第四节 单链丝状噬菌体载体
第四节-单链丝状噬菌体载体

第四节单链丝状噬菌体载体一、M13 噬菌体的生物学1.M13 噬菌体的组成和结构M13 噬菌体颗粒是丝状的,只感染雄性大肠杆菌。
感染宿主后不裂解宿主细胞,而是从感染的细胞中分泌出噬菌体颗粒,宿主细胞仍能继续生长和分裂。
M13 噬菌体的基因组为单链 DNA ,由 6407 的碱基组成(GenBank 注册号为V00604)。
基因组 90% 以上的序列可编码蛋白质,共有 11 个编码基因,基因之间的间隔区多为几个碱基。
较大的间隔位于基因Ⅷ 和基因Ⅲ 以及基因Ⅱ 和基因Ⅳ 之间,其间有调节基因表达和 DNA 合成的元件。
M13 噬菌体基因组可编码 3 类蛋白质,包括复制蛋白(基因Ⅱ,Ⅴ 和Ⅹ),形态发生蛋白(基因Ⅰ,Ⅳ 和Ⅺ),结构蛋白(基因Ⅲ 、Ⅵ、Ⅶ、Ⅷ 和Ⅸ)。
所有结构蛋白在形态发生之前都插入在宿主细胞的质膜中。
基因组 DNA 为正链,按基因Ⅱ 至基因Ⅳ 方向合成,与噬菌体的 mRNA 序列同义。
图 3-20 是 M13 噬菌体的遗传图谱。
M13 噬菌体颗粒为丝状长管状结构,长 880nm ,直径 6-7nm 。
噬菌体颗粒的核心由 2700 个基因Ⅷ 编码的结构蛋白呈管状排列而成(图 5-32),成熟的基因Ⅷ 的产物为由 50 个氨基酸残基组成的α 螺旋蛋白。
顶端由 5 个基因Ⅶ 和 5 个基因Ⅸ 产物组成,作用于间隔区中的包装信号。
5 个基因Ⅲ 蛋白和5 个基因Ⅵ 蛋白位于丝杆的末端,参与对性纤毛的吸咐。
图 3-21 是 M13 噬菌体结构模型。
2.M13 噬菌体的增殖吸附是噬菌体感染的第一步, M13 噬菌体只感染具有性纤毛的菌株,携带 F 质粒的菌株可产生性纤毛。
在吸附过程中,噬菌体的基因Ⅲ 蛋白与性纤毛发生作用。
随后丝状噬菌体穿入到性纤毛,基因Ⅲ 蛋白与宿主的 TolQ 、TolR 和 TolA 蛋白发生作用,去除外壳蛋白,致使病毒 DNA 及附着于其上的基因Ⅲ 蛋白进入宿主菌体内。
在宿主细胞内,感染性的单链噬菌体 DNA (正链)在宿主酶的作用下转变成环状双链 DNA ,用于 DNA 的复制,因此这种双链 DNA 称为复制型 DNA (replicative form DNA),即 RF DNA 。
基因工程第三章分子克隆载体

2、可扩增性 质粒就其复制方式而言分为两类:松弛型复制及严谨型复
制。严谨型质粒每个细胞中拷贝数有限,大约 1 ~几个;松驰型
质粒拷贝数较多,可达几百。
严谨控制型 拷贝数少,一般<10个,分子量大;
松弛控制型 拷贝数多,10-200个,分子量小;
复制受限,受细菌宿主DNA复制系 统的控制;
复制不受细菌DNA复制系统限制, 当宿主蛋白质合成受抑制时(如培 养中加入抗生素时),其拷贝数可 猛增至1000-3000之多,该性质对 基因工程技术十分有利。
7、载体DNA的分子量适当,可容纳较大的外源DNA片段。
第一节 质粒载体
一、质粒的基本特性
质粒(plasmid):是 独立于许多细菌及某些 真核细胞染色体外共价 闭合环状的DNA分子, 能独立复制的最小遗传 单位,大小在1—200kb 之间。和病毒不同,它们 没有衣壳蛋白(裸DNA)。
绝大多数质粒DNA是双链环形,它具有3种不同的构型:
来自pSCl01的 四环素抗性基因 Tetr
