水稻品种对黑条矮缩病和灰飞虱抗性的关联性分析
水稻品种抗水稻黑条矮缩病鉴定和介体飞虱体内病毒的定量测定

水稻品种抗水稻黑条矮缩病鉴定和介体飞虱体内病毒的定量测定水稻黑条矮缩病毒(RBSDV)属于呼肠孤病毒科,斐济病毒属,引起的水稻黑条矮缩病是东亚和东南亚地区最严重的水稻病害之一。
近年来该病害尤其在稻麦轮作区的发病率迅速上升,严重威胁水稻、小麦和玉米的安全生产。
灰飞虱(Laodelphax striatellus Fallen)和其他2种飞虱白脊飞虱(Unkanodes sapporona Matsumura)白带飞虱(U.albifascia Matsumura)均以持久增殖型方式传播RBSDV。
此外,由白背飞虱(Sogatella furcifera Horvath)传播的南方黑条矮缩病毒(SRBSDV)于2001年在中国广东省阳江市首次发现,已经传播蔓延到中国南部、越南和日本,成为杂交水稻的主要威胁。
本研究旨在通过筛选的水稻品种对RBSDV的抗病性,并探索不同介体飞虱体内病毒含量的时空动态。
在温室对收集自河南、山东和辽宁等地的66个水稻品种进行了抗RBSDV鉴定。
结果表明,两优3399、大粮207、X008、宁粳4号、圣稻301和两优1129等7个水稻品种表现抗病;沈农1406、D264、博377、中稻1号、春潮88、临香1号、圣稻15号、苏秀9号、苏秀10号、郑稻19、徐稻7号、津优9702、原稻108、苏秀326、原旱稻3号、X007和扬两优6号等16个水稻品种表现为中抗,其他品种表现为中感或高感。
大田抗病性鉴定与温室抗病性鉴定具有明显的正相关性(P=0.0003)。
另外,本研究成功利用实时定量PCR(q RT-PCR)Taq Man探针法检测定量了水稻中RBSDV的拷贝数,抗感水稻品种中RBSDV的拷贝数与其抗病性具有显著的相关性。
利用绝对定量q RT-PCR对整头介体昆虫或其不同器官进行定量检测RBSDV的拷贝数,结果表明,仅在灰飞虱中检测到RBSDV,整头灰飞虱中RBSDV的拷贝数随时间延长增多,说明了RBSDV在介体灰飞虱体内存在明显的复制。
不同生态型水稻品种对水稻黑条矮缩病的抗性反应及遗传研究

不同生态型水稻品种对水稻黑条矮缩病的抗性反应及遗传研究摘要2011年鲁南苏北稻区春季水稻灰飞虱大暴发且黑条矮缩病带毒率高。
对来自不同生态区域以及不同生育期的水稻品种进行田间自然鉴定,同时进行抗水稻黑条矮缩病遗传初步研究,以期筛选抗源类型,为抗水稻黑条矮缩病育种提供一些有价值的资料。
关键词水稻品种;黑条矮缩病;灰飞虱;抗性水稻黑条矮缩病(rice black-streaked dwarf viral disease,RBSD)是以灰飞虱(Laodelphax striatellus Fallen)为主要传播介质的病毒病,属于呼肠孤病毒科斐济病毒属(Fi-fivirus),具有暴发性[1-2]。
水稻一旦感染该病,则无法医治。
该病曾在20世纪60年代中期在江浙地区稻、麦、玉米等作物上严重发生,以后基本上销声匿迹。
近年来,由于耕作制度的改变,持续暖冬及越冬寄主增多导致带毒灰飞虱虫口密度加大,越冬地带北移[3]。
20世纪末在浙江开始蔓延,近几年在江苏由南到北不断发展,2008年造成较大危害,2009年受害面积逾33.33万hm2。
山东鲁南稻区2008年零星发生;2009年在济宁稻区大暴发,许多感病品种地块几近绝收。
