遗传标记在植物遗传育种中的应用 ppt课件
《分子标记辅助育种》课件
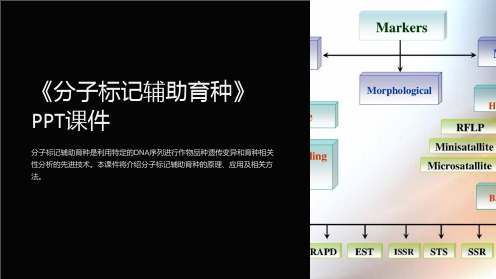
基于分子标记的品种鉴定方法
SSR
利用SSR标记的特定序列,对植 物品种进行指纹图谱构建。
SNP
AFLP
通过对SNP位点进行基因型检测, 进行品种鉴定和鉴别。
通过AFLP分析,对DNA片段进 行电泳分离,进行品种鉴定。
基于分子标记的遗传分析方法
1
关联分析
通过比较基因型和表型数据,寻找遗传
遗传图谱
2
基于分子标记的基因组选择方法
SNP array
利用高通量芯片分析大规模SNP位点,实现基因组宽选择。
Genotyping-by-sequencing
通过测序分析SNP位点,实现高密度基因组选择。
Marker-assisted recurrent selection
基于标记信息辅助进行循环选择,加快育种进程。
意义
分子标记辅助育种能解决传统育种中的难题,如长时间、低效率、受到环境因素等限制,同 时提高育种效率和精度。
优势
该技术可以加快育种进程、提高选择准确性、节省育种材料和资源,并可对育种过程进行精 确控制。
分子标记的种类及原理简介
SSR
简单重复序列,通过PCR扩增 的缺陷突变位点。
SNP
单核苷酸多态性,常见基因组 标记,便于高通量测定。
《分子标记辅助育种》 PPT课件
分子标记辅助育种是利用特定的DNA序列进行作物品种遗传变异和育种相关 性分析的先进技术。本课件将介绍分子标记辅助育种的原理、应用及相关方 法。
什么是分子标记辅助育种?
定义
分子标记辅助育种是一种利用特定DNA序列的基因标记来辅助育种的技术,通过分析和检 测基因标记,可以更快、更准确地选育出优良的品种。
AFLPபைடு நூலகம்
SSR标记在植物遗传育种中的应用

S R标记 具 有 的主 要优 点 :① 数 量丰 富 ,均 匀 、随机 、广 泛地 分布 于植 物基 因组 中 , S
覆 盖整 个基 因组 ;② 每个 位 点均 有许 多等 位形 式 , 多态性 丰 富 ,信息 含量 高 ;③ 以孟 德
尔方 式 遗传 ,呈共 显 性 ,能够 鉴别 出纯合 基 因型和 杂 合基 因型 。揭示 等位 位 点的差 异 ,
遗 传 学家 It1Jt研 究 了几个 S R 位 点上 的l个 等位 基 因 时发 现 ,这 些序 列 中有 3个 i ̄ I y t ]u S 2
2 8
小
麦
研
究
3 卷 2
( AAT 或4 ( )n 个 AGA )n 核苷 酸 重复 的序 列 , T 并将其 命 名 为Mirstlt即微 卫星 。 S c0aele i SR 这一 技 术一 经 问世 ,便 很 快在 动植 物 的遗 传分 析 中得到 广 泛应用 。 1 2 S R标 记 的原 理 . S 由于基 因组 中某一 特 定 的微卫 星 的侧 翼序 列通 常都 是保 守性 较 强 的单一 序列 ,因而 可 以将 微 卫 星侧 翼 的 DN 片段 克 隆 、测序 ,然后 根据 微卫 星 的侧 翼序列 就 可 以人工 合 A 成 引物 进 行 P R扩增 ,从而 将 单个微 卫 星位 点扩 增 出来 。由于单 个微 卫星 位 点重 复单 元 C
由于 S R标记 具 有诸 多优 点 , S 使得 其在 构 建植物 遗 传 图谱 、 传 多样性 、 因定位 、 遗 基 抗病 基 因的筛 选 等方 面广 泛 应用 。 2 1 建 遗 传图 谱 .构
S R是共 显 性标 记 ,多态 性 高 ,适宜 构 建 高密 度 的遗 传 图谱 。 白 R dr “ 发 表 了 S oe 等
《遗传与作物育种》课件

04
作物育种实践
水稻育种实践
1 2
杂交育种
利用不同品种的水稻进行杂交,通过选择优良后 代,培育出具有优良性状的水稻新品种。
诱变育种
通过辐射、化学诱变等方法,使水稻基因发生突 变,进而筛选具有优良性状的突变体。
3
分子标记辅助育种
利用分子标记技术,辅助选择具有优良性状的水 稻基因型,提高育种效率和准确性。
体,再从中选择和培育。
群体改良
03
利用地理隔离或人工创造的隔离条件,使不同品种在群体内混
合授粉,产生遗传变异,从中选择和培育。
现代育种技术
分子标记辅助育种
利用分子标记技术识别与目标性 状相关的基因,实现快速、准确 的品种选育和遗传改良。
基因工程育种
通过基因克隆、转基因等技术手 段,将具有优良性状的外源基因 导入作物中,实现定向遗传改良 。
表型与表现型
表型是指生物体的形态、结构、生理 和行为特征;表现型则是表型在特定 环境下的表现形式。
02
作物育种原理
作物改良的目标
提高产量
通过改良作物的遗传特性,提 高单产和总产量,满足不断增
长的食物需求。
增强抗逆性
提高作物对环境胁迫的抗性, 如抗旱、抗寒、抗病虫害等, 以适应各种不利条件。
改善品质
,为人类提供稳定的食物来源。
03
应对气候变化的重要手段
