酶学分析ALT分析Km值
酶的km

12.非竞争性抑制作用(noncompetitive inhibition):抑制剂不仅与游离酶结合,也可以与酶-底物复合物结合的一种酶促反应抑制作用。这种抑制使得Vmax变小,但Km不变。
4.活化能(activation energy):将一摩尔反应底物中的所有分子由基态转化为活化态所需要的能量。
5.活性部位(active site):酶中含有底物结合部位和参与催化底物转化为产物的氨基酸残基的部分。活性部位通常都位于蛋白质的结构域或亚基之间的裂隙或是蛋白质表面的凹陷部位,通常都是由在三维空间上靠得很近的一些氨基酸残基组成的。
9.米氏常数(Michaelis constant,)(Km):对于一个给定反应,导致酶促反应速度的起始速度(v0)达到最大反应速度(Vmax)一半时的底物浓度。
10.双倒数作图(double-reciprocal plot):也称之Lineweaver-Burk作图。一个酶促反应速度的倒数(1/v)对底物浓度的倒数(1/[s])的作图。X和y轴上的截距分别代表负米氏常数(Km)和最大反应速度(Vmax)的倒数。
中草药提高酶促反应速度的机制比较复杂。【举例说明】
6.抑制剂的影响
凡能降低酶活性的物质称为酶的抑制剂(没有专一抑制作用的因素或物质除外,如强酸、强碱等)。抑制作用可分为不可逆性抑制与可逆性抑制。
(1)不可逆性抑制。①概念:抑制剂与酶以共价键结合使酶失去活性,不能用透析法除去抑制剂使酶活性恢复,这种抑制称为不可逆性抑制。②抑制作用特点:不可逆性抑制,抑制剂浓度越高,抑制作用时间越长,抑制作用越强。③有机磷农药中毒:农药1059、敌百虫等有机磷农药中毒,主要是它们能特异地与胆碱酯酶活性中心的丝氨酸楚基结合,使酶失活。乙酰胆碱不能被胆碱酯酶水解,以致积蓄而引起迷走神经高度持续兴奋的中毒状态。
酶动力参数km、vmax和kcat值的计算
酶动力参数K m 、V max 和K cat 值的计算(以日立U3010为例)强玮 2014-3-31一、 K m根据酶促反应方程,即米-曼式氏方程(Michaelis-Menten equation ):V =(VmaxS)/(Km + S)当v=0.5Vmax 时,Km=[S],可见Km 等于酶促反应的初速率为最大速率Vmax 一半时的底物浓度。
Km 值一般在10-6~10-2mol/L 之间,单位一般为mM 或者uM 。
Km 只与酶的性质有关,而与酶的浓度无关。
由于Km 值与酶的浓度无关,所以计算时无需对蛋白进行定量,理论上诱导破碎的菌体上清(粗酶液)即可用来测酶活计算。
具体计算方法以前常用双倒数法,但是这种算法现在稍微好一点的SCI 杂志都不认可了,相对来说现在更准确和更常用的计算方法是直接对不同浓度的底物测得的酶活数据用米氏方程(Michaelis-Menten equation V =(VmaxS)/(Km + S))进行非线性回归(nonlinear regression ),拟合的方程中就给出了计算出的Km 值。
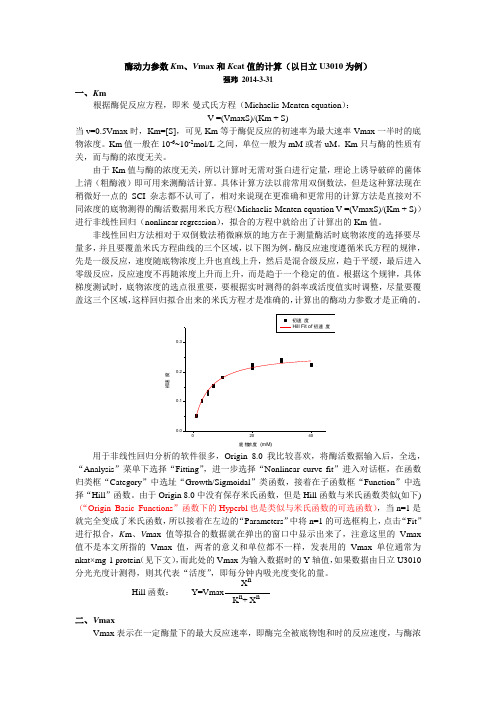
非线性回归方法相对于双倒数法稍微麻烦的地方在于测量酶活时底物浓度的选择要尽量多,并且要覆盖米氏方程曲线的三个区域,以下图为例,酶反应速度遵循米氏方程的规律,先是一级反应,速度随底物浓度上升也直线上升,然后是混合级反应,趋于平缓,最后进入零级反应,反应速度不再随浓度上升而上升,而是趋于一个稳定的值。
根据这个规律,具体梯度测试时,底物浓度的选点很重要,要根据实时测得的斜率或活度值实时调整,尽量要覆盖这三个区域,这样回归拟合出来的米氏方程才是准确的,计算出的酶动力参数才是正确的。
