第三节 心肌的生理
骨骼肌、心肌和平滑肌细胞生理

(一)收缩机制 1、4个Ca2+与钙调蛋白结合形成复合体 2、钙调蛋白复合体与肌球蛋白轻链激酶
(MLCK)结合(激活) 3、激活的MLCK利用ATP肌球蛋白轻链磷酸化 4、磷酸化的横桥被激活,与激动蛋白结合
(二)钙离子浓度的调节
细胞外Ca2+是启动平滑肌收缩的主要来源
(二)根据结合ATP酶的类型来分 氧化型:线粒体多,氧化磷酸化能力强 红肌纤维 糖解型:线粒体少,糖解酶和糖原贮存多 白肌纤维
三种类型的纤维
慢氧化肌纤维(Ⅰ) 快氧化肌纤维(Ⅱa) 快糖解肌纤维(Ⅱb)
第二节 平滑肌生理
非横纹肌、不随意肌
平滑肌(纵切)
平滑肌(横切)
一、平滑肌的结构
1.呈长梭形,直径不等,无分支。 2.单核,椭圆形或杆状,居中。 3.无横纹。
平滑肌纤维的超微结构特点
1、无横小管,可见肌膜凹 陷形成的小凹;
2、肌纤维内无肌原纤维, 可见致密体、中间丝、 粗肌丝和细肌丝;
3、相邻肌纤维之间有缝隙 连接。
二、平滑肌的电活动
平滑肌动作电位的发生以来于Ca2+,而不是 Na+,除极化开放的电压门控的Ca2+通道。
(三) 骨骼肌舒张机制
兴奋-收缩耦联后
肌膜 肌浆网膜Ca2+泵激活
肌浆网膜[Ca2+]↓
原肌凝蛋白复盖的 横桥结合位点
Ca2+与肌钙蛋白解离
骨骼肌舒张
三、骨骼肌收缩的机械特性
负荷:牵拉肌肉的力
张力:肌肉收缩时可对接触物体 产生的力
两相反的力
肌肉收缩表现为长度的缩短和张力的增加。
细肌丝:肌动蛋白:表面有与
动物生理学心肌的生理特点-概述说明以及解释

动物生理学心肌的生理特点-概述说明以及解释1.引言1.1 概述概述部分的内容可以是对动物生理学心肌的定义和背景进行介绍,可以包括以下内容:心肌是构成心脏的一种特化肌肉组织,其在维持心脏正常运转中发挥着关键的作用。
心肌组织具有独特的生理特点,通过有效的收缩和舒张过程,实现了心脏的收缩和血液的泵送。
了解心肌的生理特点对于理解心脏的功能和研究心脏疾病具有重要意义。
心肌的生理特点主要包括其组成与结构以及收缩机制。
心肌是由心肌细胞组成的,这些细胞具有丰富的线粒体、细微管和肌纤维等结构,使其能够快速而协调地收缩。
心肌细胞相互连接,形成有序的心肌组织,保证了心脏的整体收缩和血液的流动。
心肌的收缩机制是心肌细胞发生收缩的过程,涉及多种离子通道、肌纤维的交互作用和能量的耗散。
在收缩过程中,钙离子的释放和收回起着重要的调控作用。
在一系列信号传导的调节下,心肌细胞发生有序的收缩,从而推动血液通过心脏流入全身循环。
了解心肌的生理特点对于心脏疾病的预防、诊断和治疗具有重要意义。
心肌病、心肌缺血等心脏疾病常常涉及心肌的异常结构和功能,而了解心肌的生理特点可以为研究这些疾病的发病机制提供基础。
此外,对心肌的生理特点的深入研究还有助于寻找心肌保护和心肌再生的新途径,为心脏病的治疗提供创新思路。
综上所述,动物生理学心肌具有独特的组成与结构以及收缩机制,了解心肌的生理特点对于理解心脏功能和研究心脏疾病具有重要意义。
在接下来的正文中,将详细介绍心肌的组成与结构以及心肌的收缩机制等内容。
1.2文章结构文章结构部分的内容可以根据所要讲述的内容来进行编写,以下是一个可能的内容示例:2. 正文2.1 心肌的组成与结构本节将介绍心肌的组成和结构,以便更好地理解心肌的生理特点。
心肌是心脏的重要组成部分,由一种特殊的肌肉组织构成,它与其他肌肉组织有着显著的区别。