遗传资源具有适应不同环境条件的能力,通过保护和利用遗传资源,可
以培育出适应气候变化的新品种,提高农业生产应对气候变化的能力。
遗传资源的保存
原地保存
在原生地或近原生地自然生长的遗传资源称为原地保存。 这种保存方式能够保持遗传资源的自然生态环境,有利于 种质生态适应性的保持。
遗传变异和育种下课件

基因工程育种
总结词
基于基因技术的育种方法 。
描述1
基因工程育种是利用基因 重组、基因编辑等基因技 术对生物体进行精确的遗 传改良,以创造具有优良 性状的新品种的方法。
描述2
该方法可以实现跨物种基 因转移、目标基因的高效 表达、基因功能的精确调 控等。
3
应用前景
转基因技术在作物育种上有广阔的应用前景,可 以培育出抗病、抗虫、抗旱等性能更强的作物。
畜禽育种在食品安全中的地位
育种目标
畜禽育种以提高肉质、增加产量、改善疾病抗性等为目标,以确 保食品的安全和有效供给。
食品安全
通过育种技术可以减少畜禽疾病的发生,降低兽药使用量,从而提 高食品安全水平。
经济效益
遗传变异和育种课件
CONTENTS
目 录
• 遗传变异和育种概述 • 遗传变异原理与方法 • 育种技术与方法 • 各类作物的育种实践 • 育种技术前沿与展望 • 案例分析与讨论
01 遗传变异和育种概述
CHAPTER
遗传变异定义与类型
定义:遗传变异是指基因型或表型在种群中个体间存在的 差异,是生物进化的原材料。
速育种进程。
分子标记辅助育种在作物品 质改良、抗逆性育种以及畜 禽生产性能改良等方面具有
广泛应用。
04 各类作物的育种实践
CHAPTER
谷物育种
01
02
03
选育高产优质品种
利用遗传变异原理,选育 出产量高、品质优、抗逆 性强的谷物品种,满足日 益增长的食品需求。
改良抗病抗虫性状
通过基因编辑等技术手段 ,将抗病抗虫基因导入谷 物品种中,提高其对生物 胁迫的抗性。
分子标记技术及其在园林植物遗传育种中的应用 精品

分子标记技术及其在植物遗传育种中的应用近年,随着生物技术的快速发展,分子标记技术在诸多领域得到应用,尤以农业、医药业、畜牧业等行业应用得最多。
分子标记是指以生物大分子的多态性为基础的遗传标记。
分子标记的出现,使植物育种的“间接选择”成为可能,大大提高了遗传分析的准确性和选育种的有效性,因而在遗传育种领域愈来愈受到重视。
在遗传学研究中广泛应用的DNA分子标记已经发展了很多种,一般依其所用的分子生物学技术大致分为两大类:一类是以Southern杂交技术为核心的分子标记(如RFLP),此类被称为第一代分子标记;以PCR技术为核心的分子标记(如STS、RAPD、AFLP、SSR等)称为第二代分子标记,单核苷酸多态性(SNP)标记称为第三代分子标记,这也是以PCR技术为基础的分子标记技术。
现分别介绍其原理及在植物育种上的应用。
1分子标记在植物育种上的特点分子标记育种(molecular mark-assist selection,MAS)是借助分子标记在DNA水平上对遗传资源或育种材料进行选择,对作物产量,品质和抗性等综合性状进行高效改良,并针对目标性状基困连锁进行优良植株筛选,是现代分子生物学与传统遗传育种相结合的新品种选育方法。
与传统育种相比分子标记的优势是:(1)传统育种通过性状间接筛选目的基因,分子标记则通过直接与目的基因连锁进行筛选,因此,后者比前者准确,特别是在一些表现型与基因型之间对应关系较差时的筛选,(2)传统育种需要在成熟期才能筛选,分子标记筛选则可以不受植物生长发育期的限制,在苗期就可以筛选,而且不影响植株生长,(3)传统方法一次只能标记一个基因,分子标记筛选则可以同时筛选多个目的性状基因,(4)分子标记筛选利用了控制单一性状的多个等位基因,避免了传统育种通过表现型而获得不纯植株的缺陷;(5)分子标记筛选样品用量少,可以进行非破坏性筛选,从而加速育种进程,提高育种效率。
2常用分子标记的技术及其在植物育种上的应用2.1限制性内切酶片段长度多态性(restriction fragment length polymorphism,RFLP,简称限制片段长度多态性)RFLP是以分子杂交技术为基础的标记技术,其原理是碱基的突变、缺失、重排或是一段DNA的重排或插入,导致限制性内切核苷酸酶的酶切位点分布发生改变,得到的切割片段在数量和长度上不同,从而产生多态性。
分子遗传标记及其在作物诱变遗传育种中的应用

知识介绍分子遗传标记及其在作物诱变遗传育种中的应用项友斌 高明尉(浙江农业大学原子核农业科学研究所 杭州 310029) 介绍了分子遗传标记中限制性片段长度多态性(RFLP)、随机扩增多态DN A(RAPD)和多拷贝重复串联序列DN A微卫星与小卫星的基本概念和原理,以及它们在作物诱变遗传育种和品种改良中的应用。
关键词:R FLP R AP D 重复DN A序列 诱变育种 分子标记 植物育种中,一般都是通过亲本杂交与回交,从供体品种或野生种中将期望性状转到受体品种,达到改良的目的。