来自RSF2124的 氨苄青霉素抗性基 因Ampr。
特点
1. 具有多个单一的限制性内切酶位点。
2. Tetr中有BamH1切点(G↓GATCC)和SalⅠ切点 (G↓AATTC),Ampr中有PstⅠ切点(CTGCA↓G)。
3. 利用ColEl的复制子,所以在细胞中是多拷贝的,可通过氯 霉素使质粒拷贝数进一步扩增。
基因工程
第三章 分子克隆载体
商丘师范学院 马原松
பைடு நூலகம்
第三章 分子克隆载体
教学目的和要求:本章主要阐明用于核酸操作的各种载体: 质粒载体、噬菌体载体和单链丝状噬菌体载体等。通过 本章的学习要求学生了解质粒的基本特性,λ噬菌体的 分子生物学,穿梭载体、整合载体、解离载体等其它载 体;理解大肠杆菌表达载体;掌握质粒载体的工作原理, 噬菌体载体种类及工作原理,粘粒载体、噬菌粒载体的 工作原理。 重点、难点:质粒载体、噬菌体载体的种类和工作原理。 教学方法:讲授、讨论、计算机辅助教学等
12第五章 噬菌体载体

234页
噬菌体:细菌病毒。 烈性噬菌体:感染寄主细胞后能进行复制、增殖,最后 引起细胞裂解的噬菌体。只有溶菌生长周期的噬菌体。 温和性噬菌体(溶原性噬菌体):感染细菌后能使细菌 产生溶原性的噬菌体。既能进入溶菌生命周期又能进入 溶原生命周期的噬菌体。 溶原性:温和噬菌体感染细胞形成原噬菌体的过程。 原噬菌体:在溶原性细菌内存在的整合的或非整合的噬 菌体DNA。 溶原性细菌:细胞中含有以原噬菌体状态存在着的温和 噬菌体的细菌。 噬菌斑:在含细菌的固体培养基上,噬菌体使细菌细胞 裂解而形成的空斑。
(243页)
4. 噬菌体的溶源生命周期
Ⅱ、溶源周期的主要特征(以λ噬菌体为原型): (1)噬菌体的DNA分子注入细菌细胞。 (2)经过短暂的转录。 (3)噬菌体的DNA分子插入到细菌染色体基因组 DNA上,变成原噬菌体。 (4)细菌继续生长、增殖,噬菌体的基因作为细 菌染色体的一部分进行复制。
(244页)
基因工程又称重组DNA技术 其实施至少要有四个必要的条件 1. 工具酶 2. 基因 3. 载体 4. 受体细胞
(《基因工程..》静国忠,85页)
(240页)
噬菌体定义: 是一类细菌病毒的总称。 噬菌体与质粒的区别 * 质粒:仅是一种含有复制起点的裸露的DNA分 子。 * 噬菌体:DNA分子除了复制起点之外,还有编 码外壳蛋白质的基因。
Ⅰ、若干基本概念
234页
噬菌体:细菌病毒。 烈性噬菌体:感染寄主细胞后能进行复制、增殖,最后 引起细胞裂解的噬菌体。只有溶菌生长周期的噬菌体。 温和性噬菌体(溶原性噬菌体):感染细菌后能使细菌 产生溶原性的噬菌体。既能进入溶菌生命周期又能进入 溶原生命周期的噬菌体。 溶原性:温和噬菌体感染细胞形成原噬菌体的过程。 原噬菌体:在溶原性细菌内存在的整合的或非整合的噬 菌体DNA。 溶原性细菌:细胞中含有以原噬菌体状态存在着的温和 噬菌体的细菌。 噬菌斑:在含细菌的固体培养基上,噬菌体使细菌细胞 裂解而形成的空斑。
第八讲单链噬菌体载体及噬菌粒载体