目前,生产上没有品种抗黑条矮缩病,最优的为中感,如徐稻3号、连粳7号、苏秀867、大粮207等;2011年鲁南稻区水稻黑条矮缩病大面积暴发,济宁地区发生尤其严重,鱼台县很多田块几乎成为光板地。
针对这种情况,从2008年开始着手进行抗黑条矮缩病品种资源的征集、鉴定筛选,在抗黑条矮缩病品种鉴定的基础上进行杂交育种,进行遗传规律的分析研究,以期找到抗性较好的品种资源,为抗病育种服务。
1 材料与方法1.1 试验地概况试验于2011年在临沂市农科院水稻研究所试验田进行。
试验田周边为麦田。
1.2 试验材料临沂市农业科学院水稻研究品种资源及近10多年来东北稻区、西北稻区、华北稻区、黄淮稻区、江浙稻区育成的水稻品种和品系。
不同药剂配施对稻飞虱及水稻黑条矮缩病的防效试验

1 材料 与 方法
1 . 1 试验药 剂 7 0 %噻虫嗪种子处理可分散粉剂 ( 锐胜 ) ( 先 正达 中国投资有 限公 司) ; l O %p  ̄ 虫 啉 wP ( 江苏 克胜集 团有
限公司 ) ; 2 5 %毗蚜 酮 S C ( 江苏克胜集 团有 限公司 ) ; 4 0 %毒死 蜱E C( 浙 江新农化 工股份 有限公 司 ) ; 2 5 %噻 虫嗪 WG( 阿克
ห้องสมุดไป่ตู้
1 . 4 试 验地基本情 况
试 验选点在铜 山区房村镇 尚王村孟
缩病的效果 , 筛选最 佳防治方 案 , 特安排 了本次试验 。
建朋 责任 田中( 为上 年发病 田) , 该 田面积 3 0 0 0 m , 肥 力均 匀, 土壤类 型为粘壤 , 前茬 作物小麦 , p H值 6 . 3 。供试水稻 品 种为镇稻 9 9 , 6 月1 2 E l 各处 理区 同时播种 , 各试验 区均按照 试验设 计 实施用 药 防治 。收获期 测算平 均株 高 、 每穗 实粒
死蜱 E C 1 0 0 g / 6 6 7 m 。 喷雾 ; ( 5 ) 2 5 %吡 蚜酮 s c 2 o g 6 6 7 m 。 喷雾 ;
1 . 7 白背飞虱 及灰飞 虱发生 情况
背 飞虱的迁入 , 7 月8  ̄ 9日出现 白背飞虱迁 入小峰 , 空 白对 照 区于 7 月 9日调查 2 0 点, 平均虫量 2 . 6 头/ 0 . 1 l m , 7 月2 7日调
水稻黑 条矮 缩病 、 南 方水 稻黑条矮缩病分别是 由灰飞虱
每 处理 区面 积为 3 3 0 m : , 在各 区之间筑 泥梗 ( 2 0 ~ 3 0 e m) 以防
不同类型水稻品种对灰飞虱抗性的初步鉴定

供试材料 包括 7份 籼稻 品种 :R 6 I6 、 阳 5 、 阳 I3 、R 4 密 0密 4 、3 1 Y B 1 T T P 8份 粳 稻 品 种 : 稻 1 、 6 武 69 1 、 S R 、 E E ; 圣 3V 、
暴发成灾趋 势 。 目前 对水 稻灰 飞虱 的防治 主要依靠 化学
份恢复系: 明恢 6 、 9 8 C 3 广恢 9 8 1 杂交籼稻 : 3R 9、 5、 9 ;份 汉盐 优 1 号。为聚合 灰飞 虱抗 性基 因 ( T  ̄ 、 Q L ) 培育 水稻 抗性 品 种, 本研究配制了“ 圣稻 15 I3 ” 0 /R 6 组合 , 供试材料 还包括来 自该组合的 6 6份 F. 5 5高代株系。
理利用抗源及聚合 多种抗性 基因、 选育水 稻灰飞虱抗性 品种
奠定 基 础 。 1 材 料 与 方法
1 1 供 试材 料 .