这种常规育种方法,往往要创造分离群体,经表型性状筛选,实现育种目标,需要经历较长时间。
并且,在一些多基因控制的数量性状、隐性基因控制的性状以及为获得对某一病原菌抗性需要在同一基因型中积累多个抗性基因的遗传改良中,采用常规方法往往难以奏效,而利用分子标记则易于取得成功。
因为分子标记把表型与基因位点连在一起,使一些难以在表型中区分的基因在较短时间里即可鉴别出来,从而提高了选择效率。
1.形态标记、生化标记和分子标记利用可分辨的表型性状作为遗传标记,在不同植物中已构建了遗传图谱。
但直到最近,用于作图的遗传标记还是那些影响形态性状的标记,包括花色、矮秆、白化苗及形态改变。
由于表型标记并非完善的指标,它不仅取决于遗传物质,还受生长环境的影响,许多环境因素有遮蔽基因的作用;有些表型标记作用太大,可能致死;而且形态标记的数量有限;故至今形态标记遗传图谱绘制的进展极其缓慢。
同功酶是一种中性的生化遗传标记,在一些植物的制图上已获得较大成功。
但同功酶在数量上也是有限的,它的表达经常被限制在一定发育时期和特定的组织中,极大地限制了它的广泛利用。
现在人们所说的分子标记,往往都是指DN A标记。
1975年,遗传学家Crodz icker等人首次描述了DN A限制性片段长度多态性(RFLP),1980年,建立了第一套人类根据DN A标记的遗传图谱,由此引发了建立其它真核生物DN A分子的标记遗传图谱。
分子标记在植物遗传育种中的应用

分子标记在植物遗传育种中的应用EM摘要N: 分子标记是继形态标记O细胞标记和生化标记之后发展起来的一种新的较为理想的遗传标记F已被广泛地应用于生命科学研究的各个领域P在植物遗传育种中F分子标记主要用于基因组图谱构建O基因定位O辅助标记选择O种质资源评价O基因克隆O杂种优势预测O杂交育种及跟踪育种过程等方面P文章主要介绍了分子标记在植物遗传育种中的应用原理及分析方法F并对其应用前景进行了展望PM关键词N 分子标记K植物遗传育种K遗传标记优良品种是当今农业经济发展的基础资源F对目标性状#如丰产O优质O抗逆等$的选择是新品种选育过程的中心环节P 传统的育种方法主要是根据植物在田间的表现进行评价和选择P 但由于表型性状不仅取决于遗传组成F也受控于环境条件F有时环境条件的影响可遮盖植株在基因型上的差异F因此仅根据表型进行选择F效果不够理想P特别是对受多基因控制的数量性状的选择F更难做到准确P虽然育种学已建立了一套完整的选择程序F并在农业生产上培育出了许多高产O优质O抗逆的新品种F然而传统的育种方法仍存在周期长O预见性差O工作量大O工作效率低等问题P随着遗传学的发展F人们注意到利用易于鉴别的遗传标记#R6=6398SA0T607$来进行辅助选择F可提高选择效果P遗传标记已逐渐成为植物遗传育种的重要工具P尤其是分子遗传学的发展及分子标记技术的建立F使作物遗传育种进入了一个新阶段P新技术与传统方法相结合F有可能解决目前育种中一些重要环节上的主要难题F从而大大加速育种工作进程P分子标记已在遗传图谱的构建O辅助标记选择O基因克隆等方面显示出了非常诱人的前景P- 应用分子标记构建基因组图谱基因组图谱是遗传研究的重要内容F又是种质资源O育种及基因克隆等许多应用研究的理论依据和基础P因此F基因组图谱已成为当今生命科学的重要研究领域P基因组图谱包括以染色体重组交换为基础的遗传图谱#R6=6398SA+$和以U2? 的核苷酸序列为基础的物理图谱#V4W798A’SA+$P数十年来F许多遗传学家利用形态标记O生化标记和传统的细胞遗传学方法F为构建各种主要作物的遗传图谱进行了大量工作F并取得了一定的进展P但是由于形态标记和生化标记数目少F特殊遗传材料培育困难及细胞学工作量大F因而除极少数作物#如玉米O番茄$外F在分子标记出现之前F大多数作物还没有一个较为完整的遗传连锁图F极大地限制了遗传学理论研究和应用研究的进展P 进入.,年代以来F分子标记的迅速发展F大大促进了遗传连锁图的构建P目前主要农作物O果树O蔬菜等的XCYV7OX?VU7遗传图谱已相继建立M-ZINP利用分子标记构建遗传图谱的理论基础是染色体的交换与重组P 两点测验和三点测验是其基本程序P由于作图群体的不断增大和标记数量的日益扩增F如今的遗传图谱构建已不得不计算机化了P 遗传图谱构建过程主要包括[-$选择和建立适合的作图群体K!$确立遗传连锁群KG$基因排序和遗传距离的确定P具体方法如下MJFHN[以分子标记筛选U2? 序列差异较大而又不影响后代育性的材料作为亲本F用具有多态性的分子标记#双亲在等位区段表现不同的带型$检测该双亲的分离群体#如C!代群体O回交O重组自交系O加倍单倍体等$单株U2?P如是共显性标记F对同亲本V-具有相同带型的个体赋值为-F同亲本V!具有相同带型的个体赋值为GF具有V-和V!带型的杂合个体赋值为!P 如是显性标记F因E M收稿日期N !,,,:--:!"M基金项目N 烟台师范大学博士科研起动经费资助项目#!,,,-!$M作者简介N 王永飞#-"L!