初速度底物浓度 (mM)用于非线性回归分析的软件很多,Origin 8.0我比较喜欢,将酶活数据输入后,全选,“Analysis ”菜单下选择“Fitting ”,进一步选择“Nonlinear curve fit ”进入对话框,在函数归类框“Category ”中选址“Growth/Sigmoidal ”类函数,接着在子函数框“Function ”中选择“Hill ”函数。
检验科常见酶学检测项目解读

检验科常见酶学检测项目解读酶学检测是临床实验室中常用的一种检测方法,通过对体内酶的活性及相关指标的检测,可以提供一系列与机体各个系统功能相关的重要生理信息。
本文将详细介绍检验科常见的酶学检测项目及其解读。
一、丙氨酸氨基转移酶(ALT)ALT是一种存在于细胞质中的酶,主要分布在肝脏、心肌、肾脏及骨骼肌等组织中。
当细胞受到损伤时,ALT会释放到血液中,因此,血清ALT水平的升高常常是肝功能损害的指标。
正常成人血清ALT水平一般较低,男性参考范围为10-40 U/L,女性为7-35 U/L。
超出正常范围的ALT水平可能表示肝脏炎症、肝细胞坏死或药物性肝损伤等。
二、天门冬氨酸氨基转移酶(AST)AST也是一种存在于多个组织中的酶,与细胞内的氨基酸代谢有关。
AST相对于ALT而言,其升高程度较低,因此AST常作为肝功能损害的辅助指标。
正常成人血清AST水平参考范围为10-34 U/L。
AST升高可见于多种疾病,如急性肝炎、心肌梗死、肝肿瘤或药物性损伤等。
三、乳酸脱氢酶(LDH)LDH是一种参与糖代谢的酶,广泛存在于多个组织和细胞中。
血清LDH主要源于组织损伤,LDH水平的升高常常提示组织坏死、炎症或缺血等情况。
正常成人血清LDH水平参考范围为109-245 U/L。
LDH的特异性较低,因此需要结合其他指标综合分析。
四、γ-谷氨酰转肽酶(GGT)GGT主要存在于肝脏、胆道和肾脏等组织中,具有很高的组织特异性。
血清GGT水平的升高常提示肝功能障碍、酒精性肝病、胆道梗阻或药物性肝损害等情况。
正常成人血清GGT水平参考范围为0-50 U/L,男性稍高于女性。
GGT的升高还可能与肥胖以及某些药物的使用有关。
五、碱性磷酸酶(ALP)ALP主要存在于肝脏、骨骼、胆道和肾脏等组织中,与细胞膜的磷酸化作用有关。
血清ALP水平的升高可提示肝胆道疾病、骨疾病或肿瘤扩散等情况。
正常成人血清ALP水平参考范围为45-125 U/L,男性稍高于女性。
碱性磷酸酶km值测定 实验报告

碱性磷酸酶km值测定实验报告碱性磷酸酶(alkaline phosphatase)是一种重要的酶类物质,在生物体内发挥着关键的生物学功能。
为了更好地了解碱性磷酸酶的性质和功能,我们进行了碱性磷酸酶的KM值测定实验。
实验中,我们首先准备了一系列不同底物浓度的反应液,并添加了一定浓度的碱性磷酸酶。
然后,通过测定一定时间内底物浓度的变化,我们可以得到不同底物浓度下反应速率的数据。
在实验中,我们选择了两种常用的底物——对硝基苯磷酸钠(p-NPP)和酚酞磷酸钠(BTP)来进行测定。
p-NPP是一种无色底物,在碱性条件下可以被碱性磷酸酶催化水解生成对硝基苯酚,其产物可以通过测定吸光度来确定反应速率。
而BTP是一种红色底物,在碱性条件下可以被碱性磷酸酶催化水解生成酚酞,其产物可以通过测定吸光度来确定反应速率。
通过实验数据的处理和分析,我们得到了不同底物浓度下的反应速率和底物浓度的关系。
通过拟合实验数据,我们可以得到反应速率与底物浓度之间的动力学关系。
