【医学PPT课件大全】 蛋白激酶、蛋白磷酸酶与信号转导
合集下载
蛋白激酶和蛋白磷酸酶蛋白激酶

1. IP3/Ca2+途径 IP3引起细胞膜和内质网 膜上的Ca2+通道开放,胞浆Ca2+显著增加, 导致细胞活化。
2. DAG/PKC途径 DAG特异性激活丝/苏 氨酸激酶-PKC家族。
G蛋白 PI(4,5)P2 PLCb PLCg
其它磷脂酶 离子通道
PI3-K PI3
DAG
腺苷酸环化酶
Ca2+ PKC
氨酸磷酸化后可结合SHP2,进而抑制TCR相
关的NFAT活化。
(二) 蛋白激酶和蛋白磷酸酶
蛋白激酶:将磷酸供体分子如ATP的磷酸基 团转移到底物蛋白氨基酸受体的酶类。
蛋白磷酸酶:催化磷酸化蛋白分子的磷酸 酯键发生去磷酸化反应的酶类。
蛋白酪氨酸激酶
受体型蛋白酪氨酸激酶:PLC、
abcddd PI3K、GAP、STAT等
(五)转录因子
NF-kappaB:发现于B细胞kappa轻链转录调 控,受多种蛋白的精细调控
Rel蛋白家族成员组成的同源或异源二聚体,分 部最广的是p50/RelA。Rel同源结构域(RHD) 存在与其他Rel蛋白、DNA、IkappaB结合区以 及核定位序列。
IkappaB磷酸化、泛素化、降解导致NFkappaB活化、入核
最早发现于T细胞的核转录因子家族,主要 参与钙离子内流有关的膜受体信号传导。
NFAT同源结构域含有核定位序列、钙调磷 酸酶作用位点。
主要的活化方式:钙调磷酸酶催化去磷酸化、 核转位、结合DNA
四、信号传递途径
(一) IP3/Ca2+途径和DAG/PKC途径:
受体相关的磷脂酰肌醇磷脂酶g2(PI-PLCg2) 被PTK磷酸化后激活,水解细胞膜内侧的磷 脂酰4,5-二磷酸肌醇(PIP2)成为1,4,5-三磷酸 肌醇(IP3)和1,2-二酰基甘油(DAG)。
2. DAG/PKC途径 DAG特异性激活丝/苏 氨酸激酶-PKC家族。
G蛋白 PI(4,5)P2 PLCb PLCg
其它磷脂酶 离子通道
PI3-K PI3
DAG
腺苷酸环化酶
Ca2+ PKC
氨酸磷酸化后可结合SHP2,进而抑制TCR相
关的NFAT活化。
(二) 蛋白激酶和蛋白磷酸酶
蛋白激酶:将磷酸供体分子如ATP的磷酸基 团转移到底物蛋白氨基酸受体的酶类。
蛋白磷酸酶:催化磷酸化蛋白分子的磷酸 酯键发生去磷酸化反应的酶类。
蛋白酪氨酸激酶
受体型蛋白酪氨酸激酶:PLC、
abcddd PI3K、GAP、STAT等
(五)转录因子
NF-kappaB:发现于B细胞kappa轻链转录调 控,受多种蛋白的精细调控
Rel蛋白家族成员组成的同源或异源二聚体,分 部最广的是p50/RelA。Rel同源结构域(RHD) 存在与其他Rel蛋白、DNA、IkappaB结合区以 及核定位序列。
IkappaB磷酸化、泛素化、降解导致NFkappaB活化、入核
最早发现于T细胞的核转录因子家族,主要 参与钙离子内流有关的膜受体信号传导。
NFAT同源结构域含有核定位序列、钙调磷 酸酶作用位点。
主要的活化方式:钙调磷酸酶催化去磷酸化、 核转位、结合DNA
四、信号传递途径
(一) IP3/Ca2+途径和DAG/PKC途径:
受体相关的磷脂酰肌醇磷脂酶g2(PI-PLCg2) 被PTK磷酸化后激活,水解细胞膜内侧的磷 脂酰4,5-二磷酸肌醇(PIP2)成为1,4,5-三磷酸 肌醇(IP3)和1,2-二酰基甘油(DAG)。
蛋白磷酸化与蛋白激酶幻灯片

因此可同时与两种以上的其他信号分子结 合。 • 同一类结构域可存在于多种不同的信号分 子中。 • 本身均为非催化结构域。
64
2)非受体酪氨酸蛋白激酶的种类 分为11个家族,至少有30个成员。它
们介导多种生长因子受体、细胞因子受体、 淋巴细胞抗原受体以及黏附分子整合素的 信号转导。