\$F男F山西壶关县人F副教授F博士P主要从事植物遗传育种和分子生物学研究P不能区分显带的纯合个体和杂合个体!对有谱带的个体赋值为"#$$%$&或&&%&$’!谱带缺失的赋值为(#&&或$$’)统计各带型的个体数!两点测验是否连锁) 利用*+检验时!那些两点各自独立遗传而两点同时检验时偏离孟德尔比例的位点!说明存在连锁)或从第二个标记开始!检验是否与前一个标记协同分离)因为连锁分析建立在分子标记协同分离的程度上)根据分离资料!用最大似然估计标记位点间的重组率并转换成遗传图距#,-’)最后!同时考虑多个标记基因座位的共分离!即多点分析对标记进行排列!形成线性连锁图谱)生物染色体数目是特定的!标记数较少时!其连锁群可能比染色体数目多)随标记的增加!一些标记会与几个群上的标记连锁!而把它们连成一群) 当标记足够多时!标记连锁群的数目与染色体数目越趋接近!直至相等)标记图谱应是一个饱和的连锁图谱!即在所有染色体上每间隔"./ +.个交换单位或更近就有一个标记!使任何经典基因#包括数量性状基因’都包含在分子标记内)可见!分子标记连锁图本身对植物育种没有用处!只有当它与目标性状结合起来!才能发挥作用)这就必须把分子标记连锁图谱与染色体对应!有几种方法可供选用)如果将分子标记与已知染色体的同工酶标记%形态标记同时作图!根据分子标记与它们的连锁!很容易得到分子标记连锁的对应染色体)利用非整倍体如缺体%单体%三体或易位系%代换系等材料!也将分子标记连锁群与特定染色体对应常用的方法)如初级三体的染色体有(条!是正常+条染色体的"01倍!标记信号强度#如23456自显影强度’在三体上是二体上的"01倍)如水稻已获得全套初级三体!用连锁图谱上的某一分子标记探针同时与"+种三体78$ 杂交!比较"+种三体杂交带的强弱!此标记就位于信号较强的那个三体染色体上)此外!当原位杂交的灵敏度可以达到揭示单拷贝序列的杂交位点时!也可采用原位分子杂交将分子连锁图与染色体对应)此外!近年来利用分子标记在进行染色体物理图谱的构建%物理图谱与遗传图谱的比较%染色体不同部分的遗传重组值的差异等研究方面!也均取得了较大的进展)+ 应用分子标记定位基因基因定位就是将具有某一表型性状的基因定位于分子标记连锁图中!实现分子连锁图与经典连锁图的整合)+0" 质量性状基因的定位质量性状常由单个或几个基因控制!如果实的形状%色泽等)由于这些性状只受单基因控制!所以利用分子标记来定位%识别目标基因!及对具目标性状的植株进行直观的选择相对比较简单) 只要寻找与该目标基因紧密连锁的分子标记即可) 而且连锁的紧密程度越高!结果就越可靠)若已知该性状所属的染色体!则选择该染色体上的分子标记筛选)若该性状所属染色体未知!则从所有染色体上每隔一定距离!均匀抽取分子标记筛选)用这些在相对性状中表现差异的分子标记检测3+或其他作图分离群体单株78$!记载各带型个体数) 按上述作图统计方法!分析标记与目标性状的分离!计算重组值%图距并确定顺序)寻找与目标性状连锁的分子标记的有效方法可通过近等基因系#89&:;6<=9>;,?;>96!8@4’ABC)8@4是指通过多次回交筛选得到的%品系间差异主要在于某一目标性状的品系)由于基因连锁的结果!在回交导入目标性状基因的同时!与目标基因连锁的染色体片段也随之进入回交子代中)这种现象称为D连锁累赘E#4;>F&=9G:&==;>=’)经多代回交后得到的是除目标基因及其邻近区域外!已失去了供体其它基因型的品系!这个品系与轮回亲本就构成了一对近等基因系) 理论上!除了目标基因及邻近区不同外!其他区段应完全一致)近等基因系定位的原理正是鉴别和导入与目标基因连锁的分子标记)如果在近等基因系中检测出多态性!差异就必定在目标基因及邻近区域中) 找到的标记在这个范围内与目标基因连锁) 由于每个标记#如每个探针%引物对或随机引物’至多只分析供体亲本#HII9:9:J&:9>K!H5’%近等基因系和轮回亲本#29,L::9>KJ&:9>K!25’(个样品!与目标基因连锁的分子标记应在H5和8@4间带型一致!而在8@4和25间不同)因而每次可同时进行多个标记的筛选!但近等基因系的建立需多年回交!培育时间太长)在没有近等基因系可利用的情况下!集团分离分析法#ML?F9G69=:9=&>K&>&?N6;6!MO$’也是筛选多态性标记的有效方法APC) 从一对具有表型差异的亲本所产生的任何一种分离群体中!根据目标基因的表型分别选取一定量的植株!构成+个亚群或集团#如抗病群与感病群!其基因型在3+ 中抗病群22%2:!感病群为::Q在MR"中!抗病群为2:!感病群为::Q7S 中!抗病群为22!感病群为::’)将每群植株的78$ 等量混合!形成两个相对性状D基因增刊王永飞等W分子标记在植物遗传育种中的应用原理及现状 VUT池!"#$%$&’’()*因为每个+基因池!是由特定性状或基因组区域相同而其他非连锁区域完全随机的同类个体组成,所以在每个群体内,不管其他性状"基因)如何,在目标基因表型是一致的,而在两群间表型相反*在两类群间表现多态性的分子标记,遗传上与构建该类群所用性状基因座位相连锁* 当用双亲和两+基因池!