在本节中,将详细阐述心肌的结构组成和细胞特点,并介绍心肌与其他组织的相互作用。
2.2 心肌的收缩机制本节将探讨心肌的收缩机制,这是心肌的重要生理特点之一。
心脏生理

第三节心脏生理一、选择题:1.正常心动周期中,下述哪项是正确的?()A.心房收缩期处在心室舒张期内B.心房舒张期处在心室收缩期内C.心室收缩期处在心房收缩期内D心房舒张期处在心室舒张期内2.心脏的正常起搏点是()A.窦房结B.静脉窦C.房室结D. 浦肯野纤维3.在心动周期中房室瓣关闭而动脉瓣开放是哪一期? ()A.心房收缩期B.等容收缩期C.射血期D.等容舒张期4.正常人安静时每博输出量()A.等于每分输出量与心率的乘积B.左心室每博输出量明显大于右心室C.指1次心跳两侧心室射出的血量D为一侧心室每收缩一次所射出的血量5.评定心脏泵血功能的指标有()A.心输出量B心率C心音D心电图6.心肌细胞在超常期的特点有()A没有兴奋性B兴奋性高于正常C兴奋性低于正常D产生动作电位幅度大于正常7.在正常的心动周期中,不可能发生的是.()A心房收缩时,心室处于收缩期B心房收缩时,心室处于舒张期C心房舒张时,心室处于收缩期D心房舒张时,心室处于舒张期二、判断题:1.在心肌细胞有效不应期内,给予无论多强的刺激,也不会使心肌细胞产生动作电位。
()2.心率增大,则心输出量增加。
( )3.自律细胞和非自律细胞生物电现象的主要区别是4期自动去极化。
()4.心房肌和心室肌属于工作细胞。
()三、填空题:1.心脏传导性的特点是___—慢—____,其中“慢”是指冲动从心房传向心室时在房室交接处存在_________现象,其生理意义在于使心室的收缩发生在心房收缩之___________。
2.心室肌细胞动作电位2期形成的机制是由于______内流和______外流接近于平衡状态的结果。
3.心肌细胞不发生强直收缩的生理意义包括:心脏总是保持着______和______交替的节律性活动,保证泵血功能的正常完成。
4. 根据各类心肌细胞的生物电特征,可将心肌细胞分为___________细胞和__________细胞。
五、名词解释:1. 心动周期2. 心力贮备3. 心电图4.有效不应期5.自动节律性6.每博输出量7.心输出量六、问答题:1.何谓心动周期?在一个心动周期中,心房和心室的活动是怎样的?心率增加对心动周期有何影响?2.什么是正常起搏点、潜在起搏点和异位起搏点?3.什么是心电图?心电图各波所代表的意义是什么?。
《心肌的生理特性》演示PPT

时间短 时间长
-60
↓↓
自律性高 自律性低
0.1 0.2 0.3 0.4 0.5 0.6 0.7
时间(s)
10
⑵4期自动除极的速度
若自动除极速度
从最大舒张电位到达阈 电位所需的时间缩短
单位时间内自动兴奋发 生的次数
自律性
儿茶酚胺可加速窦房结细
反之,4期自动除极速度 胞4期自动去极化速度,
缓慢,则使自律降低。 提高自律性,使心率 。
‖
‖
‖
‖
兴奋性正常 兴奋性=0
兴奋性低 兴奋性高
20
心肌兴奋时兴奋性变化的主要特点是有效不 应期特别长(平均250ms),相当于心肌整个收缩期 和舒张早期。
它 是 骨 骼 肌 与 神 经 纤 维 有 效 不 应 期 的 100 倍 和 200倍。
这一特性是保证心肌能收缩和舒张交替进行,不 出现强直收缩的生理学基础。
大部复活 Na+通道基本 恢复到备用状态
不能产生 仅能产生 局部电位 阈上刺激
阈下刺激
14
1 兴 奋 性 的 周 期 性 变 化
15