65
a)Src激酶家族 Src是原癌基因c-Src的产物,参与抗
protein,GAP)以及crk、abl和vav原癌基
因产物等。
50
SH2能特异地识别磷酸化的酪氨酸残 基以及磷酸化残基的羧基端氨基酸序列并 与其相互结合。
SH2的主要功能是介导胞质内多种信 号蛋白的相互连接,形成蛋白异聚体复合 物,从而调节信号传递。
51
Cterminal PLC SH2 domain
1. 丝氨酸/苏氨酸蛋白激酶
丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase,S/T-PK)催化 丝氨酸/苏氨酸的羟基磷酸化。
NH HC CH2 OH OC
NH HC CH2 OC
O OP O
O
13
(1)蛋白激酶A(protein kinase A,PKA) 即cAMP依赖性蛋白激酶。 全酶存在胞浆,被cAMP激活后,催化亚
7
2. 调节结构域/亚基 同源性较低。 作用: 调节酶的活性; 靶向作用,与酶的亚细胞定位有关。
8
9
(二)蛋白激酶的种类
10
11
真核细胞的蛋白激酶可分为五类: ① 丝氨酸/苏氨酸蛋白激酶 ② 酪氨酸蛋白激酶 ③ 组/赖/精氨酸蛋白激酶 ④ 半胱氨酸蛋白激酶 ⑤ 天冬氨酸/谷氨酸蛋白激酶
12
86
2)受体酪氨酸激酶激活信号蛋白的机制 一是信号分子的膜转位; 二是通过结构改变而被激活; 三是通过酪氨酸的磷酸化被激活。
64
2)非受体酪氨酸蛋白激酶的种类 分为11个家族,至少有30个成员。它
们介导多种生长因子受体、细胞因子受体、 淋巴细胞抗原受体以及黏附分子整合素的 信号转导。
65
a)Src激酶家族 Src是原癌基因c-Src的产物,参与抗
protein,GAP)以及crk、abl和vav原癌基
因产物等。
50
SH2能特异地识别磷酸化的酪氨酸残 基以及磷酸化残基的羧基端氨基酸序列并 与其相互结合。
SH2的主要功能是介导胞质内多种信 号蛋白的相互连接,形成蛋白异聚体复合 物,从而调节信号传递。
51
Cterminal PLC SH2 domain
1. 丝氨酸/苏氨酸蛋白激酶
丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase,S/T-PK)催化 丝氨酸/苏氨酸的羟基磷酸化。
NH HC CH2 OH OC
NH HC CH2 OC
O OP O
O
13
(1)蛋白激酶A(protein kinase A,PKA) 即cAMP依赖性蛋白激酶。 全酶存在胞浆,被cAMP激活后,催化亚
7
2. 调节结构域/亚基 同源性较低。 作用: 调节酶的活性; 靶向作用,与酶的亚细胞定位有关。
8
9
(二)蛋白激酶的种类
10
11
真核细胞的蛋白激酶可分为五类: ① 丝氨酸/苏氨酸蛋白激酶 ② 酪氨酸蛋白激酶 ③ 组/赖/精氨酸蛋白激酶 ④ 半胱氨酸蛋白激酶 ⑤ 天冬氨酸/谷氨酸蛋白激酶
12
86
2)受体酪氨酸激酶激活信号蛋白的机制 一是信号分子的膜转位; 二是通过结构改变而被激活; 三是通过酪氨酸的磷酸化被激活。
第五章蛋白激酶

PLC-1的PH domain与IP3结合
血影蛋白的PH domain与膜磷脂结合
e)PTB结构域
PTB结构也可识别一些含磷酸化酪氨 酸的模体。
Shc PTB Domain
非受体酪氨酸蛋白激酶重要结构域的特点:
• 一个信号分子可含有两种以上的结构域, 因此可同时与两种以上的其他信号分子结 合。
(一)蛋白激酶的结构
共同的结构特征:
• 保守的催化结构域/亚基
• 调节结构域/亚基
• 其他功能结构域
PKC
1. 催化结构域/亚基
催化核心含有12个高度保守的亚区。