作-./01分析时,表现多态性的针应是抗病集团与抗病亲本同带,感病集团与感病亲本同带*但在.2选择抗病单株时,因表型上不能区分纯合和杂合抗性单株,有可能选了杂合抗性个体进入+基因池!*此时,抗病集团可能有一条同感病集团的弱出现*当用-3041作分析,如标记与抗病基因处于相引相时,表现多态性的引物应是抗病集团与抗病亲本显相同带型,感病集团与感病亲本为零带型*如标记与抗病基因处于相斥相时,在无杂合抗病个体入选时,抗病集团与抗病亲本为零带型,感病集团与感病亲本显相同带型* 而在有杂合抗病个体入选时,他们均显感病样本带型,无法筛选* 此外,如在两集团中,分别混有性状基因与标记的交换型个体"在表型上难以区分)时,无论哪种筛选方法,两集团间均显相同带型,即使用有差异的探针或引物进行筛选,也鉴别不出来* 在组建集团时,个体越多,交换型个体混入的机会越大,会对探针或引物的筛选带来干扰* 用.5家系或对.2测交,鉴定.2 单株,组建集团时,剔去杂合体,使筛选更为可靠*总的看来,这种方法应用于-3041较-./01更方便,也较常规的-3041更有效6准确*它排除了环境及个体因素的影响,并让研究者把目标集中在植物基因组的特定区域*272 数量性状基因的定位作物许多重要的经济性状是受微效多基因控制的数量性状*89年代以前,数量遗传学家曾用经典的形态学和细胞学标记法来研究与标记相连锁的个别数量性状,试图定位数量性状基因":;<%=>=<=>?$ =@<>=(’A;1,:B/)*但由于这些标记数量太少以及技术上的局限性,极大地限制了对数量性状基因位点的深入研究*4C3 分子标记的建立和发展,使植物:B/作图及其定位方法进展很快,现已在番茄6玉米6水稻6大白菜等29多种作物上绘出D99多个数量性状的:B/图谱EFG*:B/定位法主要有以下几种*2727D 单标记定位法单标记定位法是利用线性回归原理,通过比较各标记基因型的量性状观测值的差异来定位:B/,并估计其效应的法*例如,设某一分子标记座位有两等位基因H 和I,某一数量性状基因座位有两个等位基因: 和J,两位点连锁遗传,交换值为K,两亲本基因型分别为HH::和HH:J,IIJJ*.D为HI:J,.D 形成H:,I:,H&,I&L种配子*雌雄结合产生DM个个体,共F种基NHH::,HHJJ,HI::,HI:J,HIJJ, II::,II:J和IIJJ* 分子标记HH,HI,II通过共显性标记"如-./01检测)加以区别,它们对应的数量性状可以度量*按分子标记将同标记的个体分为一组,可分成5组NHH 组6HI 组和II组*分别令这5组的数量性状平均值为OD,O2和O5,如ODP O2PO5,说明分子标记与:B/独立遗传,此时QP 97R*如5组数量性状平均值不等,OD,O2和O5作S测验,检测差异显著性,就可判断分子标记与:B/是否连锁ED9G* 但用单个标记研究:B/时,受遗传重组影响较大*如:B/与标记相距较远时,易因遗传重组失去连锁* 且两自交系在该位点为纯合基因或两亲本的:B/不同,但对该性状的作用相同时,可能根本检测不到某些实际存在的:B/,往往低估实际:B/数目*此外该法还存在以下不足之处NT不能估计:B/的确切位置UV不能确定标记是和一个还是多个:B/连锁UW:B/效应估计值偏低UX 检测:B/需要较多的个体,其效率较低*27272 区间作图法/<%Y$(和Z’=1=$>%EDDG针对单标记定位法的不足提出了区间作图法"[%=$@?<(I<&&>%\),它利用染色体上D个:B/两侧的各D对标记,建立个体数量性状观察值对双侧标记基因型指示变量的线性回归关系,以分离检验统计量中重组率和:B/效应* 统计原理是对基因组上两邻近分子标记间,按一定遗传距离的片段逐一分析*T按极大似然法计算表型效应的估计值UV 计算存在:B/时出现观察值的概率与无:B/时出现观察值的概率比"]YY@<=>’),用/]4 值"概率比以D9为底的对数)作为衡量:B/存在的尺度*这样检验同时用到了两个标记提供的信息,可将:B/ 与-./01之间重组类型鉴别出来,从而使检验灵敏度大大提高* 由于该方法假定一条染色体上只存在一个效应较大的:B/,而当一条染色体上存在2个或多个效应近似的:B/时,区间作图法难以逐一分辩:B/的效应,致使:B/定位不准确甚至有误*27275 复合区间作图法"^’I&’1>=$>%=$@?<(I<&_&>%\) 为了克服区间作图法的上述缺陷以及能利用多个遗传标记的信息,‘$%\ED2G提出了复合区间作cba 西北农林科技大学学报"自然科学版) 第 2F卷图法!以提高多个连锁"#$的辨别能力及其相应位置和效应估计的准确性%&’()认为!由于染色体的结构是线性的!当不存在连锁干扰和基因互作时! 一个标记基因型值的偏回归系数只受与其相邻区间内的基因的影响!与其它区域内的基因无关%虽然连锁干扰和基因互作可能存在!