2、影响兴奋性的因素
(1)静息电位或最大复极电位的水平 (2)阈电位的水平 (3)引起0期去极化的离子通道性状
16
⑴静息电位或最大复极电位的水平
17
⑵阈电位的水平
4
2.窦房结对潜在起搏点的控制
①抢பைடு நூலகம்占领 也称夺获。 在潜在起搏点4期自动去极化尚未达到阈电位水平之前,已 被自律性最高的窦房结传来的兴奋抢先激动,使之产生与窦 房结节律相一致的动作电位,从而使潜在起搏点自身的节律 兴奋不能出现。
②超驱动阻抑 窦房结的快速节律活动,对潜在起搏点较低 频率的兴奋有直接抑制作用,称为超驱动阻抑。当窦房结停 止发放冲动或下传受阻后,则首先由自律性相对较高、受超 驱动阻抑较轻的房室交界来替代,而不是由自律性更低的心 室传导组织来替代。人工起搏器。
心肌生理特性
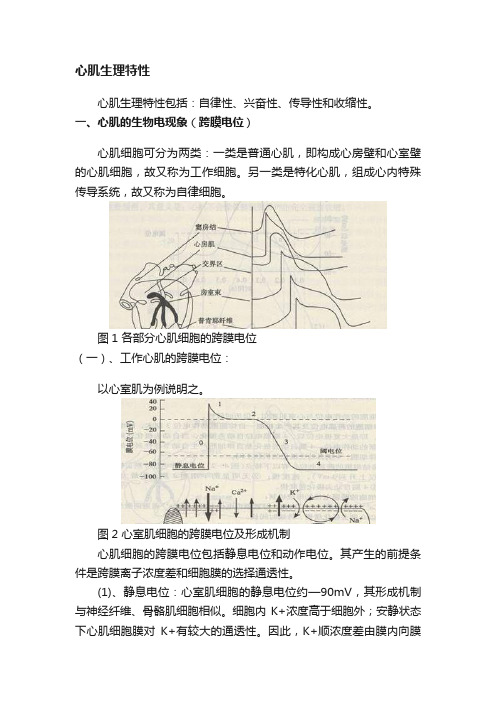
心肌生理特性心肌生理特性包括:自律性、兴奋性、传导性和收缩性。
一、心肌的生物电现象(跨膜电位)心肌细胞可分为两类:一类是普通心肌,即构成心房壁和心室壁的心肌细胞,故又称为工作细胞。
另一类是特化心肌,组成心内特殊传导系统,故又称为自律细胞。
图1 各部分心肌细胞的跨膜电位(一)、工作心肌的跨膜电位:以心室肌为例说明之。
图2 心室肌细胞的跨膜电位及形成机制心肌细胞的跨膜电位包括静息电位和动作电位。
其产生的前提条件是跨膜离子浓度差和细胞膜的选择通透性。
(1)、静息电位:心室肌细胞的静息电位约—90mV,其形成机制与神经纤维、骨骼肌细胞相似。
细胞内K+浓度高于细胞外;安静状态下心肌细胞膜对K+有较大的通透性。
因此,K+顺浓度差由膜内向膜外扩散,达到K+的电一化学平衡电位。
(2)、动作电位:心室肌细胞的动作电位分为0、1、2、3、4五个时期1、去极化:又称为0期。
在适宜刺激作用下,心肌发生兴奋时,膜内电位由原来的一90 mV上升到+30 mV左右,形成动作电位的上升支。
0期历时1~2 ms。
其产生机制:刺激使膜去极化达到阈电位(一70mV)时,大量Na+通道开放,Na+快速内流,使膜内电位急剧上升,达到Na+的电一化学平衡电位。
2、复极化:包括l期、2期、3期、4期。
1期:膜内电位由原来的+30 mV迅速下降到O mV左右,此期历时1 O ms 此期形成的原因主要是K+外流。
2期:1期结束膜内电位达O mV左右后,膜电位基本停滞在此水平达1 00~1 50 ms。
记录的动作电位曲线呈平台状,故此期称为平台期。
2期的形成主要是由Ca2+内流与K+外流同时存在,二者对膜电位的影响相互抵消。
3期:膜内电位由0MV 左右下降到-90 ,3期是Ca2+内流停止,K+外流逐渐增强所致。
4期:此期膜电位稳定于静息电位,所以也称静息期。
4期跨膜离子流较活跃,主要通过离子泵的活动,以恢复兴奋前细胞内外离子分布状态,保证心肌细胞的兴奋性。
心肌的生理特性通用课件

β受体拮抗剂
普萘洛尔 美托洛尔 阿替洛尔
钙通道阻滞剂
维拉帕米 地尔硫䓬
抗心律失常药物
利多卡因
利多卡因是一种局部麻醉药和抗心律失常药,主要用于治疗室性心律失常,可抑 制心肌收缩和传导神经,减慢心率。
胺碘酮
胺碘酮是一种广谱抗心律失常药物,可用于治疗各种心律失常,如房性、室性和 交界性心律失常等,可抑制心肌收缩和传导神经,减慢心率。
适量的有氧运动有助于提食有助于控制体重,预防心血管疾病。
戒烟限酒
戒烟有助于预防心血管疾病,限制酒精摄入对心脏健康也有益。