功能: • 与蛋白质或多肽底物结合; • 与磷酸供体ATP/GTP结合; • 转移磷酸基到底物相应的氨基酸残基上。
2. 调节结构域/亚基
p p
ANK ANK ANK ANK
Kinase domain
Mig-2
RTK
paxillin
Integrins
ECM
Generously provided to the CMC by Cary Wu (Apr-03
(9)DNA依赖性蛋白激酶 DNA依赖性蛋白激酶(DNA-dependent protein kinaes,DNA-PK)可磷酸 化许多核蛋白,包括核受体、转录因子、 DNA拓扑异构酶和RNA聚合酶Ⅱ等。 DNA-PK可发生自主磷酸化,其催化 亚基和Ku蛋白都依赖于DNA和ATP而磷酸 化。
(2)受体酪氨酸蛋白激酶
PDGF一类的跨膜受体,可磷酸化靶蛋 白的酪氨酸,因此称为受体酪氨酸蛋白激 酶(receptor tyrosine kinase,RTK)。
1)受体酪氨酸激酶的自主磷酸化位点
常位于受体的羧基末端胞质域的非催 化区,或质膜与酪氨酸激酶结构域之间。 主要功能是与靶蛋白的SH2结构域结合, 激活靶蛋白。 靶蛋白与磷酸化位点的结合依赖于 pTyr附近的氨基酸组成及顺序。
蛋白磷酸化与蛋白激酶(医学相关)

优质课件
37
(7)核糖体S6激酶(S6K)
包括S6KⅠ和S6KⅡ,能催化核糖体 S6蛋白磷酸化。
优质课件
38
优质课件
39
(8)整合素连接激酶
整合素连接激酶(intergrin-linked kinase,ILK)可直接磷酸化PKB/Akt, 其活性依赖PI3K。
优质课件
40
PINCH, ILK domain structures & interacting partners
优质课件
12
1. 丝氨酸/苏氨酸蛋白激酶
丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase,S/T-PK)催化 丝氨酸/苏氨酸的羟基磷酸化。
NH HC CH2 OH OC
NH HC CH2 OC
O OP O
O
优质课件
13
(1)蛋白激酶A(protein kinase A,PKA)
• 与蛋白质或多肽底物结合; • 与磷酸供体ATP/GTP结合; • 转移磷酸基到底物相应的氨基酸残基上。
优质课件
7
2. 调节结构域/亚基 同源性较低。 作用: 调节酶的活性; 靶向作用,与酶的亚细胞定位有关。
优质课件
8
优质课件
9
(二)蛋白激酶的种类
优质课件
10
优质课件
11
真核细胞的蛋白激酶可分为五类: ① 丝氨酸/苏氨酸蛋白激酶 ② 酪氨酸蛋白激酶 ③ 组/赖/精氨酸蛋白激酶 ④ 半胱氨酸蛋白激酶 ⑤ 天冬氨酸/谷氨酸蛋白激酶
• 广泛分布于各组织的胞质,以Ca2+依赖 的形式从胞质中移位到细胞膜上,此过程 称之为转位。PKC转位是其活化的标志。
【医学PPT课件】蛋白激酶A(PKA)介导的信号通路

• 每个调节亚基都含有2个高亲和性的 cAMP结合部位:A和B,位于肽段的羧 基端. 这2个cAMP结合部位都与蛋白激酶 A全酶的解聚以及激活有关,而且cAMP 的结合与全酶的激活呈强的正协同效应. 首先,cAMP与B部位的结合改变了调节 亚基的构象,从而使A部位更易于结合 cAMP. A部位与cAMP的结合则令调节亚
腺苷酸环化酶的激活和cAMP 的产生
• G蛋白和腺苷酸环化酶(ACase)的活化 • 腺苷酸环化酶的磷酸化抑制与脱敏 • cAMP在胞内水平的调节
• 胞外配基(如激素、核苷酸、炎症因子等)
与腺苷酸环化酶偶联的膜受体相结合, 通过激活性GTP-结合蛋白(Gs)的介导, 诱导了腺苷酸环化酶的活化. ACase通过 水解ATP产生环腺苷酸(cAMP)而提高了 胞内的cAMP水平.