并且对作图有影响!但这种影响较小% 在此基础上!&’()将多元回归分析引入了区间作图法!实现了同时利用多个遗传标记的信息对基因组的多个区间进行多个"#$的同步检验%该法能减少剩余方法!提高"#$的发现率!并可降低测验统计量的显著水平%单标记定位虽难以精确标定"#$位置!但其发现能力仍可能是最高的%*种方法的同一性是主要的!方法间变异大多小于方法内变异% 在构建"#$图谱时!应同时使用几种方法!并优先标定共同发现的"#$!并可合并估计"#$的效应%* 分子标记在种质资源研究上的应用种质资源是发展农业生产和开展育种工作的物质基础% 种质资源的研究工作包括搜集+保存+鉴定和利用等一系列工作%分子标记在种质资源的鉴定+保存和利用研究中主要有以下几方面的用途,- 绘制品种.品系/的指纹图谱.01()’2321(41()/56 种质资源的遗传多样性及分类研究57 种质资源的鉴定和选择%*89 绘制品种.品系/的指纹图谱指纹图谱是鉴别品种+品系.含杂交亲本+自交系/的有力工具%它具有迅速.数小时至数天/+准确等优点%在目前市场经济迅速发展的形势下!指纹图谱技术在检测良种质量.真伪+纯度/!防止伪劣种子流入市场!保护我国名+优+特种质及育种品种的知识产权和育种家们的权益等方面!均具有重大意义%指纹图谱技术主要应满足两方面的要求%第一是分辨率高!多态性强5第二是重复性要强!即稳定可靠%作物品种指纹图谱目前主要有两种类型!一类是蛋白质电泳指纹图谱!其中包括同工酶和贮藏蛋白.如谷蛋白+醇溶蛋白/%这类指纹图谱在:;年代以前研究较多!目前仍不失其利用价值%另一类是<=> 指纹图谱!该项技术主要是:;年代之后发展起来的% 当前用来作<=> 指纹图谱的标记主要有?0$@A+小卫星<=>+微卫星<=>+>0$@A及?>@<A等% <=> 指纹的应用主要表现在以下几个面,-品种鉴定和种子纯度鉴定%借助<=> 指纹可精确地进行品种鉴定!尤其适于品种间有遗传差异而品种内一致的分析材料%例如无性繁殖的大多数果树的品种鉴定!即使是关系密切的品种也易借助<=> 指纹将其区别开来B9*C%6 知识产权保护%尽管目前国内尚未有此方面的报道!但D@EF.世界植物品种保护联盟/已将其列为鉴定植物品种真伪的手段!且已得到育种者们的共识!可能在不久的将来!指纹图谱会充分应用于品种产权保护中去%7种质资源的筛选和保存% 指纹技术在筛选和保存种质材料及保证遗传类型多样性等方面可能起重要作用%在资源保存方面!通过指纹图谱进行资源鉴别!可以避免在资源保存中经常发生的重复+混淆!避免同名异物和同物异名等现象%*8G 种质资源的遗传多样性及分类研究分子标记是检测种质资源遗传多样性.H’(’41I J1K’2A14L/的有效工具!目前主要用于以下M方面的研究,- 研究种质资源考察时取样量的大小+取样点的选择56 保护种质资源遗传完整性的最小繁种群体和最小繁种量的确定57 核心种质.NO2’IOPP’IQ41O(/筛选5R种质资源.含亲本材料/的分类%目前有关此类研究已有不少报道!并在某些方面取得了新的进展%*8G89 种质资源遗传多样性的评价种质资源是农作物育种的物质基础!对种质资源的可用性评价又是合理利用种质资源的前提% 经典的种质资源评价主要是依据对表型性状的观察进行的% 对质量性状而言!依据表型性状观察比较容易选到具有目标性状的单株或群体%而作物的许多重要农艺性状是由多基因控制的数量性状!改良数量性状而利用的种质一般不是单株!而往往是群体%描述群体特征常用性状平均数+方差+变异系数等指标作遗传多样性分析%分子标记特别是共显性分子标记!可以揭示整个基因组的变异!并分离出各种等位基因!从而可以计算群体中某位点等位基因数+多态位点比例.多态位点是指具有G个以上等位基因+且每个等位基因频率小于或等于;8::的位点% 测定的全部位点中!多态位点所占的比例称为多态位点比例/+等位基因平均数.等位基因总数与位点数之比/+杂合度X!YX为群体中某位点第X等位基因的频率!W为该位点等位基因目/+平均杂合度.\: 分子标记在追踪育种过程上的应用杂交育种是品种改良的最重要方法之一"杂交育种的过程!从理论上讲!双亲遗传物质的交换与重组是其基础"但在实践中一直缺乏直接了解这一过程的有效手段!因而大大限制了选育品种的效率和对杂交育种的深刻理解" 陈绍江等;%<=用5,738技术对大豆育种过程亲子遗传关系进行了研究" 他们以组合:<<%/中)&的亲本及其育成品系.)#.%为材料进行5,738分析!发现双亲在遗传上有较大差异!母本#父本分别有)(!+条特征带!其在子代能够重组!从而.)#.% 均具有双亲5,738标记的特征?但经多代选择!双亲5,738特征在育成的子代品系中的分布是不均衡的!.)更多地继承了母本:<<%/的绝大部分5,738特征!遗传上更近于母本!.% 则表现出倾向于父本" 这一结果与田间观察结果基本一致!