根据麦克斯韦-玛尔蒙方程,我们可以得到碱性磷酸酶的KM值。
KM值是一个重要的酶学参数,它反映了底物与酶结合的亲和力。
KM值越小,表示底物与酶结合的亲和力越强,底物浓度较低时就能够达到较高的反应速率。
而KM值越大,表示底物与酶结合的亲和力较弱,需要较高的底物浓度才能达到较高的反应速率。
通过实验测定,我们可以得到碱性磷酸酶的KM值,进而了解其底物结合特性。
这对于研究碱性磷酸酶的功能和调控机制具有重要意义。
同时,通过比较不同底物的KM值,我们还可以了解底物对于碱性磷酸酶的亲和力差异,进一步揭示酶底物特异性的原理。
除了测定KM值,我们还可以通过其他实验方法来研究碱性磷酸酶的功能和特性。
例如,可以通过测定酶的最适温度和最适pH值来了解酶的适应范围。
此外,还可以通过测定酶的抑制剂对酶活性的影响来研究酶的抑制机制。
这些实验方法的综合应用可以更全面地了解碱性磷酸酶的生物学功能。
综上所述,碱性磷酸酶KM值的测定是研究酶特性和功能的重要手段之一。
酶促反应动力学 碱性磷酸酶Km值测定
❖ 在反竞争性抑制中,抑制剂必须在酶和底物结合以后 才能和酶结合形成EIS,其动力学特征是;Km和Vm都降 低。
碱性磷酸酶液
0.1 0.1 0.1 0.1 0.1 0.1 0.1 -
混匀后,370C水浴 15 min(准确计时)
酚试剂
1.0 1.0 1.0 1.0 1.0 1.0 1.0 1.0
碱性磷酸酶液
- - - - - - - 0.1
10% Na2CO3
3.0 3.0 3.0 3.0 3.0 3.0 3.0 3.0
研究前提
单底物、单产物反应
酶促反应速率一般在规定反应条 件下,用单位时间内底物的消耗 量或产物的生成量来表示。 反应速率取其初速率,即底物的消
耗量很小(≤5%)时的反应速率。
底物浓度远远大于酶浓度。
矩形双曲线
米-曼氏方程式
Km 即为米氏常数, Vmax为最大反应速度
当反应速度等于最大速度
一半时,即V = 1/2 Vmax, Km = [S]
在最适温度条件下, 反应速度最大。
pH 对反应速度的影响
在一定的pH 下, 酶具有最大的催化活性,通常称 此pH 为最适 pH。
激活剂对反应速度的影响
酶的激活剂
必需激活剂,
大多为金属离 子,Mg2+,K+ ,Mn2+,等
非必需激 活剂
抑制剂对反应速度的影响
不可逆性 抑制作用
抑制剂通常以共 价键与酶活性中 心或活性中心以 外的必需基团相 结合,使酶失活 。
影响酶促反应速度的因素
酶动力参数Km、Vmax和Kcat值地计算

酶动力参数Km、Vmax 和K c at 值的计算(以日立U3010 为例)强玮2014-3-31一、Km根据酶促反应方程,即米-曼式氏方程(Michaelis-Menten equation ):V =(VmaxS)/(Km + S)当v=0.5Vmax 时,Km=[S] ,可见Km 等于酶促反应的初速率为最大速率Vmax 一半时的底-6~10-2mol/L 之间,单位一般为mM 或者uM 。
Km 只与酶的性质有物浓度。
Km 值一般在10关,而与酶的浓度无关。
由于Km 值与酶的浓度无关,所以计算时无需对蛋白进行定量,理论上诱导破碎的菌体上清(粗酶液)即可用来测酶活计算。
具体计算方法以前常用双倒数法,但是这种算法现在稍微好一点的SCI 杂志都不认可了,相对来说现在更准确和更常用的计算方法是直接对不同浓度的底物测得的酶活数据用米氏方程(Michaelis-Menten equation V =(VmaxS)/(Km + S) )进行非线性回归(nonlinear regression),拟合的方程中就给出了计算出的Km 值。
