⽣物基因结构特点是( )A.基因不连续性B.单顺反⼦C.含重复序列D.⼀个启动基因后接有⼏个编码基因E.含内含⼦参考答案:ABE24. (多选题)哪些是顺式作⽤元件? ( )A.启动⼦B.增强⼦C.内含⼦D.反应元件E.外显⼦参考答案:ABD25. (多选题)乳糖操纵⼦中具有调控功能的基因是( )A.A基因参考答案:CD26. (多选题)下列对增强⼦特征描述中,正确的是( )A.增强⼦可远距离发挥作⽤B.增强效应的位置和⽅向⽆关C.可通过启动⼦提⾼同⼀条链上的靶基因的转录效率D.没有基因的专⼀性,可在不同的基因组合上表现增强效应E.增强⼦是具有转录调节功能的特异DNA序列参考答案:ABCE27. (多选题)以下关于cAMP对原核基因转录的调控作⽤的叙述正确的是( )A.cAMP可与分解代谢基因活化蛋⽩(CAP)结合成复合物B.cAMP-CAP复合物结合在启动⼦前⽅C.葡萄糖充⾜时,cAMP⽔平不⾼D.葡萄糖和乳糖并存时,细菌优先利⽤乳糖E.葡萄糖和乳糖并存时,细菌优先利⽤葡萄糖参考答案:ABCE28. (多选题)以下关于反式作⽤因⼦的叙述哪些是正确的? ( )A.反式作⽤因⼦是调节基因转录的⼀类蛋⽩因⼦B.转录因⼦是⼀类反式作⽤因⼦C.增强⼦结合蛋⽩属反式作⽤因⼦D.阻遏蛋⽩是⼀类负调控反式作⽤因⼦E.RNA聚合酶是反式作⽤因⼦参考答案:ABCD29. (多选题)以下关于顺式作⽤元件的叙述哪些是正确的? ( )A.顺式作⽤元件是⼀类调节基因转录的DNA元件B.增强于是⼀类顺式作⽤元件C.启动⼦中的TATA盒和GC盒都是顺式作⽤元件D.操纵基因是原核⽣物中的⼀类负调控顺式作⽤元件E.顺式作⽤元件只对基因转录起增强作⽤参考答案:ABCD30. (多选题)受体与配体结合的特点包括A.⾼度专⼀性E 特定的作⽤模式参考答案:ABCDE31. (多选题)能与GDP/GTP结合的蛋⽩质是A.G蛋⽩B.Raf蛋⽩C.Rel A蛋⽩D.Grb-2蛋⽩E.Ras蛋⽩参考答案:AE32. (多选题)与配体结合后,⾃⾝具有酪氨酸蛋⽩激酶活性的受体是A.胰岛素受体B.表⽪⽣长因⼦受体C.⾎⼩板衍⽣⽣长因⼦受体D.⽣长激素受体E.⼲扰素受体参考答案:ABC33. (多选题)胞内受体的激素结合区能A.与配体结合B.与G蛋⽩偶联C.与热休克蛋⽩结合D.使受体⼆聚体化 E.激活转录参考答案:ACDE。
第二节真核基因转录水平的调控(精)

第二节真核基因转录水平的调控一、真核生物的RNA聚合酶有三种RNA聚合酶:RNA聚合酶Ⅰ;RNA聚合酶Ⅱ;RNA聚合酶Ⅲ。
二、真核基因顺式作用元件(一)、顺式作用元件概念指DNA上对基因表达在调节活性的某些特定的调控序列,其活性仅影响其自身处于同一DNA分子上的基因。
(二)、种类启动子、增强子、静止子1、启动子的结构和功能启动子与原核启动子的含义相同,是指RNA聚合酶结合并起动转录的DNA序列。
但真核同启动子间不像原核那样有明显共同一致的序列。
而且单靠RNA聚合酶难以结合DNA而起动转录,而是需要多种蛋白质因子的相互协调作用。
RNA聚合酶Ⅱ启动子结构1)TATA框(TATA frame):其一致顺序为TATAA(TAA(T。
TATA框中心在-30附近,相当于原核的-10序列(pribnow box)。
对大多数真核生物来说,RNA聚合酶与TATA框牢固结合之后才能开始转录。
TATA框的左右富含G┇C 序列,这就有利于该框与RNA聚合酶形成开放性启动子复合物。
2)CAAT框(CAAT frame):位置在-75附近,一致序列为GGC(TCAATCT。
CAAT框可能控制着转录起始的频率。
(3)GC框在-90bp左右的GGGCGG序列称为GC框。
一个在-30—+15即核心启动子(core promoter element,另一为上游启动子区(upstream promoter element在-150—-50,不同物种的启动子因子有显著差异,启动子区没有和mRNA的TATA和CAAT盒顺序,故物种间大前体-rRNA基因的转录起始是不同的。
基因间间隔含一个或几个终止信号可终止其之前的基因的转录而其本身不转录,间隔区含多种反向顺序可作为增强子结合转录因子2、增强子的结构和功能增强子(enhancer):又称为远上游序列(far upstream sequence 。
它是远距离调节启动子以增加转录速率的DNA序列,其增强作用与序列的方向无关,与它在基因的上下游位置无关。
真核生物基因转录水平的反式调控ppt

从功能上讲,植物Myb蛋白作为转录因子参与诸如