第八讲单链噬菌体载体及噬菌粒载体吴乃虎中国科学院遗传与发育生物学研究所第八讲单链噬菌体载体及噬菌粒载体一、单链噬菌体的一般生物学1.单链噬菌体的优越性2.M13噬菌体的生物学特性二、M13克隆体系1.M13克隆体系2.M13克隆体系-半乳糖苷酶的显色反应原理3.M13载体系列的发展4.M13载体系列的优点三、噬菌体展示载体1.噬菌体展示载体的构建原理2.噬菌体展示载体3.噬菌体表面展示文库4.应用噬菌体展示载体分离有关蛋白质的实例四、噬菌粒载体1.M13噬菌体载体克隆的若干难点2.噬菌粒3.若干常用的噬菌粒载体4.pBluescript噬菌粒载体5.pUC118和pUC119噬菌粒载体第八讲单链噬菌体载体一、单链噬菌体一般生物学大肠杆菌丝状单链DNA噬菌体有M13噬菌体、f1噬菌体及fd 噬菌体,它们均含有分子量约为6400个核苷酸的单链闭环DNA分子。
1.单链DNA phage的优越性A.具有双链的复制型DNA(RF DNA),可如质粒质粒一样进行遗传操作;RF DNA:Replication Form DNA。
B.RF DNA和ssDNA均可感染感受态的寄主细胞——形成phaque或colony。
C.不受包装的限制。
因为单链DNA phage的大小是受其DNA 多寡制约的。
D.可容易地测出外源DNA的插入取向。
E.可产生大量的含有外源DNA的单链DNA分子,这种单链DNA分子有如下用途(作为模板):*1用作双脱氧链终止法进行DNA测序*2制备单链的放射性标记的杂交用DNA探针*3利用寡核苷酸进行定点突变2.M13 phage的生物学特性A.M13 phage同f1 phage亲缘关系十分密切,例如:①基因组组织形式相同;②病毒颗粒大小、形状相近;③DNA同源性高达98%以上。
B.在M13 phage颗粒中只有(+)链DNA,感染具F性须的大肠杆菌菌株,因此M13噬菌体是雄性E.coli特有的;M13噬菌体的(+)链DNA,又称为感染性单链DNA。
单链丝状噬菌体展示系统、λ噬茵体展示系统和T4噬茵体展示系统

单链丝状噬菌体展示系统、λ噬茵体展示系统和T4噬茵体展示系统一、单链丝状噬茁体展示系统1、pIII展示系统及噬苗体抗体丝状噬茵体是单链DNA病毒,pIII是病毒的次要外完蛋白(minor coat protein)、位于病毒颗粒的一端,每个病毒颗粒都有3—5个拷贝pIII蛋白,pIII有两个位点可供外源序列插入,即N端和近N端可伸屈胃内。
当抗体片段或蛋白质融合到PIII的N端时,噬菌体仍有感染性,但若融合到后一位点则会切去N端而丧失感染性,这时就需有辅助噬菌体提供野生型 pIII蛋白。
PIII很容易为蛋白水解酶水解。
所以有辅助噬菌体超感染时.可以使每个噬菌体平均显示不到一个融合蛋白,即所谓“单价“噬茵体,从而使抗体部分最大限度地保持原构型而功能完好。
PIII展示系统的主要用途是制备噬茵体抗体,它的突出优点是模拟了自然免疫选择系统。
自然免疫系统中.抗原结合于B细胞表面受体而使其活化并分裂增殖、分化成有抗体分泌功能的浆细胞。
这个过程可以从约5×109个鼠细胞和约1012人细胞中选出一个至几个特异B细胞,并有选择性地富集特异性B细胞,通过多轮突变和选择使抗体亲和力成熟。
pIII展示系统完全模拟了自然选择系统;噬菌体展示的抗体片段可以由抗原包被的板、柱等选择,或者用生物素标记的抗原从液相中捕获。
结合在固相抗原的噬茵体抗体经洗涤后可用可溶性半抗原、酸、碱等洗脱,然后感染大肠杆菌培养扩增,再经下一轮的“吸附—洗脱—扩增”筛选。