预防心血管疾病
控制血压
01
控制血脂
02
控制血糖
03
治疗心肌疾病的方法
药物治疗
介入治疗
手术治疗
THANK YOU
03
心肌的生物化学特性
心肌的能量代 谢
01
02
03
04
心肌的物质代谢
心肌的荷尔蒙调节
04
心肌的病理生理特性
心肌缺血与缺氧
心肌缺血 心肌缺氧
心肌肥厚与扩张
心肌肥厚
心肌扩张
心肌扩张是指心肌细胞体积增大,但 数目不变,导致心室腔扩大,心脏体 积增大。
心肌炎与心肌病
心肌炎
心肌病
05
心肌的药物治疗
心肌的生理特性通用件
• 心肌的生物化学特性 • 心肌的病理生理特性 • 心肌的药物治疗 • 心肌疾病的预防与治疗
01
心肌的概述
心肌的细胞构成
心肌细胞
工作细胞 自律细胞
心肌的功能特点
01
02
收缩性
节律性
03 传导性
心肌的电生理特性
01
心肌的生理特征
心肌的生理特征
心肌的生理特征:
1、形态特征:
a. 构造:心肌细胞由残余质原杆质蛋白和三种肌肉纤维组成:心肌细胞,肌膜细胞和胞质外分泌物质。
b. 大小:心肌细胞的大小在10~20微米之间,长度为50~200微米。
2、收缩特征:
a. 心肌有自发性收缩和被动性收缩两种收缩特性,自发性收缩能对外界刺激作出反应;而被动性收缩则可吸收紧缩力及回复缩短力,可把肌腱扭紧和释放收缩力。
b. 梗阻性心肌可以形成持久性收缩(Stasket's Law),是一种受控的、持续发生的收缩。
3、电性特征:
a. 心肌细胞的膜上有一种特殊的电导体,可以通过外源电刺激形成极化。
b. 心肌细胞有两个膜电位:安体膜电位和心肌收缩膜电位,两者之差就是心肌内电压。
4、代谢特征:
a. 心肌的代谢特点主要体现在能量生成和分子消耗上,其能量主要是来自糖酵解,氧消耗也比其他肌肉高。
b. 心细胞可以将6脂肪酸中的2个脂肪酸分解,也可以将肝胆糖原代谢为水解葡萄糖,可以调节收缩周期。
5、合成功能:
a. 心肌细胞具有酶系统,能够产生钠、钙离子和磷脂质,合成许多蛋白质,如RNA及DNA合成、细胞色素c合成以及胆碱选择性接受蛋白的合成等。
b. 它还可以分泌一种类似胰岛素的肽激素,可以促进脂肪酸、糖酵解,增强心脏合成蛋白的能力,调节心率及心室收缩力量等。
(最新整理)第三节离子与心肌
②细胞外Na+促进跨膜Na+-Ca2+交换,使细胞内液Ca2+外运增多。 这两者均使[Ca2+]降低,Ca2+激发心肌兴奋-收缩耦联作用减弱。 因此高血钾时心肌的收缩性降低,收缩力减弱。
2021/7/26
8
3、心电图的变化
高钾时,由于APA和Vmax降低,心电图中显示心房的P波 电压降低和波幅增宽,心室的R波电压低而QRS综合波增宽。心 室复极化加速,显示T波高耸狭窄,为高血钾心电图的主要特 点。心室肌APD缩短,则Q-T间期缩短,但也可因QRS综合波增 宽而延长。房室传导减慢,故P-R间期延长。
缩短,故ERP(有效不应期)也随之缩短。
2021/7/26
6
(2)传导性的变化
轻度高钾时,由于RP略有减小,与阈电位更为接近,
则局部电位更易达到阈电位而形成兴奋扩布,故传导性略
有增高。但中度以上高钾时,由于RP显著减小,Na+内流的
电位梯度显著降低和钠通道的部分失活,使APA和Vamx显著
降低,传导性也随之降低而形成传导阻滞。因此,当血钾
2021/7/26
4
(一)高钾
[K+]o高于5.5mmol/L称为高钾血症。[K+]o 5.5~7.Ommol/ L 为 轻 度 高钾 , [K+]。 7 ~ 9mmol / L 为 中度 高 钾 , [K+] 超 过 9mmol/L为重度高钾。