生产上 的一类重要害虫 。水稻 经灰飞虱取食后 , 不仅植 株黄
叶、 早枯萎缩甚至霉烂枯死 , 或稻穗发 黑霉变 , 直接 影响灌浆
结实 , 造成空秕率上升 、 千粒重下降 、 品质降低 。另外 , 稻米 灰 飞虱不仅传播水稻条纹叶枯病 , 也是黑条 矮缩病 的主要 传毒
一
1O一 1
江苏农业科 学 2 1 0 2年第 4 O卷第 7期
朱文银, 朱其 松, 张洪瑞 , 等.不同类型水稻 品种对灰飞虱抗 性的初步鉴定[ ] J .江苏农业科 学,0 2 4 7 :1 2 1 ,0( ) 1 0一ii i
不 同类 型水 稻 品种 对灰 飞 虱抗 性 的初 步 鉴 定
15 I3 ” 0 /R 6 组合的 F. 5 代株系材料的灰飞虱抗 性进行 了初步 鉴定。筛 选 出对灰 飞虱具有不 同程度抗性 的水稻 品种 1 0
水稻播期灰飞虱虫量与黑条矮缩病的发病关系

收稿 1期 :0 0 0 — 4 3 2 1— 10
近 年 施 用 、 治 效 果 和 农 药 特 性 的 比较 , 度 核 定 克 胜 集 团 防 再
等 企 业 生 产 的名 牌 新 品 为 “ 海 市 2 1 上 0 0年 度 农 药 重 点 推 荐 品种”启动 21 . 0 0年 病 虫 害 防治 补 贴 药剂 前 期 工 作 。克 胜 集
2 结 果 与 分 析
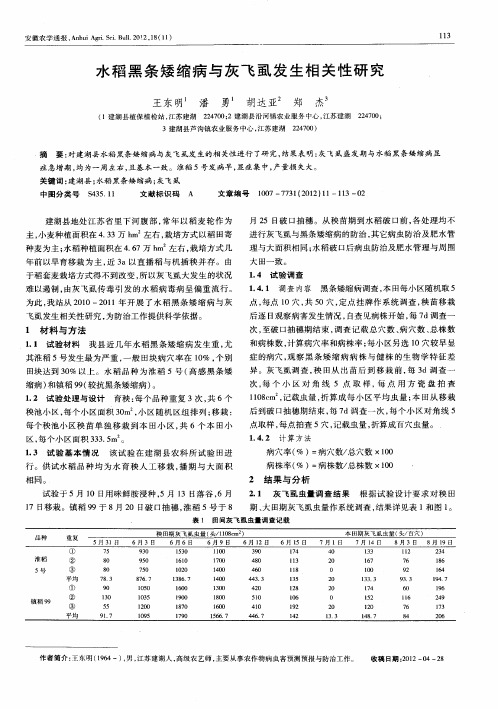
2 1 水 稻 播 种期 与 灰 飞 虱 虫量 的 关 系 .
落 谷 的平 均 病 穴 率 分 别 为 8 、%和 43 % . 均 病 株 率 分 % 6 .3 平
别 为 28 % 、 .%和 13 .与 5月 1 播 种 的 处 理 区 相 . 7 1 8 .% 01 3
穴数 、 株 数 . 计 病 穴 率 和病 株率 , 病 情稳 定 时 的 发 病株 病 统 将
率作 为 本 田发病 率 。
大 田 始查 见 病 株 , 7月 上 中 旬 显 症 加 速 , 旬 基 本 稳 定 。 验 下 试
结果 显示 ( 表 1 , 见 ) 5月 l 日 、 0 5月 2 、 0 1 5月 3 三 期 3 01 3
分析各播期灰 飞虱虫量与黑 条矮缩病发病 率关系可 以 看 出 . 上旬 , 田期 灰 飞 虱 虫 量 较 大 . 致 黑 条 矮 缩 病 发 5月 秧 导 病 较 重 . 随播 种 时 间 的 推 迟 . 而 田间 虫 量 则 明 显 减 少 . 害发 病 生 也 明 显减 轻 通 过对 各 播 期 秧 田灰 飞 虱 虫量 和发 病 率 进行
试验结 果显 示 , 种期 不 同 , 田期 虫量差 异很 大 。5月 1 播 秧 0 1 5月 2 和 5 3 落 谷的 , 田灰 飞虱虫 量每 67m 分 3、 01 3 月 01 3 秧 6
水稻黑条矮缩病与灰飞虱发生相关性研究
14 1
安徽 农 学通 报 , n u A . c B l 2 1 ,8 1 ) A hi Si u1 0 2 1 ( 1 . .
生, 叶枕 多有 重叠或 错位 , 穴表 现矮 缩 , 株 高度 只有 正 整 植
常植株 的 12左右 。病 苗 中大部分 为 整穴 发病 , / 也有 少 部
14 1 调查 内容 ..