首轮筛选可使特异性噬菌体富集20一1000倍,一般经4轮筛选,可富集107倍。
对初步筛选的抗体,可以用错构酶及PCR锗配技术等实行多轮突变或采用链置换法使其亲和力成熟。
用PIII系统制备抗体的基本程序是:从经免疫或未免疫者获取淋巴细胞(外周血淋巴细胞,或脾、淋巴结、骨髓等的淋巴细肥),提取细胞mRNA(或细胞基因组DNA),逆转录成cDNA,用PCR方法扩增抗体重链和轻链基因,若制备ScFv抗体片段,还需设计接头Linker,如(Gly 4— Ser),做成VH—Linker—VL连接)。
第四章 基因工程的质粒载体

SC
2 质粒DNA的转移
(1)质粒的类型:在大肠杆菌中的质粒,可 以分为:
接合型质粒:能自我转移
具有自主复制的基因,控制细菌配对和质粒接合转 移的基因。
非接合型质粒 不能自我转移
按接合转移功 能分类
非接合型质粒
主要基因
自主复制基因,产生大肠杆菌素基因
按抗性记号 分类
Col质粒
接合型质粒
自主复制基因,抗菌素抗性基 因
第二代 酵母表达 穿梭质粒 体系
第三代 哺乳类细 病毒、脂质体 胞表达体系
第四代 基因直接 DNA本身 导入
细菌 酵母 培养动物细胞 生殖细胞、 体细胞、个体
(三)基因工程载体必须具备的条件:
※(1)有复制起点 ※(2)具有若干个限制性内切酶的单一识别位点 ※(3)具备合适的筛选标记 ※(4)具备合适的拷贝数目
(c)所示,F质粒无力帮助mob-突变体进行转移,其中F性须和转移装置虽已 形成,但ColE1 DNA并没有发生缺口。
(d)表示另一种具mob+表型并带有一个顺式显性突变的ColE1突变体,它缺 失了bom位点。在这样的寄主细胞中,虽然能够合成mob蛋白质,但由于不 能发生缺口,因此仍然不能够转移。
3.若质粒DNA经过适当的核酸内切 限制酶切割之后,发生双链断裂形成 线性分子(IDNA),通称L构型
3-基因工程载体的构建
三、单链噬菌体载体
单链环状DNA的丝状大肠杆菌噬菌体: M13、f1、fd 噬菌体
M13 噬菌体
1. 单链DNA噬菌体的特点 (1)+DNA。(ssDNA) (2)复制型(RF)是双链环状DNA。 (3)RF DNA和ssDNA都能转染感受态 大肠杆菌。并产生噬菌斑。
(4)不存在包装限制。
(5)可产生大量的含有外源DNA插入片 段的单链分子,便于作探针或测序。
pUC118/p pUC18/ UC119 pUC19 pBS pUC
M13
M13 f1
M13变异 XS127, XS101 株 M13K07 MV1184
M13K07 XL1-Blue
辅助噬菌体用来复制和包装噬菌粒载体。
(3)pUC118/pUC119
① 构成
1)M13的基因间隔区(IG) 带有M13复制起点。 2)pUC18/pUC19质粒载体: 质粒复制起点 Ampr lacZ’ MCS
包装
辅助M13
外壳蛋白
③ 噬菌粒载体的包装
1)辅助噬菌体:
如M13K07等。
自身的复制起点发生了突变,复制效率 低下,但感染细菌后能表达出外壳蛋白 和复制蛋白,帮助噬菌粒载体复制。 2)包装: 辅助噬菌体 外壳蛋白和 复制蛋白 噬菌体
噬菌粒载体
ssDNA