1、膜电位的变化
(1)RP:高钾时,膜内、膜外K+浓度梯度减小,RP绝对值减小, 这种现象称为高钾去极化。
2021/7/26
《心肌的生理特性》课件
Part One
单击添加章节标题
Part Two
心肌的结构和功能
心肌细胞的形态和结构
心肌细胞呈梭形, 有横纹
心肌细胞有收缩性 和舒张性
心肌细胞有自律性 ,可以自动节律性 收缩
心肌细胞有传导性 ,可以传递兴奋
心肌的功能概述
心肌是心脏的主要组成部分,负责心脏的收缩和舒张 心肌具有自动节律性,能够自主地、有规律地收缩和舒张 心肌具有兴奋性,能够对刺激产生反应,并传导兴奋 心肌具有收缩性,能够产生力量,推动血液流动
心脏起搏点的作用
控制心脏跳动的频率和节奏 产生心脏跳动的电信号 维持心脏的正常功能 调节心脏的收缩和舒张
心肌自动节律性的影响因素
离子通道:心肌细胞膜上的离子通道对心肌的自动节律性有重要影响 细胞内钙离子浓度:细胞内钙离子浓度的变化会影响心肌的自动节律性 神经调节:自主神经系统对心肌的自动节律性有调节作用 激素调节:激素水平对心肌的自动节律性有影响 心肌细胞膜电位:心肌细胞膜电位的变化会影响心肌的自动节律性
心肌的电生理特性
心肌细胞:心肌细胞是心肌的主要组成细胞,具有兴奋性和传导性
心肌电生理特性:心肌细胞具有自动节律性、传导性、兴奋性和收缩性
心肌电生理特性的生理意义:心肌电生理特性是心肌正常生理功能的基 础,也是心肌疾病诊断和治疗的重要依据 心肌电生理特性的研究进展:近年来,心肌电生理特性的研究取得了重 要进展,为心肌疾病的诊断和治疗提供了新的思路和方法。
能量供应
心肌细胞具有较高的线粒体 密度,以适应其高代谢率的
需求
心肌的能量来源
心肌细胞通过氧化磷酸化过程产生能量 主要能量来源是葡萄糖和脂肪酸 心肌细胞通过糖酵解和脂肪酸氧化获取能量 心肌细胞在缺氧状态下,主要通过糖酵解获取能量
医学基础知识:心肌的生理特性
在临床医学考试中,循环系统的内容是是考查的重点,更是考查的难点,其中重要的内容就是心肌的生理特性的知识,关于心肌的生理特性的重要考点,中公卫生人才网为大家进行了汇总,希望能帮助大家更好的掌握。
一、心肌的生理特性考点总结
1.绝对不应期加局部反应期等于:有效不应期。
2.绝对不会产生动作电位的时期是:有效不应期。
3.需要阈上刺激的时期是:相对不应期。
4.由于钠通道复活较少,而兴奋性降低的时期是:相对不应期。
5.阈下刺激可兴奋的时期是:超常期。
6.钠通道完全复活的时期是:超常期。
7.传导速度最慢的部位是:房室交界。
8.心脏正常起搏点:窦房结。
9.传导速度最快的是:浦肯野纤维网。
10.自律性最高的是:窦房结。
11.心肌收缩依赖的离子是:钙离子。
12.自律性的基础:4期自动去极化。
13.保证心房心室不同时收缩的是:房-室延搁。
14.保证心室肌几乎完全收缩的是:浦氏纤维。
15.心房收缩完毕后进行的是:心室收缩。
以上就是需要大家掌握的心肌的生理特性内容,在备考的过程中,希望大家勤于总结,高效复习,小编也祝愿大家顺利通过考试。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第三节心肌的生理在循环系统中,心脏起着泵血的功能,推动血液循环。
心脏的这种功能是由于心肌进行节律性的收缩与舒张及瓣膜的活动而实现的。
心肌的收缩活动又决定心肌具有兴奋性,传导性等生理特性。