黑条矮 缩病 调查 , 田每 小区 随机取 5 本
点, 点 1 每 0穴 , 5 共 0穴 , 定点 挂 牌作 系统 调 查 , 苗 移 栽 秧 后逐 日观察病 害发 生情 况 , 自查 见病株 开 始 , 7 每 d调查 一 次 , 破 口抽穗期 结 束 , 至 调查 记 载 总 穴数 、 穴数 、 株 数 病 总
2 球
V
+ 淮稻 5 号 一一 镇稻9 9
分为病 健混 生 。发病植 株 叶部病 症 , 开始 表 现为 叶基 部 皱 缩 , 分病 株心 叶上 有 黄 绿条 斑 , 背 叶脉 处 有 短 线 腊 白 部 叶
秧池 小 区 , 每个 小 区面积 3 m , 区随 机 区组 排 列 ; 栽 : 0 小 移
后 到破 口抽穗 期结 束 , 7 每 d调查 一 次 , 个 小 区对 角 线 5 每
点取 样 , 点拍查 5穴 , 载虫 量 , 每 记 折算 成百 穴虫量 。
1 4 2 计 算方法 ..
中图 分 类 号 ¥3.1 45 1 文 献标 识 码 A 文章 编 号 10 0 7—73 (0 2 1 一l3— 2 71 21 )1 1 0
建 湖县地 处 江 苏 省 里下 河 腹 部 , 年 以 稻麦 轮作 为 常
主 , 麦种 植面积 在 4 3 小 . 3万 h 左右 , 培方 式 以稻 田寄 m 栽 种麦 为 主 ; 水稻种 植 面积在 4 6 .7万 h 左 右 , 培方 式几 m 栽 年前 以旱 育 移栽 为 主 , 3 近 a以直 播 稻 与 机 插秧 并 存 。由 于 稻套 麦栽 培方式 得不 到改 变 , 以灰 飞虱大 发 生 的状 况 所
我国水稻黑条矮缩病及其防治研究进展

分也可在灰飞虱体内越冬 。而南方水稻黑条矮缩病 则是以白背飞虱传毒为主, 其寄主范 围与已知斐 济 病毒 存在一 定 的差异 [ 1。 5 J
灰 飞虱属 温 带区 害虫 , 我国分 布广泛 , 在 以华北
冬麦 区和长江 中下 游 稻 区发 生 较 多 , 在华 南仅 在 早
稻 上和较 高海 拔地 区发 生 。其 发 生世代 受地理 和气
成 lJ 认 为 高抗 黑 条 矮 缩 病 水 稻 品种 有 “ 稻 2 等则 0 泗 1” “ 稻 1” “ 2 、郑 8 、 盐粳 9号 ” “ 辐粳 8号 ” “ 连 、扬 、武 粳 1 ”“ 号 、镇稻 9 ” 9。
4 1 2 改进栽 培制 度 .. 积极推 广旱 育秧 或半 干 旱 育秧 , 田尽 量 连 片 秧
系为抗病 型 , 通过 籼 粳 杂交 培 育 的粳 稻 恢 复 系基 本
留下的空穴里, 然后适当加施速效肥 , 促进稻苗生长, 通过杂交稻分蘖势较强 的特点来减少产量损失 。 4 2 化 学 防治 . 药 剂 防治 是 目前 防治 水稻 黑条矮缩 病 的主要手 段, 选用有效农药 , 注重高效与速效相统一, 当增 适 加 防治 密度 。具 体 策略是 : 治秧 田、 大 田、 保 治前期 、 保后期 I 5。
我国水稻黑 条 矮缩 病 (i l ks ekdd r r eba —ra e waf c c t
d e e最早于 16 年在浙江省余姚 等县发 现, ia ) ss 93 同 时在上海市嘉定和奉 贤县 、 江苏省苏州和镇江等专
区 的水稻 上 、 扬州 和 南 通 等专 区 的玉 米 上有 局 部 危 害[ 3。水稻黑 条矮 缩病是 主要 由灰 飞虱 [ adl IJ - L oe一
南方水稻黑条矮缩病苗期抗性基因的关联分析、精细定位及候选基因分析

南方水稻黑条矮缩病苗期抗性基因的关联分析、精细定位及候选基因分析水稻(Oryza sativa L.)是世界上最重要的粮食作物之一,全球有超过一半的人口以稻米为主食,南方水稻黑条矮缩病(SRBSDV)的爆发给水稻生产造成极大损失。