(4)pBluescript 噬菌粒载体(pBS) Stratagene公司发展的一类噬菌粒载体。 ① 组成 由pUC质粒载体、f1噬菌体的复制起点和 T3、T7噬菌体的启动子组成。 ② 特点 1)定向体外转录
M13
(2)噬菌体展示载体的构建
把外源DNA片段插入基因Ⅲ的序列前端, 并保持两者的阅读框正确,表达出融合 蛋白。 启动子 外源DNA M13基因Ⅲ
DNA重组(DNA recombination)技术:DNA重组的载体-2

DNA重组(DNA recombination)技术:DNA重组的载体-2二、噬菌体载体作为细菌寄生物的噬菌体,大多数具有编码多种蛋白质的基因,能利用宿主细胞的蛋白质合成体系,进行生长和增殖。
构建的噬菌体载体,以λ噬菌体、M13和粘粒最为常用。
㈠λ噬菌体载体野生型λDNA是一种基因组为4.8 kb的线性双链DNA,全部序列已知,共编码50多个基因。
其中约一半基因参与了噬菌体的生命周期活动,称为必需基因;另一部分基因,当被外源基因取代后,并不影响噬菌体的生命功能。
另外,在分子两端各有12个碱基的互补单链,是天然的粘性末端,称cos位点。
野生型的λ噬菌体是一种中等大小的温和噬菌体,即既能进入溶菌性生命周期又能进入溶源性生命周期。
λ噬菌体感染细菌后,λDNA通过粘性末端而环化。
既可溶菌性生长(裂解),即λDNA在宿主中多次复制并合成大量基因产物,装配成噬菌体颗粒,最后裂解宿主菌;又可以溶源性生长,即λDNA整合到宿主染色体基因组D NA中,与之一起复制并遗传给子代,但宿主细胞不被裂解。
裂解途径和溶源途径的选择取决于宿主与噬菌体之间的许多因素的相互作用,以及噬菌体基因组的精确调控。
λ噬菌体具有二个重要特征,使之成为一类重要的、很有价值的基因克隆载体。
①除必需基因外的功能相关(溶源生长和重组)基因成簇排列在一起位于中间部分,约占全基因组的30%,不是溶菌生长所必需的。
因此,该区可缺失或作外源DNA的插入区。
②噬菌体颗粒成熟的包装过程(外壳蛋白包装DNA),只要重组DNA不小于野生型λDNA全长75%和大于105%,均可被包装成具有噬菌体活性的成熟颗粒。
因此大约可插入长达15kb的外源DNA。
当外源DNA片段克隆到插入型载体上,噬菌体便丧失某些生物学功能,即插入失活效应。
又可分为免疫功能失活的和β-半乳糖苷酶失活的两种亚型。
①免疫功能失活的插入型载体。
插入位点位于基因组中的免疫编码区,外源DNA片段插入后,就会使载体合成阻遏蛋白的能力被破坏,导致不能进入溶源生长周期。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
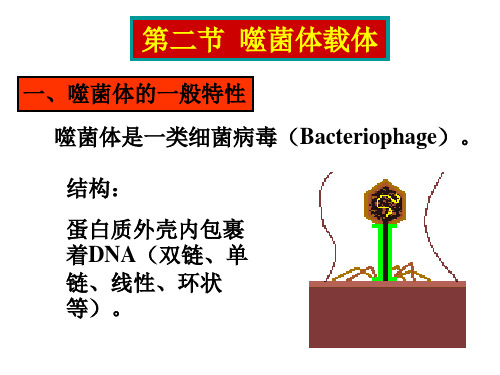
第四节单链丝状噬菌体载体一、M13 噬菌体的生物学1.M13 噬菌体的组成和结构M13 噬菌体颗粒是丝状的,只感染雄性大肠杆菌。
感染宿主后不裂解宿主细胞,而是从感染的细胞中分泌出噬菌体颗粒,宿主细胞仍能继续生长和分裂。
M13 噬菌体的基因组为单链 DNA ,由 6407 的碱基组成(GenBank 注册号为V00604)。
基因组 90% 以上的序列可编码蛋白质,共有 11 个编码基因,基因之间的间隔区多为几个碱基。
较大的间隔位于基因Ⅷ 和基因Ⅲ 以及基因Ⅱ 和基因Ⅳ 之间,其间有调节基因表达和 DNA 合成的元件。