非线性回归方法相对于双倒数法稍微麻烦的地方在于测量酶活时底物浓度的选择要尽量多,并且要覆盖米氏方程曲线的三个区域,以下图为例,酶反应速度遵循米氏方程的规律,先是一级反应,速度随底物浓度上升也直线上升,然后是混合级反应,趋于平缓,最后进入零级反应,反应速度不再随浓度上升而上升,而是趋于一个稳定的值。
根据这个规律,具体梯度测试时,底物浓度的选点很重要,要根据实时测得的斜率或活度值实时调整,尽量要覆盖这三个区域,这样回归拟合出来的米氏方程才是准确的,计算出的酶动力参数才是正确的。
初速度Hill Fit of 初速度0.30.2度速初0.10.00 20 40底物浓度(mM)用于非线性回归分析的软件很多,Origin 8.0 我比较喜欢,将酶活数据输入后,全选,“Analysis ”菜单下选择“Fitting ”,进一步选择“Nonlinear curve fit ”进入对话框,在函数归类框“Category”中选址“Growth/Sigmoidal ”类函数,接着在子函数框“Function”中选择“Hill ”函数。
alt的名词解释生物化学

alt的名词解释生物化学
ALT是肝功能生化检查中谷丙转氨酶的指标,也是肝功酶学的一个指标。
谷丙转氨酶是机体内参与氨基酸代谢的酶,在肝脏中含量最高,其正常值是0~40U/L,如果肝细胞受损,ALT就会释放到血中,反映肝细胞受损。
因此,ALT是肝细胞损伤的敏感指标。
谷丙转氨酶升高的原因有多种,例如饮酒、肝炎或者各种药物导致肝细胞损害。
如果患者的谷丙转氨酶出现异常,应及时到专业医院就诊、检查,明确病因后积极配合医生进行治疗,以免贻误病情。
以上内容仅供参考,建议咨询专业医生获取更具体准确的信息。
酶工程实验报告五(纤维素酶米氏常数—Km的测定)

v = Km + [S ]v :反应初速度(微摩尔浓度变化/min)V :最大反应速度(微摩尔浓度变化/min)[S]:底物浓度(mol/L)Km:米氏常数(mol/L)此方程表明,当已知Km及V时,酶反应速度与底物浓度之间的定量关系。
3.4 双倒数作图法测定Km值:1 Km 1 1v V [S] V1/v对1/[S]作图可得一条曲线,其斜率为Km/Vmax,截距为1/Vmax 。
若将直线延长与横轴相交,则该交点在数值上等于-1/Vmax。
本实验采用最适pH、最适温度下,测定不同浓度时酶活性。
再根据林贝法作图求出Km值。
3.5 纤维素酶水解纤维素产生的纤维二糖、葡萄糖等还原糖能将碱性条件下的3,5-二硝基水杨酸(DNS)还原,生成棕红色的氨基化合物,在540nm波长处有最大光吸收,在一定范围内还原糖的量与反应液的颜色强度呈比例关系,利用比色法测定其3mLDNS反应终止→沸水浴5min →定容至25ml→测定OD540吸光值G 作双倒数图求Km4.3 实验注意事项本实验是一个定量测定方法,为获得准确的实验结果,应尽量减少实验操作中带来的误差。
底物溶液时应用同一母液进行稀释,保证底物浓度的准确性。
严格控制准确的酶促反应时间。
反应温度准确,酶液准确稀释。
不同pH所用酶液用对应pH缓冲液稀释试管上编号:贴上用圆珠笔写上编号的胶布,以防止保温或沸水加热时脱落;移液管使用时量取精准,保证结果可靠准确。
精确记时:每一管加入酶液的时间要做记录,每管之间间隔的时间要合理;避免试管进水:煮沸和用流水冲洗时;由图-1/Km=-7,可得出:Km=0.143 mg/ml(g/L)/(2×104)=0.715×10-5mol/L。
6、分析与讨论:米氏常数(Km)是酶的一个特征性物理量,其大小与酶的性质有关。