实践证明,发掘利用新抗源、培育抗病新品种,是控制水稻病害最经济有效、最环保的措施。
然而,迄今为止,尚无与SRBSDV抗性相关的基因或QTL的报道。
本研究利用SNP标记,对广西地方稻种资源核心种质的SRBSDV苗期抗性进行全基因组关联分析。
同时对普通野生稻导入系D4的SRBSDV苗期抗性主效QTL进行精细定位,结合基因表达量分析鉴定出候选基因,为抗SRBSDV水稻分子育种提供新的基因资源和可靠的分子标记,为该基因的图位克隆和功能分析奠定坚实的基础。
1.广西地方稻种资源核心种质SRBSDV苗期抗性的全基因组关联分析利用人工接种方法,对广西地方稻种资源核心种质进行SRBSDV苗期抗性鉴定,结果显示,419个地方品种的SRBSDV苗期抗性表现出明显差异,发病率变异范围为6.11-100%,平均值为84.80%。
利用211,818个SNP标记,全基因组关联分析(GWAS)共检测到5个与水稻SRBSDV苗期抗性相关的QTL,分别为qSRBSDV1、qSRBSDV4、qSRBSDV5、qSRBSDV11及qSRBSDV12,单个QTL可解释7.84-17.80%的表型变异。
在所有的关联SNP位点中,共检测到221个候选基因,其中46个候选基因在籼稻群体和总体样本群体中均被检测到。
2.普通野生稻导入系D4的SRBSDV的抗性特征及其遗传分析D4为含有普通野生稻血缘的高抗SRBSDV材料,本研究采用排驱性及抗生性测验分析了 D4对传毒介体白背飞虱的抗性水平,结果表明D4对传毒介体白背飞虱既无排驱性也无抗生性,表明其对SRBSDV的抗性为抗病毒性而非抗虫性。
同时,采用人工接种鉴定方法对广恢998/D4 F2代进行SRBSDV苗期抗性遗传分析,结果显示,F2群体的水稻SRBSDV苗期抗性呈偏正态分布,表明抗源D4的SRBSDV苗期抗性受主效基因和微效基因共同控制。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
水稻品种对黑条矮缩病和灰飞虱抗性的关联性分析打开文本图片集摘要:为分析水稻品种对黑条矮缩病和灰飞虱抗性的关联性,对49个品种进行田间接种鉴定、人工接种鉴定、抗生性和排驱性试验。
结果表明,田间接种中发病率低于20%的品种较多,田间接种鉴定受到外部环境影响因素较大,发病率明显低于人工接种鉴定。
在田间接种和人工接种鉴定中,籼稻品种的发病率低于粳稻品种。
抗生性试验显示,仅5个籼稻品种的抗生性值低于81%,粳稻品种均感虫。
籼稻品种排驱性值明显低于粳稻品种,籼稻品种对灰飞虱的抗性高于粳稻品种。
对水稻品种的黑条矮缩病抗病性和灰飞虱抗虫性进行关联性分析结果,田间鉴定中品种发病率与对灰飞虱排驱性呈极显著正相关,而与品种对灰飞虱抗生性没有相关性。
人工接种鉴定中品种发病率与品种对灰飞虱的抗生性和排驱性都呈显著正相关。
排驱性与田间接种鉴定发病率和人工接种鉴定发病率均呈极显著正相关。
表明在灰飞虱传毒过程中,抗虫品种在一定程度上影响了传毒效果,培育抗灰飞虱品种在一定程度上可以减轻水稻黑条矮缩病的危害。
关键词:水稻;黑条矮缩病;灰飞虱;关联性收稿日期:2022-04-15目前,国内部分学者已开展水稻黑条矮缩病和灰飞虱遗传基础研究,发现了一批抗灰飞虱水稻品种,如RH、Mudgo、ASD7、IR36、IR64等,并定位到近20个抗灰飞虱QTL[4-9]。
王宝祥等开展了抗黑条矮缩病水稻种质资源筛选和相关基因定位工作,筛选到了多个抗病品种,如来自东南亚的籼稻品种Kanyakumari29、Madurai25和Vietnam160等[10]。