M13 噬菌体基因组可编码 3 类蛋白质,包括复制蛋白(基因Ⅱ,Ⅴ 和Ⅹ),形态发生蛋白(基因Ⅰ,Ⅳ 和Ⅺ),结构蛋白(基因Ⅲ 、Ⅵ、Ⅶ、Ⅷ 和Ⅸ)。
所有结构蛋白在形态发生之前都插入在宿主细胞的质膜中。
基因组 DNA 为正链,按基因Ⅱ 至基因Ⅳ 方向合成,与噬菌体的 mRNA 序列同义。
图 3-20 是 M13 噬菌体的遗传图谱。
M13 噬菌体颗粒为丝状长管状结构,长 880nm ,直径 6-7nm 。
噬菌体颗粒的核心由 2700 个基因Ⅷ 编码的结构蛋白呈管状排列而成(图 5-32),成熟的基因Ⅷ 的产物为由 50 个氨基酸残基组成的α 螺旋蛋白。
顶端由 5 个基因Ⅶ 和 5 个基因Ⅸ 产物组成,作用于间隔区中的包装信号。
5 个基因Ⅲ 蛋白和5 个基因Ⅵ 蛋白位于丝杆的末端,参与对性纤毛的吸咐。
图 3-21 是 M13 噬菌体结构模型。
2.M13 噬菌体的增殖吸附是噬菌体感染的第一步, M13 噬菌体只感染具有性纤毛的菌株,携带 F 质粒的菌株可产生性纤毛。
在吸附过程中,噬菌体的基因Ⅲ 蛋白与性纤毛发生作用。
随后丝状噬菌体穿入到性纤毛,基因Ⅲ 蛋白与宿主的 TolQ 、TolR 和 TolA 蛋白发生作用,去除外壳蛋白,致使病毒 DNA 及附着于其上的基因Ⅲ 蛋白进入宿主菌体内。
在宿主细胞内,感染性的单链噬菌体 DNA (正链)在宿主酶的作用下转变成环状双链 DNA ,用于 DNA 的复制,因此这种双链 DNA 称为复制型 DNA (replicative form DNA),即 RF DNA 。
通过θ 复制方式, RF DNA 进行扩增,基因的转录也随即开始。
基因组中的任意一个启动子都可以启动基因的转录,单方向地终止于下游的终止子。
启动子和终止子的位置关系使得靠近终止子的基因转录更频繁。
M13 噬菌体在感染细胞中的复制见图 3-22 。
当基因Ⅱ 蛋白在亲代 RF DNA 的正链特定位点上产生一个切口时,便启动噬菌体基因组进行滚环复制。
此时,在大肠杆菌的 DNA 聚合酶Ⅰ 的作用下,以负链为模板在切口的 3' 末端加入核苷酸,并持续 DNA 的合成,用新合成的 DNA 替换原有的正链。
当复制叉环绕模板整整一周时,被取代的正链由基因Ⅱ 产物切去,经环化后形成单位长度的噬菌体基因组 DNA 。
在感染开始的 15~20 分钟内,这些子代正链在宿主细胞酶的作用下,又转变成 RF DNA ,然后以之为模板继续转录并继续合成子代正链 DNA 。
当感染细胞内累计有 100-200 个 RF DNA 时,细胞内也产生了足够的单链 DNA 结合蛋白,即基因Ⅴ 蛋白。
该蛋白可以抑制翻译活性,特别是抑制基因Ⅱ mRNA 的翻译,并且强烈地结合在新合成的正链 DNA 上,阻止其转化成 RF DNA 。
此时, DNA 的合成几乎只产生子代正链 DNA 。
另外,基因Ⅹ 蛋白和基因Ⅴ 蛋白也是噬菌体特异 DNA 合成的强力抑制子,从而限制感染细胞内 RF DNA 的数量。
结果,感染细胞内 RF DNA 的数目和子代正链 DNA 的产生速率都能保持适度。
成熟噬菌体颗粒由 11 个病毒蛋白中的 5 个组成,至少 4 个其他蛋白(如基因Ⅰ 、Ⅳ 和Ⅺ 蛋白,以及宿主的硫氧还蛋白(thioredoxin)对噬菌体颗粒的组装和分泌是必须的。