类黄酮类化合物的生物合成、毛状体发育、分生组织增 殖、GA应答以及干旱胁迫响应等信号转导过程。而编码 植物Myb蛋白的myb相关基因 是一个很大的基因家族, 因此也能调节发育进程以及参与对外界胁迫应答相关基 因的表达。
15
• 有些转录因子同时包含特异与非特异的DNA结合域, 而且后者对于目标基因的转录激活是必需的;
29
阴阳因子-1(Yin Yang-1,YY1)是一类新的转录因 子,它通过与其它蛋白发生互作从而具有转录激活和阻 遏功能,同时YY1也是核基质结合蛋白,其结合基序的 多样性可能是其功能多样性的基础。
由于YY1的结合基序核心是 CCAT、ACAT、TCAT,
其旁侧序列也具有很大的异质性,其结合基序的可变性 很大程度上允许YY1结合和影响一大批基因的转录,而 结合位点的可变性也使 YY1因子结合基序与大量其它因 子的结合基序互相靠近或重叠,从而使转录调控更加严
同,大多数bZIP类因子的亮氨酸拉链通常包含有4个或5
个7单位重复,而在拟南芥的ATB2中则有9个。 另外寡聚化结构域以外区域的差异也会影响到其功
能。
25
细胞核定位信号 真核生物转录因子是选择性的进入细胞核的,因此 转录因子也包含以富含精氨酸和赖氨酸核心肽为特征的
NLS,而碱性核心区的功能也受旁侧残基的影响。转录
• VP1是一种植物基因(包括小麦的Em基因)的转录因
子,含有一个弱的非特异性的DNA结合域-BR2(B2) 和一个序列特异的结合域BR3(B3),能够识别Sph元
件(CATGCATG),而该元件存在于Em以及其它一些
植物基因中。 • 有意思的是,VP1是通过BR2而非BR3来激活Em表达
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第二节真核基因转录水平的调控
一、真核生物的RNA聚合酶
有三种RNA聚合酶:RNA聚合酶I; RNA聚合酶U; RNA聚合酶川。
二、真核基因顺式作用元件
(一)、顺式作用元件概念
指DNA上对基因表达在调节活性的某些特定的调控序列,其活性仅影响其自身处于同一 DNA 分子上的基因。
(二)、种类
启动子、增强子、静止子
1、启动子的结构和功能
启动子与原核启动子的含义相同,是指 RNA聚合酶结合并起动转录的 DNA序列。
但真核同启动子间不像原核那样有明显共同一致的序列。
而且单靠RNA聚
合酶难以结合DNA而起动转录,而是需要多种蛋白质因子的相互协调作用。
RNA聚合酶U启动子结构
1) TATA 框(TATA frame ):其一致顺序为 TATAA(TAA(T。
TATA 框中心在-30附近,相当于原核的-10序列(prib now box)。
对大多数真核生物来说,RNA聚合酶与TATA框牢固结合之后才能开始转录。
TATA框的左右富含G : C序列,这就有利于该框与 RNA聚合酶形成开放性启动子复合物。
2) CAAT框(CAAT frame ):位置在-75附近,一致序列为 GG
C(TCAATCT。
CAAT框可能控制着转录起始的频率。
(3)GC 框
在-90bp左右的GGGCGG序列称为GC框。
一个在-30—+15即核心启动子(core promoter elemen,另一为上游启动子区(upstream promoter elemen在-150— -50,不同物种的启动子因子有显著差异,启动子区没有和mRNA的TATA和CAAT盒顺序,故物种间大前体-rRNA基因的转录起始是不同的。
基因间间隔含一个或几个终止信号可终止其之前的基因的转录而其本身不转录,间隔区含多种反向顺序可作为增强子结合转录因子
2、增强子的结构和功能
增强子(enhance):又称为远上游序歹U( far upstream sequenee 它是远距离调节启动子以增加转录速率的 DNA序列,其增强作用与序列的方向无关,与它在基因的上下游位置无关。
增强子有强烈的细胞类型选择,即不同细胞类型,增强作用不同。
1)它能通过启动子大幅度地增加同一条 DNA链上靶基因转录的频率,一般能增加10〜200倍,有的甚至可达千倍。
(2)增强子的作用对同源或异源的基因同样有效,如把SV40的增强子连接到兔禺珠蛋白的基因上,可使转录强度增大 100倍;
(3)增强子的位置可在基因5上游、基因内或其3下游的序列中,而其作用与所在基因旁侧部位的方向似无关系,因为无论正向还是反向,它都具有增强效
应;
(4)增强子所含核苷酸序列大多为重复序列,其内部含有的核心序列,对于它进入到另一宿主之后重新产生增强子效应至关重要;