• 在哺乳动物中,依据氨基酸顺序的不同, 催化亚基主要分为两类同工酶:Cα和Cβ, 它们催化的底物具有如下共有序列: XArg-Arg-X-Ser-X (X代表任何1个中性氨 基酸,Arg在激酶的底物识别中有着重要 的作用). PKA的催化亚基含有3个结合部 位:Mg-ATP结合部位,催化部位以及底 物肽结合部位.
• 导致了调节亚基的结构变化而与催化亚 基解离产生1个催化亚基二聚体和2个激 活的催化亚基单体[2]. 其解离反应式 可以表示如下:R2C2+4cAMP R2(cAMP)4+2C*(指活化态,对(ⅰ) 蛋白 激酶A的催化亚基. 蛋白激酶A为一丝
氨酸/苏氨酸蛋白激酶,它的催化亚基 可以使特异底物肽段的Ser或Thr残基磷酸 化.
• 随着克隆技术的快速发展,新发现的蛋 白激酶及其同工酶以几何级数的速度增 长着,对它们的研究也日趋深入. 其中, 蛋白激酶A是最小、最简单,也是研究最 为透彻的蛋白激酶之一.
蛋白激酶

protein,GAP)以及crk、abl和vav原癌基
因产物等。
SH2能特异地识别磷酸化的酪氨酸残 基以及磷酸化残基的羧基端氨基酸序列并 与其相互结合。
SH2的主要功能是介导胞质内多种信 号蛋白的相互连接,形成蛋白异聚体复合 物,从而调节信号传递。
Cterminal PLC SH2 domain
c)SH3结构域
可见于多种胞质信号蛋白及肌动蛋白 结合蛋白中。
SH3识别的部位是一些富含脯氨酸的 区域PXXP。
功能:参与PTK介导的蛋白质间的相 互作用,可能在亚细胞定位和细胞骨架蛋 白相互作用中起作用。
SH3 Domain
d)PH结构域
最初于一种血小板内PKC底物 pleckstrin中发现的结构域,称为 pleckstrin 同源 (pleckstrin homology, PH) 结构域。
PINCH, ILK domain structures & interacting partners
Nck-2
SH3 SH3 SH3
SH2
PINCH
LIM5 LIM4 LIM3 LIM2
LIM1
ILK
pp
ANK ANK ANK ANK PH
RTK
CH-ILKBP/actopaxin parvins/affixin
黏附斑激酶(focal adhesion kinase, FAK)在黏附分子整合素介导的细胞与细 胞外基质的黏附和信号转导中具有起始作 用。
整 合 素 结 构 模 型
Linear structure of FAK
(2)受体酪氨酸蛋白激酶
PDGF一类的跨膜受体,可磷酸化靶蛋 白的酪氨酸,因此称为受体酪氨酸蛋白激 酶(receptor tyrosine kinase,RTK)。
因产物等。
SH2能特异地识别磷酸化的酪氨酸残 基以及磷酸化残基的羧基端氨基酸序列并 与其相互结合。
SH2的主要功能是介导胞质内多种信 号蛋白的相互连接,形成蛋白异聚体复合 物,从而调节信号传递。
Cterminal PLC SH2 domain
c)SH3结构域
可见于多种胞质信号蛋白及肌动蛋白 结合蛋白中。
SH3识别的部位是一些富含脯氨酸的 区域PXXP。
功能:参与PTK介导的蛋白质间的相 互作用,可能在亚细胞定位和细胞骨架蛋 白相互作用中起作用。
SH3 Domain
d)PH结构域
最初于一种血小板内PKC底物 pleckstrin中发现的结构域,称为 pleckstrin 同源 (pleckstrin homology, PH) 结构域。
PINCH, ILK domain structures & interacting partners
Nck-2
SH3 SH3 SH3
SH2
PINCH
LIM5 LIM4 LIM3 LIM2
LIM1
ILK
pp
ANK ANK ANK ANK PH
RTK
CH-ILKBP/actopaxin parvins/affixin
黏附斑激酶(focal adhesion kinase, FAK)在黏附分子整合素介导的细胞与细 胞外基质的黏附和信号转导中具有起始作 用。