潘存红等利用珍汕97B/明恢63的重组自交系群体,对水稻黑条矮缩病抗性QTL进行分析,共检测到6个QTL,并对其中1个进行了精细定位,这些位点的抗性等位基因来自抗病亲本明恢63[11-12]。
王英利用Tetep/淮稻5号构建分子连锁图谱,共检测到4个抗性QTL[13],但通过重复鉴定表明,明恢63和Tetep的发病率可达20%以上,高抗种质资源的发掘和相关抗病基因的定位仍较为迫切。
水稻黑条矮缩病暴发的严重程度也与携带RBSDV的灰飞虱种群密度密切相关[14-15],有效控制灰飞虱种群可以间接防治由其传播的病毒病,因此,研究水稻品种对黑条矮缩病的抗病性和对灰飞虱的抗虫性二者之间的关联性显得尤为重要。
本研究分别开展了水稻品种对黑条矮缩病抗病性及传毒介体灰飞虱抗虫性试验,并对这2种抗性之间的关联性进行了分析,以期为阐明水稻黑条矮缩病抗性构成机制和寻求新的抗性途径解决生产实际问题提供理论依据。
1材料与方法1.1供试材料水稻品种共计49个,其中籼稻品种19个,粳稻品种30个(表1)。
1.2试验方法1.2.1黑条矮缩病田间自然接种鉴定材料种植于东海县黄川试验田,选用四周为麦田的地块种植待鉴定的材料,以保证获得足量虫源。
2022年5月15日(麦收前3周)播种,每个品种播80粒,均匀撒播1行,行长50cm,行距10cm。
6月20—25日移栽,单苗栽插,株距15cm,行距20cm,每个品种和家系栽插40穴。
常规肥水管理,不施用任何农药。
通过盘拍法调查灰飞虱虫口密度[10],试验田块秧苗期的灰飞虱为3048万头/hm2,虫口基数巨大。
水稻分蘖盛期统计发病率。
此时期发病植株表现出极明显的矮缩,心叶突破下叶叶鞘而出,部分植株表现为从下叶枕口呈螺旋状伸出,部分叶片的顶端出现旋状卷曲;而健康稻株分蘖旺盛,生命力强,但尚未拔节,发病植株和健康植株反差非常明显。
以2个重复发病百分率平均值作为品种抗黑条矮缩病表型值进行统计。
将亲本、抗病和感病对照品种浸种催芽,将待鉴定品种播种于长70.5cm、宽50.5cm、高41.5cm的塑料箱内,每个品种播种50粒,呈“一”字形,保持水层约3cm。
秧苗长至2.5~3.0叶时间苗,淘汰病弱苗,保留约40株整齐一致的健苗,塑料箱顶端用纱布封口。
以2个重复发病百分率平均值作为品种抗黑条矮缩病表型值进行统计。
1.2.3排驱性试验参照Nemoto等的方法[16]并适当改进:催芽后的水稻种子播于长65cm、宽44cm、高14cm的塑料周转箱内,每个品种1行,每行25株。
随机排列,重复2次。
幼苗长至1.5~2.0叶,剔除弱小苗,每行留苗15株,按每株5头接入2~3龄灰飞虱若虫,自然光照,室温(26±1)℃。
24h后调查每个单株上的虫数,每天1次,调查后驱虫,使其尽量均匀分布。
5d后计算每个品种单株上的平均虫数,作为排驱性测验值。
1.2.4抗生性试验参考Duan等的方法[6],单管单苗(种子萌发后第8天的幼苗)接入5头1~2龄的灰飞虱若虫,接虫后6h开始统计死虫数,计算若虫存活率,每天统计1次,连续5d。
抗生性分数(antibioicore,AS)计算公式为:AS=[(A1某1)+(A2某2)+…+(An某n)]某100%/(1+2+…+n)。
式中:n为接虫后的天数,An为该天灰飞虱若虫存活率。
每个试验品种测定10株幼苗。
AS值在0~81%的定为抗虫,AS>81%的品种为感虫,AS值越小,其抗生性抗性越强,反之则弱。
试验温度(25±1)℃。
1.3灰飞虱带毒率检测参照周彤等的方法[17]随机对100头灰飞虱进行水稻黑条矮缩病毒的RT-PCR检测,结果显示黑条矮缩病毒的携毒率平均达到5%,具备了足量毒源。
1.4数据分析利用SPSS软件对不同组试验结果进行分析。