说明利用分子标记追踪育种过程#探讨亲子遗传关系是可行的"& 分子标记在植物遗传育种中的应用前景分子标记为植物遗传育种开创了新的途径"它在基因组图谱构建#基因定位#辅助选择#种质资源评价#基因克隆#杂种优势预测!杂交育种及跟踪育种过程等方面已显示出非常诱人的前景!但要将分子标记广泛而深入地应用到遗传育种的各个领域!还应注意以下几个方面@A 寻找新型的分子标记?B简化分子标记技术!降低成本!实现检测过程的自动化?C改进分析基因组的方法和技术"相信随着分子标记技术的日臻完善!分子标记必将在植物育种中发挥越来越重要的作用";参考文献=; )= 沈法富!刘风珍!于元杰D分子标记在植物遗传育种中的应用;E=D山东农业大学学报!)&&F!%:1)2@:(G&)D;%= 贾继增D分子标记种质资源鉴定和分子标记育种;E=D中国农业科学!)&&+!%&1/2@)G)<D;(= 周奕华!陈正华D分子标记在植物学中的应用及前景;E=D武汉植物学研究!)&&&!)F1)2@F0G:+D;/= F/)D;0= 刘勋甲!尹艳!郑用琏D分子标记在农作物遗传育种中的运用及原理;E=D湖北农业科学!)&&:!1)2@%FG(%D;+= 张德水!陈受宜D34,分子标记#基因作图及其在植物遗传育种上的应用;E=D生物技术通报!)&&:!102@)0G%%D&:(%D;&= 冯宗云!荀琳!何萍!等D34,分子标记与作物量性状改良;E=D 西南农业学报!)&&:!))1增刊2@+FGF%Dmml 西北农林科技大学学报1自然科学版2 第 %&卷。
林木遗传育种(课件)

林木遗传育种学一、林木育种的概念及其特点(一)概念林木育种(forest tree breeding)是以遗传进化. 理论为指导,研究林木选育和良种繁育原理和技术的学科。
1、优良品种的选育选种:利用种内存在的丰富变异,在种的范围内进行选择。
种源选择(群体选择)林分选择(群体选择)优树选择(个体选择)引种:从外地或外国引进本地没有的树种。
杂交育种:通过人工有性杂交培育新品种。
多倍体育种:毛白杨三倍体、刺槐四倍体、欧洲白桦三倍体等诱变育种:(辐射育种、突变体筛选等);生物技术育种:(花药(花粉)培养、原生质体培养与融合、基因工程等)。
原生质体融合植物基因工程育种2、优良品种的繁育种子园:是由优树无性系或家系营建的,以生产优质种子为目的的特种林。
采穗圃:是由优树无性系营建的,用以生产优质种条(插穗和接穗)的繁殖圃。
3. 遗传测定(二)林木育种工作的特点1. 育种资源丰富2、育种周期长(1) 早期预测早――晚期生长相关分析形态――生长的早期鉴定生理――生长的早期鉴定法(2) 促进提早开花结实(3) 采用多种育种途径相结合,不断为生产提供改良程度逐步提高的繁殖材料。
3、树木可供研究利用的时间长,可以在遗传测定后进行再选择。
4、优良性状可以通过无性繁殖方法得到保存和利用5、多数为异花授粉树种,遗传基础广泛和稳定6、在多数情况下,选育和繁殖遗传基础广泛的林木品种或使用混合品种是适宜的。
二、林木遗传育种的回顾林木引种可追溯到2000年前,大规模的引种工作是从19世纪50年代由澳大利亚、新西兰等南半球国家引种松树开始的。
杨树、桉树、云杉、花旗松及落叶松等树种都已远远超越了各自的自然分布区,已成为国际性的重要造林树种。
种源试验是开展最早的林木遗传育种活动。
法国学者De Vilmorin于1823~1882年首先进行了欧洲赤松种源试验,随后法国、俄国、奥地利、瑞士等林学家对落叶松、云杉、松、橡等树种作了种源试验,证实了种内存在着明显的差异。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
生化标记的优点:
1.表现近中性,对植物经济性状一般没有大的不良影响。 2.直接反映了基因产物差异,受环境影响较小。
但目前可使用的生化标记数量还相当有限,且有 些酶的染色方法和电泳技术有一定难度,因此其实际 应用受到一定限制。
遗传标记在植物遗传育种中的应用
生化标记的应用
1.生化标记在品种纯度和真实性鉴定上的应用 2.生化标记在品种的亲缘关系和分类研究上的应用 3.生化标记在杂种优势的预测上的应用 4.生化标记在抗性鉴定上的应用 5.生化标记在品种鉴定上的应用
遗传标记在植物遗传育种中的应用
三 、生化标记
利用电泳技术对蛋白质、酶等生物大分子进行鉴定。主要 包括同工酶和等位酶标记。 同工酶:具有功能相同而结构和理化性质不同的一类酶。 等位酶:同一基因座上的不同等位基因编码的同工酶。
种子贮藏蛋白的电泳分析已广泛用于作物品种鉴别和种子纯 度鉴定。
遗传标记在植物遗传育种中的应用
(亚洲棉×陆地棉 )杂种F1与其亲本的形态特征比较
器官
粗细
主
茸毛
茎
腺体
(个/cm2)
形状
大小
色泽
叶
茸毛
片
叶柄
托叶
腺体 (个/cm2)
形状
大小
苞
苞外蜜腺
叶
围铃状态
腺体 (个/cm2)
大小
色泽
萼
片
上缘形状
腺体 (个/cm2)
花冠颜色
花
大小
花药颜色
花药数
A2 较粗 少而短