心肌细胞膜的生物电活动是兴奋性和传导性等生理特性的基础。
故本节先讨论心肌细胞的生物电活动,进而阐明心肌的生理特性。
在此基础上,再进一步讨论心脏的生理功能。
心肌的生理特性心肌组织具有兴奋性、自律性、传导性和收缩性四种生理特性。
兴奋性、自律性和传导性是以肌膜的生物电活动为基础的,故又称为电生理特性。
心肌细胞的生物电现象和神经组织一样,心肌细胞在静息和活动时也伴有生物电变化(又称跨膜电位)。
研究和了解心肌的生物电现象,对进一步理解心肌生理特性具有重大意义。
从组织学,电生理特点和功能可将心肌细胞分为两大类。
一类是普通细胞,含有丰富的肌原纤维,具有收缩功能,称为工作细胞,工作细胞属于非自律性细胞,它不能产生节律性兴奋活动,但它具有兴奋性和传导兴奋的能力。
它们包括心房肌和心室肌。
另一类是一些特殊分化了的心肌细胞,它们含肌原纤维很少或完全缺乏;故已无收缩功能,它们除具有兴奋性、传导性外,还具有自动产生节律性兴奋的能力,故又称自律细胞。
主要包括P细胞和浦肯野细胞。
它们与另一些既不具有收缩功能又无自律性,只保留很低的传导性的细胞组成心脏中的特殊传导系统。
特殊传导系统是心脏中发生兴奋和传导兴奋的组织,起着控制心脏节律性活动的作用。
特殊传导系统包括窦房结、房室交界、房室束和末梢浦肯野纤维。
一、心肌的兴奋性心肌细胞有两类,一类是具有收缩能力的心房肌和心室肌,称工作细胞即非自律细胞;另一类是特殊分化的细胞,自律细胞,构成心脏的特殊传导系统(一)心室肌细胞跨膜电位(非自律细胞)静息电位(Rp)及其形成机制心肌细胞和骨骼肌一样在静息状态下膜内为负,膜外为正,呈极化状态。
这种静息状态下膜内外的电位差称为静息电位。
不同心肌的静息电位的稳定性不同,人和哺乳类动物心脏的非自律细胞的静息电位稳定,膜内电位低于膜外电位/90mV左右(以膜外为零电位,膜内侧为-90mV)。
在自律性细胞如窦房结细胞和浦肯野细胞的静息电位不稳定,称为舒张期电位,不同部位的自律细胞舒张期最大电位不同,浦肯野细胞的最大舒张电位为-90mV,窦房结细胞的最大舒张电位较小,约为-70mV左右。
心肌细胞静息电位产生的原理基本上与神经、骨骼肌相似,主要是由于K+外流所形成。
动作电位(Ap)心肌细胞兴奋过程中产生的並能扩布出去的电位变化称为动作电位。
与骨骼肌相比心肌细胞动作电位升支与降支不对称。
复极过程比较复杂。
不同部分心肌细胞动作电位形态波幅都有所不同。
按照心肌细胞电活动的特点,可以分为快反应细胞和慢反应细胞。
快反应细胞包括:心室肌、心房肌和浦肯野细胞,前二者属非自律细胞,后者属自律细胞。
快反应细胞动作电位的特点是去极化速度快,振幅大,复极过程缓慢並可分几个时相(期)。
由于去极速度快、波幅大,所以兴奋传导快。
慢反应细胞包括窦房结和房室结。
慢反应细胞的主要特点是去极化速度慢,波幅小,复极缓慢且无明显的时相区分,传导速度慢。
1.快反应细胞动作电位及其形成机制快反应细胞的动作电位可分为五个时相(期):0期又称除极或去极过程,心肌细胞受到刺激发生兴奋时出现去极。
膜内电位迅速由静息状态的-80~-90mV上升到+30mV左右,即膜两侧原有的极化状态被消失並呈极化倒转,从去极化到倒极化形成动作电位的升支,其超过0电位的电位称为超射。
0期短暂,仅占1~2ms,而上升幅度大,可达120mV。
其最大除极速度在心房心室肌约为200~300V/S,而浦肯野细胞可达400~800V/S。