整 合 素 结 构 模 型
Linear structure of FAK
(2)受体酪氨酸蛋白激酶
PDGF一类的跨膜受体,可磷酸化靶蛋 白的酪氨酸,因此称为受体酪氨酸蛋白激 酶(receptor tyrosine kinase,RTK)。
蛋白激酶与磷酸酶ppt课件

2019/9/4
2
磷酸化修饰的氨基酸
2019/9/4
3
蛋白质的磷酸化修饰
导致受体蛋白的激活或失活; <2% 蛋白质的磷酸化修饰发生在Tyr残基上; 大部分的早期信号转导是由酪氨酸蛋白激酶介导
的(protein tyrosine kinase, PTKs); Ser/Thr磷酸化修饰多发生于信号转导级联反应
18
蛋白质磷酸酶
丝氨酸/苏氨酸蛋白磷酸酶 (PP1, PP2A); 酪氨酸蛋白磷酸酶 (PTP1B); 双特异性蛋白磷酸酶 (DUSP1); 脂质磷酸酶 (PTEN, SHIP)。
2019/9/4
19
Ser/Thr与Tyr蛋白磷酸酶
Ser/Thr磷酸酶 (PP1)
催化机制:
含有双核的金属离子中心; 通过水分子来进行亲核攻击; 磷酸基团不转移到磷酸酶上,
2019/9/4
13
双特异性蛋白激酶
MAPKK可同时催化MAPK激酶的T与Y的磷酸化 修饰:
磷酸化位点的三肽模体 — TXY ERK — TEY p38 — TGY JNK — TPY
MAPK是通过Thr和Tyr的双位点同时磷酸 化而被激活。
2019/9/4
14
磷酸肌醇循环
phosphatidylinositol
Fe
OO
HO
P O-Ser-substrate
2019/9/4
8
蛋白激酶的分类
酪氨酸蛋白激酶
受体激酶 (EGFR,FGFR,PDGFR) 非受体激酶 (JAK,Src,Abl)
丝氨酸/苏氨酸蛋白激酶 (PKC,MAPK,Plk,Rho Kinases)
双特异性蛋白激酶 (MAPKK)
蛋白质的转运与信号传导课件.ppt
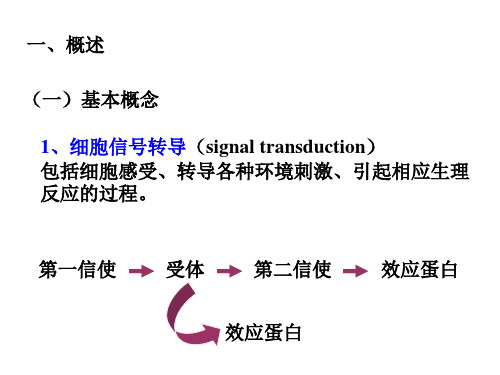
三、反应
同时,植物细胞信号转导系统在某些方面还保留了低等 原核细胞的信号转导机制,例如植物激素乙烯受体ETR1 与细菌双组份信号转导系统之间具有极大的相似性。
二、植物细胞信号转导过程
植物细胞的信号转导过程可以简单概括为: 刺激与感受——信号转导——反应三个重要的环节。
(一)刺激与感受 受体
(二)信号转导
此过程相当复杂,主要包括胞外信号的跨膜转换、细 胞内第二信使系统和信号的级联放大以及蛋白质的可 逆磷酸化。
1、信号跨膜转换
(1)通过离子通道连接受体跨膜转换信号
离子通道(ion channel)是存在于膜上可以跨膜转运 离子的一类蛋白质。
(2)酶促信号直接跨膜转换
该过程的跨膜信号转换主要由酶连受体来完成。
cell
胞外环境信号:指机械刺激、磁场、辐射、温度、风、 光、CO2、O2、土壤性质、重力、病原因子、水分、营养 元素、伤害等影响植物生长发育的重要外界环境因子。
胞间信号:指植物体自身合成的、能从产生之处运到别 处,并对其他细胞作为刺激信号的细胞间通讯分子,通 常包括植物激素、气体信号分子NO以及多肽、糖类、细 胞代谢物、甾体、细胞壁片段等。
3、第二信使(second messenger)
又称次级信使,是指细胞感受胞外环境信号和胞间信 号后产生的胞内信号分子,从而将细胞外信息转换为 细胞内信息。
一般公认的细胞内第二信使有钙离子(Ca2+)、肌 醇三磷酸(inositol 1,4,5-trisphosphate,IP3)、二酰 甘油(1,2-Diacylglycerol,DG)、环腺苷酸 (cAMP)、环鸟苷酸(cGMP)等。
•高特异性 •高的亲合力 •高饱和性 •可逆性 •通过磷酸化(与动物细胞信号传 导的区别)
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