272.5 5裂 较大 深绿 少而短 长 披针型
遗传标记的类型
1.形态学标记(morphological marker) 2.细胞学标记(cytological marker) 3.生化标记(biochemical marker) 4.分子标记(molecular marker):DNA标记。
遗传标记在植物遗传育种中的应用
一、形态标记
即植物的外部形态特征。主要包括肉眼可见的 外部特征,如:矮秆、紫鞘、卷叶等;也包括色素、 生理特性、生殖特性、抗病虫性等有关的一些特性。
遗传标记在植物遗传育种中的应用
(1)核型和核型分析
核型----体细胞染色体在光学显微镜下所有可以测定的表型特 征的总称。
主要包括染色体的数目与形态结构特征两大方面的内容。 染色体的数目包括染色体的基数、多倍体、非整倍体、 超数染 色体(B染色体,是存在于许多有机体中的额外染色体及染色体断 片)。染色体的形态主要包括染色体的绝对大小和相对大小、着 丝点和次缢痕以及随体的数目和位置等特征。
遗传标记在植物遗传育种中的应用
核型分析:核型分析就是指对核型的各种特征进行定 量和定性的表述。核型分析是分析染色体数目和结构 变异的基本手段之一。它在杂种细胞的染色体研究和 基因定位,单个染色体的识别等方面具有重要意义。
遗传标记在植物遗传育种中的应用
遗传标记在植物遗传育种中的应用
遗传标记在植物遗传育种中的应用
44.7 心脏型
大 无 紧抱
75.3 小 浅绿 微波状
261.7
黄色,基部有红斑 较大 黄色 较多
G1 细 多而长
90.0 卵圆 小 灰绿 多而长 短 剑状
205.0 剑状 小 有 张开
182.0 较大 浅绿 线状齿长
157.5
A2A2G1G1 较粗 多而长
110.0 3-5裂 较大 灰绿 多而长 较短 披针型
(AD)1×(A2G1) 较粗 多而长
66.4
卵圆裂叶 较大 灰绿
较多而长 长
披针型
131.0 心脏型
较大 有 张开
53.4 较大
绿 宽齿长
128.0 深粉红色, 基部有红斑
大 黄色 较少
遗传表现
近似[AD]1, A2 偏向G1
A2, G1
[AD]1, A2, G1 近似[AD]1, A2 偏向G1 偏向G1 近似[AD]1, A2 近似[AD]1, A2
G1
近似[AD]1, A2 近似[AD]1, A2 近似[AD]1, G1 偏向G1
A2, G1
超亲
近似[AD]1, G1
[AD]1,G1互补
A2,G1
超亲型
超亲型 偏向A2 偏向G1
遗传标记在植物遗传育种中的应用
二、细胞学标记
即植物细胞染色体的变异。 1.染色体数目(单体、缺体、三体、四体)
2.染色体结构(缺失、重复、倒位、易位) 核型、带型
蚕豆的核型分析
2n=12, 染色体长度:大小 臂比:大小
遗传标记在植物遗传育种中的应用
核型分析内容:
1.染色体的数目 2.染色体的形态
核型分析的应用:
1. 植物的分类研究 2. 探讨物种的起源 3. 外源染色体的检测与验证杂种的真实性
遗传标记在植物遗传育种中的应用
遗传标记在植物遗传育种中的应用
遗传标记在植物遗传育种中的应用
遗传标记在植物遗传育种中的应用
遗传标记(genetic marker):指可追踪染色体、染色 体某一片段、某个基因座在家系中传递的任何一种遗 传特性。它具有两个基本特征,即可遗传性和可识别 性;因此生物的任何有差异表型的基因突变型均可作 为遗传标记。
遗传标记在植物遗传育种中的应用
(2)带型
特殊的染料、染色方法,使同一染色 体的不同区段呈现不同的染色效果-带型 ,各种分带技术C、G、Q、R、N的出现, 为染色体的精确鉴别提供了一条崭新的途 径。
遗传标记在植物遗传育种中的应用
人类染色体核型分析(Q带)
遗传标记在植物遗传育种中的应用
女
男
遗传标记在植物遗传育种中的应用
与形态标记相比,细胞学标记的优点是能进行一 些重要基因的染色体或染色体区域定位。但细胞学标 记材料需要花费较大的人力和较长时间来培育,难度 很大;同时某些物种对染色体变异反应敏感;还有些 变异难以用细胞学方法进行检测。因此到目前为止, 真正应用于作物遗传育种研究中的细胞学标记还很少。
64.0 披针型 较大 有或无 张开
95.3 较大 红色 宽齿长
181.8
(AD)1(无) 粗
少而长
0 5裂 大 绿 少而短 长 披针型
0 宽心脏型
大 有 紧抱
0 大 黄绿 宽有齿
0
ห้องสมุดไป่ตู้
浅粉红,基部有红斑
粉红色,基部有红斑
乳白色,基部无红斑
小
小
较大
白色
黄色
乳白色
遗少 传标记在植物较遗少 传育种中的应用多
优点:形态标记简单直观、经济方便。 缺点:(1)数量在多数植物中是很有限的。
(2) 表现易受环境影响。 (3)有一些标记与不良性状连锁。 (4)形态标记的获得需要通过诱变、分离纯 合的过程,周期较长。 形态标记在作物遗传育种中的作用是有限的。
遗传标记在植物遗传育种中的应用
遗传标记在植物遗传育种中的应用