二、M13 噬菌体载体单链 DNA 的酶切和连接是比较困难的,因此 M13 噬菌体在用作载体时是利用其双链状态的 RF DNA。
RF DNA 很容易从感染细胞中纯化出来,可以象质粒一样进行操作,并可通过转化方法再次导入细胞。
1.载体的插入位点在 M13 噬菌体基因组中绝大多数为必需基因,只有两个间隔区可用来插入外源DNA(基因Ⅱ/Ⅳ 和基因Ⅷ/Ⅲ 之间)。
基因Ⅱ 和基因Ⅳ 之间的 508bp 间隔区是主要的外源片段插入位点。
在基因Ⅷ 和基因Ⅲ 之间的小间隔区也可用来插入外源片段,但是在操作过程中,需要获得感染细胞的菌落进行影印筛选,比利用可见的噬菌斑方法更慢更麻烦。
基因Ⅹ 也可用来克隆外源片段。
2.M13 噬菌体载体组成现在所使用的 M13 噬菌体载体是 Messing 及其同事建立的 mp 系列载体,以基因Ⅱ 和基因Ⅳ 之间的区域作为外源 DNA 插入区。
mp 载体系列都是从同一个重组 M13 噬菌体(M13mpl)改造而来的。
在 M13mpl 载体中,间隔区内的HaeⅢ 位点插入了一小段大肠杆菌 DNA ,引入α 互补筛选。
3.M13mpl8 和 M13mpl9M13mpl8 和 M13mpl9 这两个载体含有 13 个不同的酶切位点,可供插入由多种各不相同的限制酶切割而成的 DNA 片段(图 3-23)。
M13mpl8 和 M13mpl9 DNA 的全序列已经测定完成(GenBank 注册号为 M77815 和 L08821),这两种载体只是在lacZ 区内不对称的多克隆区的方向上有所不同。
当 RF DNA 被两种不同的限制酶切割以后, M13mpl8 和 M13mpl9 轻易不能重新环化。
仅当连接混合液中含有带匹配末端的外源双链 DNA片段时,方可闭合成环。
这一片段在 M13mpl8 和 M13mpl9 中将以两个互为相反的方向插入。
这样一来,在 M13mpl8 的正链中含有外源 DNA 双链的其中一条链,而在 M13mpl9 正链中则含有外源 DNA 的另一条链,即 M13mpl8 重组体的子代噬菌体内含有外源DNA 的一条链, M13mpl9 重组体的子代噬菌体内则含有它的互补链。
故用M13mpl8 和 M13mpl9 作为一对载体,就可能用一个引物(通用引物),从所插入 DNA 片段的任一端开始,测定互为相反的两条链的 DNA 序列,并可制备只与外源 DNA 的任意一条链互补的 DNA 探针。
三、 M13 噬菌体载体的宿主菌由于 M13 噬菌体通过 F 质粒编码的性纤毛进入宿主细胞内,故只能用雄性细菌来增殖病毒。
Messing 及其同事已经构建了许多携带 F' 质粒并便于 M13 载体进行基因操作的多种大肠杆菌菌株,其中最重要的遗传标志有:(1)lacZ D Ml5 lacZ 基因缺失突变体。
(2) D(lac-proAB)lac 基因缺失突变体。
(3)lacI q lacI 基因的突变体。
(4)proAB 细菌染色体上脯氨酸生物合成酶类的编码区域。
(5)traD36 抑制 F' 因子接合转移的突变。
(6)hsdR l7 与hsdR 4 对大肠杆菌 K 株Ⅰ 类限制 - 修饰系统失去限制活性但仍保留修饰功能的突变体。
(7)recA1大肠杆菌重组酶基因。
(8)supE 琥珀抑制基因。