2结果与分析2.1不同水稻品种对黑条矮缩病及灰飞虱抗性的分析对49个水稻品种进行抗黑条矮缩病田间接种和人工接种鉴定表明,人工接种鉴定发病率高于田间接种发病率,仅1个品种的发病率低于20%(RH)。
而田间接种中发病率低于20%的品种较多,与人工接种鉴定表型值相差较大,说明田间接种鉴定受到外部环境影响因素较大(表1、图1-A、图1-B),发病率明显低于人工接种鉴定。
在田间接种和人工接种鉴定中,籼稻品种的发病率低于粳稻品种(表1、图1-A、图1-B)。
水稻品种对灰飞虱抗生性试验结果,仅5个籼稻品种的抗生性值低于81%,粳稻品种抗生性值均高于81%表现为感虫;籼稻品种排驱性值明显低于粳稻品种(表1、图1-C、图1-D),表明籼稻品种对灰飞虱的抗性高于粳稻品种。
利用SPSS软件对水稻品种的黑条矮缩病抗病性和灰飞虱抗虫性进行关联性分析,田间鉴定品种发病率与对灰飞虱排驱性呈极显著正相关,而与品种对灰飞虱抗生性不呈相关,表明田间接种易受品种对灰飞虱排驱性的影响;另外田间接种鉴定发病率与人工接种鉴定发病率呈显著正相关,表明2种鉴定方法结果具有一致性。
人工接种鉴定品种发病率与其对灰飞虱的抗生性和排驱性都呈显著正相关,因此不同水稻品种对灰飞虱的排驱性和抗生性均影响人工鉴定结果。
另外,排驱性与田间接种鉴定发病率和人工接种鉴定发病率均为极显著正相关,表明水稻品种对灰飞虱排驱性是影响鉴定结果的重要因素(表2)。
3讨论传毒介体、病毒和植物三者之间的互作导致了病毒病害的发生,相关对水稻黑条矮缩病抗性构成机制的研究报道较少,但在其他病毒病害研究中已展开相关研究。
日本学者Nemoto等对由灰飞虱传播的水稻条纹叶枯病研究发现,中抗品种IR50和Tadukan对病毒和介体昆虫都有抗性,提出水稻对昆虫的抗性可减少条纹叶枯病毒传播,水稻对条纹叶枯病的抗性包含对水稻条纹叶枯病毒的抗性和对介体灰飞虱的抗性两层含义,对病毒的抗性又可分为抗病毒侵染和对病毒的忍耐性[16]。
另外,Kobayahi等[18]和Takita等[19]分别在1983年和1986年的研究中发现水稻品种IR42对东格鲁病毒的抗性主要由对介体昆虫的抗性所致。
本研究利用49个不同地区和类型水稻品种进行田间接种鉴定、人工接种鉴定、抗生性和排驱性试验,在田间接种和人工接种鉴定中,籼稻品种的发病率低于粳稻品种;在品种抗生性和排驱性试验中,相对粳稻品种,籼稻品种对灰飞虱的抗性较强。
因此,本研究中籼、粳2种类型水稻品种对黑条矮缩病和传毒介体灰飞虱的抗性存在一定关联性。
品种田间鉴定发病率和人工鉴定发病率与抗生性值和排驱性值之间的相关性分析结果表明,品种发病率与品种对灰飞虱排驱性呈极显著正相关,在田间鉴定中品种发病率与品种对灰飞虱抗生性不相关,说明田间接种易受品种对灰飞虱排驱性的影响,因此品种对灰飞虱的排驱性是影响田间鉴定结果的重要因素。
在人工接种鉴定中品种发病率与其对灰飞虱的抗生性和排驱性均表现为显著正相关,品种对灰飞虱的排驱性和抗生性均影响人工鉴定结果。
因此,在灰飞虱传毒过程中,抗虫品种在一定程度上影响了传毒效果,培育抗灰飞虱品种在一定程度上可以降低水稻黑条矮缩病的发生[20-23]。
籼、粳稻对相同病毒病害往往表现出不同的抗性反应。
Yamaguchi等提出条纹叶枯病毒的抗源广泛存在于籼稻、爪哇稻和日本的旱稻中,而其他粳稻品种中不存在抗源[24-27]。
本研究结果表明,在田间接种和人工接种鉴定中,籼稻品种的发病率低于粳稻品种,籼稻品种对灰飞虱的抗性高于粳稻品种,结果表明,籼稻品种对黑条矮缩病抗性强于粳稻品种,可能是由于籼稻品种较抗传毒介体灰飞虱的缘故。
本研究结果为阐明水稻品种对黑条矮缩病和灰飞虱抗性的关联性提供了理论基础。