Km 值随测定的底物种类、反应的温度、pH 及离子强度而改变。
因此,对某一酶促反应而言,在一定条件下都有特定的Km 值,可用来鉴别酶,例如对于不同来源或相同来源但在不同发育阶段,不同生理状况下催化相同反应的酶是否属于同一种酶[1]。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
一、酶的分离
1.材料的选取
在酶的分离过程中,首先应该考虑的一个问题是如何选择材料。同 一种酶在不同组织细胞中的含量可能相差千倍或上万倍,因此,如果不 是对某种酶在各组织中的含量进行比较分析或是某些特殊需要,一般选 择含量丰富的材料进行提纯。
2.细胞的破碎
常用的细胞破碎方法有:(1)绞碎、匀浆、高速组织捣碎机捣碎等; (2)对于细胞壁较厚的微生物材料,通常采用加石英砂研磨、超声波破碎、 反复冻融等方法;(3)对于细胞膜上的酶还可以采用有机溶剂处理、去垢 剂处理等。
第四节
酶法分析
酶法分析是指应用酶作为工具的分析方 法,可以对酶的底物、辅酶、激活剂或抑制 剂进行定量分析。常用的方法有动力学分析 法、终点测定法和酶标免疫测定法。其中终 点测定法的应用最为普遍。所谓终点测定法
就是借助某种酶的作用,使被测物质定量地
进行转变。在转化完成后,测定底物、产物 或辅酶等物质的变化量。它可分为单酶反应 定量法和偶联酶反应定量法。
二、酶浓度对反应速度的影响
在酶促反应系统中,当底物浓度大大超过酶的浓度,使酶被底物饱 和时,反应速度与酶的浓度成正比例关系。
三、温度对反应速度的影响
酶是生物催化剂,温度对酶促反应速度具有双重影响。温度升高时, 一方面可加快酶促反应速度,同时也增加酶的变性。综合这两种因素,酶 促反应速度最快时的环境温度称为酶促反应的最适温度。温血动物中酶的 最适温度多在35~40℃之间。温度高于最适温度时,反应速度则因酶变性 而降低。
第四章
酶学分析
酶(Enzyme)是生物体内具有催化功能的蛋白质,又叫生物催 化剂。生物体内的化学反应几乎都是在酶的催化下进行的,因此 酶学的研究,对于了解生命活动的规律,阐明生命现象的本质以 及指导有关的医学实践和工农业生产都有重要意义。
第一节
酶的分离纯化
酶的化学本质是蛋白质,故 适用于蛋白质的分离纯化方法一 般也适用于酶的分离纯化。
(1)盐析法:
盐析用的盐通常有硫酸铵和氯化钠等。由于硫酸铵溶解度 大,对大部分酶无毒副作用,且在浓度较高时对一些酶还有稳
定作用,易透析除去,故盐析绝大多数都用硫酸铵。
(2)聚乙二醇沉淀法: 聚乙二醇是最常用的分离核酸和蛋白质的沉淀剂。在部 分纯化的酶溶液中,缓慢加入聚乙二醇,使溶液达到混浊, 可以使某些酶以结晶的形式沉淀下来,使酶的提纯效果提高 3~5倍,且很少引起酶的失活。
4.电化学法
包括离子选择性电极测定法、氧电极测定法及电流测 定法等。
5.同位素法
用同位素标记底物,随着酶催化反应的进行,一部分标记的
底物将转化为产物。反应终止后,分离底物和产物,测底物的放 射性减少量或产物的放射性增加量。常用于底物标记的同位素有
3H、14C、32P、35S和131I。同位素法的特点是灵敏度极高,但操作
(ALT)活力单位按King氏法定义为:每100ml血清在37℃,pH-7.4
条件下与底物作用60 min,生成1μmo1丙酮酸为1个活力单位。
2.酶的比活力
比活力的大小,就是酶含量的大小,即酶蛋白所具有的酶活 力,一般用活力单位/毫克蛋白(U/mg蛋白)来表示,有时也用 每克酶制剂或每毫升酶制剂含有多少个活力单位来表示(单位/ 克或单位/毫升)。