在 M13 噬菌体载体中进行克隆时常用的宿主菌株如下。
JM101 supE D (lac-proAB)[F'traD 36 proAB + lac I q lacZ D M15]JM105 JM101/ hsdR 4JM107 JM101/ hsdR 17JM109 JM101/ hsdR 17 recA1TG1 JM101/ hsd D5(不修饰不限制)XL1-Blue supE lac- hsdR 17 recA1 [F' proAB + lac I q lacZ D M15]Tn 10 (Tet r)四、丝状噬菌体载体克隆中经常遇到的问题在丝状噬菌体载体的克隆中经常遇到两类问题。
(1)外源 DNA 区段的部分缺失(2)外源 DNA 总是以单方向插入五、噬菌粒噬菌粒(phagemid)实际上是带有丝状噬菌体大间隔区的质粒载体,是集质粒和丝状噬菌体的有利特征于一身的载体,具有 ColE1 复制起点及抗生素抗性选择标记的质粒,以及丝状体噬菌体的间隔区。
此间隔区含噬菌体 DNA 合成的起始与终止及噬菌体颗粒形态发生所必需的全部顺式作用序列。
含噬菌粒的细菌被噬菌体感染后,基因Ⅱ 蛋白可作用于噬菌粒的间隔区,启动滚环复制产生 ssDNA 并进行包装。
克隆于这些载体内的外源 DNA 区段可以象质粒一样用常规方法进行增殖。
而带有这种质粒的细菌被 M13(或 f1)丝状噬菌体感染后,在病毒的基因Ⅱ 蛋白影响下,质粒的复制方式发生改变。
基因Ⅱ 产物与质粒所携带的基因间隔区相互作用,启动滚环复制,产生质粒 DNA 一条链的拷贝,最终包装在子代噬菌体颗粒中。
pUC118 和 pUC119 是功能比较完善的噬菌粒载体,对外源 DNA 片段的大小不那么敏感,并且保留了 pUC 质粒在克隆操作方面的优点(图 3-24)。
在 pUC18/19 中增加了带有 M13 噬菌体 DNA 合成的起始与终止以及包装进入噬菌体颗粒所必需的顺式序列(IG),当含这些质粒的细胞被适当的 M13 丝状噬菌体感染时,可合成质粒 DNA 的其中一条链,并包装在子代噬菌体颗粒中。
通过纯化噬菌体颗粒,可制备单链 DNA ,用于 DNA 序列测定、定点诱变或制备探针。
噬菌粒具有以下特征:① 双链 DNA 既稳定,又高产,具有常规质粒的特征;② 免却了将外源 DNA 片段从质粒亚克隆于噬菌体载体这一既繁琐又费时的步骤;③ 由于载体足够小,故可得到长达 10kb 的外源 DNA 区段的单链。
噬菌粒的主要优点在于它可以产生大段外源 DNA 的适量单链拷贝,又不必担心发生缺失突变,而这些外源 DNA 区段常常由于太大而不能克隆于常规 M13 载体。
但许多噬菌粒都有一个主要缺点,即辅助噬菌体感染后,有时候单链 DNA 的产量较低且重复性较差。
六、M13KO7 辅助噬菌体M13KO7 辅助噬菌体是 M13 的衍生株,大小为 8.7kb ,其结构组成见下图。
M13KO7 噬菌体带有来自 p15A 质粒的复制起点,因此可以象质粒一样复制,且可以与带 ColE1 的质粒共存于同一个宿主菌中。
还带有一个来自转座子 Tn903 的卡那霉素抗性基因(kan r),用作选择标记。
基因Ⅱ 带有一个 G 至 T 的突变(第 6125 核苷酸),导致基因Ⅱ 蛋白第 40 位氨基酸由甲硫氨酸变为异亮氨酸。