DNA依赖性蛋白激酶(DNA-dependent protein kinaes,DNA-PK)可磷酸 化许多核蛋白,包括核受体、转录因子、 DNA拓扑异构酶和RNA聚合酶Ⅱ等。
• 与蛋白质或多肽底物结合; • 与磷酸供体ATP/GTP结合; • 转移磷酸基到底物相应的氨基酸残基上。
2. 调节结构域/亚基 同源性较低。 作用: 调节酶的活性; 靶向作用,与酶的亚细胞定位有关。
(二)蛋白激酶的种类
真核细胞的蛋白激酶可分为五类: ① 丝氨酸/苏氨酸蛋白激酶 ② 酪氨酸蛋白激酶 ③ 组/赖/精氨酸蛋白激酶 ④ 半胱氨酸蛋白激酶 ⑤ 天冬氨酸/谷氨酸蛋白激酶
蛋白激酶(protein kinase,PK):
是一类磷酸转移酶,其作用是将 ATP 的 - 磷酸基转移到底物特定的氨基酸残基 上,使蛋白质磷酸化, 发挥其生理生化功 能。
(一)蛋白激酶的结构 共同的结构特征:
• 保守的催化结构域/亚基 • 调节结构域/亚基 • 其他功能结构域
PKC
1. 催化结构域/亚基 催化核心含有12个高度保守的亚区。 功能:
CH1 CH2
K inase dom ain
N -te r LIM S
M igfilin
FERM PH FERM
M ig-2
p a x illin
In te g rin s
ECM
G enerously provided to the C M C by C ary W u (Apr-03
(9)DNA依赖性蛋白激酶
(7)核糖体S6激酶(S6K)
包括S6KⅠ和S6KⅡ,能催化核糖体 S6蛋白磷酸化。
(8)整合素连接激酶
整合素连接激酶(intergrin-linked kinase,ILK)可直接磷酸化PKB/Akt, 其活性依赖PI3K。
PIN C H , ILK dom ain structures & interacting partners
C1是膜结合区,与佛波酯、DAG和PS结合 有关。
C2是Ca2+结合位点。
• 催化区:
C3是ATP结合位点。
C4是底物蛋白结合位点。
PS DAG
Ca2+
ATP
PKC底物:
• 参与信号转导的底物,如表皮生长因子 受体、胰岛素受体、T细胞受体(TCR)、 Ras、GTP酶活化蛋白等;
• 参与代谢调控的底物,如膜上的通道和 泵;
活化需要Ca2+ 、DAG和PS。
B组-新型PKC (new PKC,nPKC):、、 (L) 、、 和 。 活化不需要Ca2+。
C组-非典型PKC (atypical PKC,aPKC): 和 / 。 活化不需要Ca2+和DAG。
各亚型均由一条肽链组成,分为四个保 守区C1~C4和五个可变区V1~V5。 • 调节区:
N c k -2
SH3 SH3 SH3
SH2
PIN C H
LIM 5 LIM 4 LIM 3 LIM 2
LIM 1
IL KΒιβλιοθήκη ppANK ANK ANK ANK PH
RTK
C H -ILK B P/actopaxin p a rv in s /a ffix in
N -te r
Actin C ytoskeleton Filam in
1. 丝氨酸/苏氨酸蛋白激酶
丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase,S/T-PK)催化 丝氨酸/苏氨酸的羟基磷酸化。
NH HCCH2 OH OC
NH
O
HCCH2 OPO
OC
O
(1)蛋白激酶A(protein kinase A,PKA)
即cAMP依赖性蛋白激酶。
• 调节基因表达的底物,如转录因子、翻 译因子、S6K、Raf激酶等。
PKC分布:
• 广泛分布于各组织的胞质,以Ca2+依赖 的形式从胞质中移位到细胞膜上,此过程 称之为转位。PKC转位是其活化的标志。
• 佛波酯(TPA or PMA)是一种促癌剂,其 结构与DAG相似,可持续活化PKC,促进 细胞增殖。
第五章
蛋白激酶、蛋白磷酸酶 与信号转导
【目的要求】