1期(快速复极化期),在动作电位去极完毕后,转入复极期在复极初期,膜电位迅速由30mV下降到0mV左右,占时约2ms,1期在不同的快反应细胞明显程度不同,在浦肯野细胞很明显。
2期(缓慢复极化期又称平台期),在2期内,复极速度极为缓慢,几乎停滞在同一膜电位水平,因而形成平台,故又称平台期,平台期是心肌细胞动作电位的主要特征。
不同心肌细胞平台期的电位水平和时程长短不同。
心室肌和房室束近端的浦肯野细胞平台期的电位为零电位附近。
在束支远端或末梢的浦肯野细胞为-40mV。
心室肌细胞平台期时程约占100ms、浦肯野细胞为200~300ms。
平台期的存在是心肌快反应细胞动作时程明显长于神经、骨骼肌的主要原因。
3期(快速复极化末期),2期复极结束后,复极过程又加速,膜内电位下降至静息电位或舒张电位水平,完成复极化过程,占时约为/100~150ms。
4期(动作电位复极完毕后的时期)又称之为电舒张期。
在非自律细胞如心房肌,心室肌细胞4期内膜电位稳定于静息电位,称为静息期。
在自律细胞4期内膜电位不稳定,有自发的缓慢去极倾向称为舒张除极。
当4期除极达到阈电位水平就可产生一次新的动作电位。
形成机制:快反应心肌细胞动作电位形成的原理与骨骼肌基本相似,也是与离子在细胞两侧不均匀分布所形成的浓度梯度和细胞膜上存有特殊离子通道有关。
已知细胞外Na+浓度大于细胞内(约大4倍多)。
而细胞外的K-浓度则比细胞内小30多倍。
相应离子经细胞膜上特殊离子通道的越膜扩散,是形成心肌动作电位的基础。
但心肌跨膜电位形成中涉及的离子远比骨骼肌要复杂得多。
在心肌细胞动作电位的形成除由于离子越膜被动扩散外,由细胞膜上离子泵活动所产生的离子主动转运,在细胞的电活动中也起着重要作用。
以下具体讨论快反应细胞动作电位形成的离子基础。
快反应细胞0期去极与Na+快速内流有关,而Na+的内流除与膜内外Na+浓度梯度有关外,更主要的是决定于Na+通道的状态。
如前所述Na+通道可表现为激活、失活和备用三种状态。
在适当的刺激作用下,首先引起Na+通道的部分开放,少量Na+内流,而引起膜内电位上升。
当膜电位由-90mV升至-70mV时,则Na+通道被激活而开放,通透性增高。
此电位水平即称为阈电位。
由于膜外Na+浓度大于膜内和膜内外电位梯度的影响,大量Na+快速进入膜内,膜内电位急剧上升,由负变为正(-90mV→+30mV)。
而形成动作电位的上升支。
当膜电位负值减少至-55mV以上时,则/Na+通道失活关闭,Na+内流迅速终止。
Na+通道的激活与失活十分迅速故称为快通道。
由快通道开放而出现的电位变化称为快反应电位。
故具有这种特性的心肌细胞称为快反细胞。
关于心肌动作电位1期的形成原理,过去认为是C1-内流所引起,近年研究表明,1期电位可被K+通道阻滞剂四乙基胺和4-氨基吡啶所阻断,因之认为K+的跨膜外流是引起1期的主要离子。
2期(平台期)形成的原因主要是Ca2+的缓慢内流和少量K+外流所形成。
已经证明,心肌膜上存在一种慢Ca2+通道。
慢Ca2+通道的激活,以及再复活所需时间均比Na+通道要长,故称慢通道。
慢通道也是电压依从性,激活慢通道的阈电位水平是-50~-35mV。
由于慢钙通道的选择性不如快钠通道那样专一,它虽然对Ca2+的通透性较高,但也有一定的Na+通透性,约为Ca2+内流的1/70~100。
故在平台期也有一定量的Na+内流。