(5)增强子一般都具有组织和细胞特异性;
(6)增强子在DNA双链中没有5与3固定的方向性;
(7)增强子可远离转录起始点,通常在 1〜4 kb (个别情况可达30 kb)外起作用;
(8)增强子的活性与其在DNA双螺旋结构中的空间方向性有关。
另外,许多增强子
还受到外部信号的调控,如金属硫蛋白基因的增强子就可对环境中的锌、镉浓度作出反应。
3、静止子
类似增强子但起负调控作用的顺式元件。
静止子与反式作用因子(蛋白质) 结合后,使正调控系统失去作用。
三、转录的起始调节
(一)转录起始因子与起始复合物的装置
RNA聚合酶需要先分别同SL1、TF H D、TF E B等一些转录起始因子结合,形成转录起
始复合物(initiation complex )才能开始其转录活动。
转录因子都属于多蛋白复合物,是由 TATA结合蛋白和各自独有的一套TBP 相关因子组成。
类型II基因的转录因子普遍性转录因子:作用于基本核心启动子如TATA box、
INR(转录起始区,每种细胞类型都必需的,如TFIID/A/B/E/F/G/H/I等。
特异性转录因子:作用于转录起始复合物形成过程的靶分子和控制位点,含 DNA特异
性序列结合结构域普遍性转录因子的结构与功能TFIID的TBP(TATA
binding protein结构域结合启动子的 TATA box,促进其它转录因子的结合。
TFII I 结合INR。
许多普遍性转录因子含有与 RNA聚合酶因子相似的结构域,识别特异启动子起始转录。
和激活结构域(有的两者都有。
RNA pol.ll的结构与功能:CTD结构域含YSPTSPS的重复单位,不同物种重复数不同,CTD对转录活性是必需的,其Ser/Thr可以被不同程度磷酸化在转录起始与延伸中具有重要作用。
例:RNA聚合酶U转录起始复合物的组装
第一步是转录因子TFH D与TATA框特异性结合,形成TF II D-启动子复合体,后者进而指导聚合酶U和其他基本转录因子与启动子进行有序装配,最后形成一个稳定的起始复合物。
四、调控转录的反式作用因子
能识别或结合在顺式作用元件核心序列上参与调控靶基因转录效率的结合蛋白,称为反式作用因子。
(一)反式作用因子的结构特征
1、DNA识别或DNA结合结构域
2、激活基因转录的功能结构域
3、结合其他蛋白或调控蛋白的调节结构域
(二序列特异性DNA结合蛋白的几种结构域
1•螺旋-转角-螺旋结构螺旋-转角-螺旋(helix-turn-helix )
2•锌指结构锌指(zinc finger)是由一小群氨基酸与一个锌原子结合,在蛋
白质中形成相对独立的一个结构域,故而得名.
3、亮氨酸拉链结构亮氨酸拉链(leucine zipper, ZIP)结构也是转录因子
DNA结合区的一种结构模式
4 •螺旋-环-螺旋结构螺旋-环-螺旋(helix-loop-helix,HLH )是新近发现的一种DNA结合区的结构模式
多细胞真核生物的一些基因表达常受体内外激素(hormo ne)的控制,
五、真核基因表达的激素调节
1、激素(hormone)的调控基因转录
(1)种类:甾类激素:
多肽激系
(2)甾体激素作用机制
甾体激素与受体蛋白结合,与靶基因DNA上激素应答成分结合,再和其他
因子协同作用来调控该基因的转录(如下图)。
六、Britten-Davidson模型
(一)Britten-Davidson 调节模型
在个体发育期,许多基因可被协同调控,且重复序列在调控中具有重要的作用。
(二)参与调控的遗传因子:
1、受体位点,位于结构基因5端,可被激活因子激活因子激活。
2、整合基因,产生激活因子的基因。
3、感受位点,接受生物体对基因表达调控的信号。
通过特定的激活因子可以同时控制不连锁但含用相应受体位点的多个结构基因协同表达。
含有相同受体位点的基因组成一组基因,类似原核生物的一个操纵子。
而整合基因类似于调节基因,但其转录受感受位点控制。
受体位点类似操纵基因,如果一个结构基因附近具有几个不同的受体位点,各个受体位点可以被特异的激活因子所识别,结构基因能在不同的情况下表达,也就是说一个结构基因可以属于几个不同的组(图 10-12B)。
如果一个感受位点可控制几个整合基因,则可同时产生几种激活因子,使不同组的基因也能同时被激活而进行协同表达。
(二)重复序列在协同调控中的作用
真核生物基因表达的协同调控是多级别,也是经济的调控方式,一种信号可以使不同的基因得到协同表达,其基础是整合基因、受体位点上具有重复序列。