1.掌握蛋白激酶的概念及其催化的蛋白质磷酸 化反应;丝氨酸/苏氨酸蛋白激酶的主要种类及 其激活剂;PKC的分类及其特点;酪氨酸蛋白 激酶的概念、分类及其重要的结构域;受体酪 氨酸蛋白激酶的概念;蛋白磷酸酶的概念及分 类。
【目的要求】
2.熟悉受体酪氨酸蛋白激酶的分类;双重特异 性蛋白激酶;蛋白激酶在信号转导中的作用; 丝氨酸/苏氨酸蛋白磷酸酶的生物学作用;酪氨 酸蛋白磷酸酶的分类及其在信号转导中的作用; 双重特异性蛋白磷酸酶。
乙酸豆蔻佛波
(3)钙/钙调素依赖性蛋白激酶(CaMK)
包括肌球蛋白轻链激酶(myosin light chain kinase,MLCK)、磷酸化酶 激酶、CaMKⅡ等。
(4)CMGC组蛋白激酶
• 脯氨酸依赖性激酶(proline depedent kinase,PDK)
• 酪蛋白激酶Ⅱ(casein kinase Ⅱ ,CK Ⅱ )家族
PDK:
细胞周期素依赖性蛋白激酶(cyclin depedent kinase,CDK)家族
丝裂原活化蛋白激酶(mitogenactivated protein kinase,MAPK)家族
糖原合成酶激酶3(glycogen synthetase kinase 3,GSK3)
CDK样激酶(CDK-like kinase, CLK)家族
全酶存在胞浆,被cAMP激活后,催化亚 基可① 调节代谢;②调节离子通道;③调 节其他信号转导途径的蛋白;④ 进入细胞 核调节基因表达。
(2)蛋白激酶C
即Ca2+和磷脂依赖的蛋白激酶,受 Ca2+ 、DAG和PS激活。
PKC有11种亚型:
A组-典型PKC (classical PKC,cPKC): 、Ⅰ、Ⅱ和
MAPK途径:
EGF途径
(5)蛋白激酶G
即cGMP依赖性蛋白激酶( cGMP – dependent protein kinase,PKG),以 cGMP 为变构剂,在脑和平滑肌中含量较 丰富。
(6)G蛋白偶联受体激酶
有肾上腺素受体蛋白激酶( adrenergic receptor kinase, -ARK)、 -ARK相关激酶和视紫红质激酶等。
• 与蛋白质或多肽底物结合; • 与磷酸供体ATP/GTP结合; • 转移磷酸基到底物相应的氨基酸残基上。
2. 调节结构域/亚基 同源性较低。 作用: 调节酶的活性; 靶向作用,与酶的亚细胞定位有关。
(二)蛋白激酶的种类
真核细胞的蛋白激酶可分为五类: ① 丝氨酸/苏氨酸蛋白激酶 ② 酪氨酸蛋白激酶 ③ 组/赖/精氨酸蛋白激酶 ④ 半胱氨酸蛋白激酶 ⑤ 天冬氨酸/谷氨酸蛋白激酶
蛋白激酶(protein kinase,PK):
是一类磷酸转移酶,其作用是将 ATP 的 - 磷酸基转移到底物特定的氨基酸残基 上,使蛋白质磷酸化, 发挥其生理生化功 能。
(一)蛋白激酶的结构 共同的结构特征:
• 保守的催化结构域/亚基 • 调节结构域/亚基 • 其他功能结构域
PKC
1. 催化结构域/亚基 催化核心含有12个高度保守的亚区。 功能:
CH1 CH2
K inase dom ain
N -te r LIM S
M igfilin
FERM PH FERM
M ig-2
p a x illin
In te g rin s
ECM
G enerously provided to the C M C by C ary W u (Apr-03
(9)DNA依赖性蛋白激酶
(7)核糖体S6激酶(S6K)
包括S6KⅠ和S6KⅡ,能催化核糖体 S6蛋白磷酸化。
(8)整合素连接激酶
整合素连接激酶(intergrin-linked kinase,ILK)可直接磷酸化PKB/Akt, 其活性依赖PI3K。
PIN C H , ILK dom ain structures & interacting partners
C1是膜结合区,与佛波酯、DAG和PS结合 有关。
C2是Ca2+结合位点。
• 催化区:
C3是ATP结合位点。
C4是底物蛋白结合位点。
PS DAG
Ca2+
ATP
PKC底物:
• 参与信号转导的底物,如表皮生长因子 受体、胰岛素受体、T细胞受体(TCR)、 Ras、GTP酶活化蛋白等;
• 参与代谢调控的底物,如膜上的通道和 泵;
活化需要Ca2+ 、DAG和PS。
B组-新型PKC (new PKC,nPKC):、、 (L) 、、 和 。 活化不需要Ca2+。
C组-非典型PKC (atypical PKC,aPKC): 和 / 。 活化不需要Ca2+和DAG。
各亚型均由一条肽链组成,分为四个保 守区C1~C4和五个可变区V1~V5。 • 调节区:
N c k -2
SH3 SH3 SH3
SH2
PIN C H
LIM 5 LIM 4 LIM 3 LIM 2
LIM 1
IL KΒιβλιοθήκη ppANK ANK ANK ANK PH
RTK
C H -ILK B P/actopaxin p a rv in s /a ffix in
N -te r
Actin C ytoskeleton Filam in
1. 丝氨酸/苏氨酸蛋白激酶
丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase,S/T-PK)催化 丝氨酸/苏氨酸的羟基磷酸化。
NH HCCH2 OH OC
NH
O
HCCH2 OPO
OC
O
(1)蛋白激酶A(protein kinase A,PKA)
即cAMP依赖性蛋白激酶。
• 调节基因表达的底物,如转录因子、翻 译因子、S6K、Raf激酶等。
PKC分布:
• 广泛分布于各组织的胞质,以Ca2+依赖 的形式从胞质中移位到细胞膜上,此过程 称之为转位。PKC转位是其活化的标志。
• 佛波酯(TPA or PMA)是一种促癌剂,其 结构与DAG相似,可持续活化PKC,促进 细胞增殖。
第五章
蛋白激酶、蛋白磷酸酶 与信号转导
【目的要求】
1.掌握蛋白激酶的概念及其催化的蛋白质磷酸 化反应;丝氨酸/苏氨酸蛋白激酶的主要种类及 其激活剂;PKC的分类及其特点;酪氨酸蛋白 激酶的概念、分类及其重要的结构域;受体酪 氨酸蛋白激酶的概念;蛋白磷酸酶的概念及分 类。
【目的要求】
2.熟悉受体酪氨酸蛋白激酶的分类;双重特异 性蛋白激酶;蛋白激酶在信号转导中的作用; 丝氨酸/苏氨酸蛋白磷酸酶的生物学作用;酪氨 酸蛋白磷酸酶的分类及其在信号转导中的作用; 双重特异性蛋白磷酸酶。
乙酸豆蔻佛波
(3)钙/钙调素依赖性蛋白激酶(CaMK)
包括肌球蛋白轻链激酶(myosin light chain kinase,MLCK)、磷酸化酶 激酶、CaMKⅡ等。
(4)CMGC组蛋白激酶
• 脯氨酸依赖性激酶(proline depedent kinase,PDK)
• 酪蛋白激酶Ⅱ(casein kinase Ⅱ ,CK Ⅱ )家族
PDK:
细胞周期素依赖性蛋白激酶(cyclin depedent kinase,CDK)家族
丝裂原活化蛋白激酶(mitogenactivated protein kinase,MAPK)家族
糖原合成酶激酶3(glycogen synthetase kinase 3,GSK3)
CDK样激酶(CDK-like kinase, CLK)家族
全酶存在胞浆,被cAMP激活后,催化亚 基可① 调节代谢;②调节离子通道;③调 节其他信号转导途径的蛋白;④ 进入细胞 核调节基因表达。
(2)蛋白激酶C
即Ca2+和磷脂依赖的蛋白激酶,受 Ca2+ 、DAG和PS激活。
PKC有11种亚型:
A组-典型PKC (classical PKC,cPKC): 、Ⅰ、Ⅱ和
MAPK途径:
EGF途径
(5)蛋白激酶G
即cGMP依赖性蛋白激酶( cGMP – dependent protein kinase,PKG),以 cGMP 为变构剂,在脑和平滑肌中含量较 丰富。
(6)G蛋白偶联受体激酶
有肾上腺素受体蛋白激酶( adrenergic receptor kinase, -ARK)、 -ARK相关激酶和视紫红质激酶等。