在平台期早期,Ca2+的内流与K+外流所负载的跨膜正电荷量相等,故膜电位稳定于1期复极的电位水平,随着时间推移,慢Ca2+通道逐渐失活,K+外流逐渐增多,膜内电位缓慢下降,而形成平台期晚期。
3期的形成主要是由于Ca2+通道完全失活,而膜对K+通透性增高,K+外流随时间而递增导至膜的复极愈来愈快,直至复极完成。
在4期内,工作细胞膜电位基本上稳定于静息电位水平。
但膜内外离子分布都与静息电位时不同,即由于前一阶段的变化,膜内Na+,Ca2+有所增加,而K+有所减少。
因此只有把动作电位期间进入细胞内的Na+、Ca2+排出去,把外流出去的K+摄取回来,才能恢复细胞内外正常的离子浓度梯度,保持心肌的正常兴奋能力。
这些离子的转运都是逆浓度梯度进行的主动转运过程。
这种主动转运过程主要也是通过Na+-K+泵的作用,形成Na+-K+交换而实现的。
关于进入膜内Ca2+的转运一般认为与Na+顺浓度梯度的内流相耦合而进行的。
即Na+的内流促使Ca2+外流形成Na+Ca2+交换。
由于Na+的内向性浓度梯度的维持是依靠Na+-K+泵而实现的,故Ca2+的主动转运的能量也是由Na+-K+泵提供的,Ca2+的转运决定于膜两侧Na+的浓度梯度。
故当细胞内Na+的浓度增加时(导致Na+内向性浓度梯度减小),Ca2+的外运也相应减少,细胞内Ca2+将因此而增加。
快反应自律细胞(浦肯野细胞),在4期内膜电位不稳定,研究资料表明,在浦肯野细胞4期出现主要是Na+随时间推移而渐增的内向流动所引起,这种Na+内流的膜通道在3期复极电位达-60mV左右,开始激活开放,其激活程度随膜电位复极化,膜内负电位的增加而增加,至-100mV就充分激活。
因此,Na+内流逐步增大,膜的除极程度逐渐增加,一旦达阈电位水平即能产生另一次动作电位,虽然这种通道允许Na+通过,但与快钠通道不同,因为二者激活的电位水平不同,此外具有阻断快钠通道的河琢毒素(TTX)也不能阻断此通道。
(二)影响心肌兴奋性的因素1.Rp与阈电位差值:与兴奋性成反比2.Na通道状态1)Na通道功能状态:激活、失活、备用2)兴奋性→取决于Na通道的备用状态→取决于膜Rp 是否正常(三)一次兴奋过程中兴奋性的周期性变化(四)兴奋性周期性变化与心肌收缩活动关系1、ERP特别长→保证心肌收 / 舒交替进行不发生强直收缩2、期前收缩(早搏)和代偿间歇特殊情况下,心肌可以接受在窦性节律之外又效不应期之后的刺激,将此额为刺激的收缩称为期前收缩(早搏),在下一个窦性节律未到之前,常常出现一段较长的心室舒张期,成为代偿间歇二. 心肌的自动节律性(一)心肌细胞的分类心肌细胞按有无自律性分:1. 工作细胞(非自律细胞)心房、心室肌→兴、传、收(无自)2. 自律细胞P 细胞、浦肯野细胞等→兴、传、自(无收缩性)(二)自律细胞与自律细胞的跨膜电位1. 自律细胞分类1)快反应自律细胞:去极化因快Na通道开放引起2)慢反应自律细胞:去极化因慢Ca通道开放引起2. 自律性产生基础:自律细胞有4期缓慢自动去极化1)自律细胞没有稳定静息电位(4期),表现为自动发生缓慢去极化无稳定的静息状态,其静息电位在前一个动作电位复极化完毕时为最大(绝对值)称最大舒张电位2)4期缓慢自动去极化产生原因:复极化末期外向电流逐渐减弱(K离子),内向电流逐渐增强 (多为Na离子)3. 自律细胞跨膜电位根据自律细胞Ap 0期去极化速度和产生机制不同,心脏自律细胞可分为窦房结,房室交界的自律细胞属慢反应细胞。
与快反应细胞跨膜电位相比,慢反应细胞电位具有以下特点:(1)慢反应细胞的静息电位和阈电位比快反应电位低。