烦琐、费时,有放射性,操作者须特别小心。Leabharlann 三、酶的活力单位(U)及比活力
酶活力的大小也即酶量的大小,用酶的活力单位来度量。
1.酶的活力单位
1976年国际酶学生化学会委员会规定:1个酶活力单位,是 指在特定条件下,1分钟内能将l微摩尔(μmol)底物转化为产物 所需的酶量,或是转化底物中1微摩尔的有关基团的酶量。 在临床上酶活力单位常用习惯表示法。如对血清谷丙转氨酶
二、测定方法
测定产物增加量或底物减少量的方法很多,常用的方法有 化学滴定、比色、比旋光度、气体测压、紫外吸收测定、电化学 法、荧光测定及同位素技术等。具体选择哪一种方法,要根据底 物或产物的物理化学性质而定。
1.化学法
这是一种取样法。终止酶反应通常采用加入酶的变性 剂的方法。如果反应底物或产物热稳定性好,也可用加热
一、底物浓度对反应速度的影响
在其他因素不变的情况下,底物浓度对反应速度影响的关系曲线呈 矩形双曲线。在底物浓度较低时,反应速度随底物浓度的增加而急剧上 升,两者成正比关系;随着底物浓度的进一步增高,应速度的增幅度不 断下降;如果继续加大底物浓度,反应速度将不再增加,此时酶的活性 中心已被底物饱和。
最适pH也不是酶的特性常数,它受底物浓度、缓冲液的种类与 浓度,以及酶的纯度等因素的影响。溶液pH高于或低于最适pH时, 酶的活性降低,远离最适pH时甚至会导致酶的变性失活。因此在测 定酶的活性时,应选用pH适宜的缓冲液以保持酶活性的相对恒定。
五、激活剂对反应速度的影响
使酶由无活性变为有活性或使酶活性增加的物质称为酶的激活剂 (activator),可分为必需激活剂和非必需激活剂。必需激活剂大多 为金属离子,如Mg2+、K+、Mn2+等,它们对酶促反应是不可缺少的, 否则将测不到酶的活性。非必需激活剂是指激活剂不存在时,酶仍有 一定的催化活性,但它的存在能提高酶的催化活性。
(2)非竞争性抑制作用:抑制剂与酶活性中心外的必需基团结合, 不影响酶与底物的结合,酶和底物的结合也不影响酶与抑制剂的结
合。这类抑制作用使酶的 υmax 降低,但不影响酶的Km。
3.反竞争性抑制作用
抑制剂仅与酶和底物形成的中间产物结合,使中问产物的量下降。 这类抑制作用同时降低反应的 υmax 和Km值。
3.电泳法 电泳法是利用酶蛋白所带电荷和分子量大小与其他杂蛋白 的不同而达到分离的目的。酶纯化中常用的电泳有聚丙烯酰胺 凝胶电泳、等电聚焦等,详见本书第二章。
4.结晶
结晶主要是利用酶蛋白在硫酸铵等溶液中的溶解度随
着温度升高而降低的特性将酶进行提纯。达到一定纯度的 酶才可以结晶,同时结晶本身也是一种提纯的方法。用于 结晶的酶制剂只要纯度达到50X以上就可以了。
使酶失活的方法终止酶反应,但时问较难控制。该法工作
量大,本身包含一定的误差,对速度快的反应常不易得到 十分准确的结果。
2.比色法
比色法是根据底物和产物在某~波长上有明显的特征 吸收差别建立起来的连续观测方法。
3.荧光法
利用酶反应的底物、产物或辅助物具有荧光,通过测荧光强度变化来 测酶活力。例如,某些脱氢酶的辅酶NADH(或NADPH)的中性溶液能发出强 烈的蓝白色荧光(460nm),而NAD+(或NADP+)则没有。用荧光法测酶活力 时,通常以单位时间内荧光强度的变化来表示。 荧光法的优点是灵敏度极高,比分光光度法还要高2~3个数量级,故 特别适用于酶量或底物量极低的快速酶分析。其缺点是:荧光读数与浓度 之间没有直接的比例关系,而且常因测定条件(如温度、散射、仪器等)的 不同而不同。
3.酶的转换数
指每秒钟每个酶分子转换底物的微摩尔数(μmol/s),它相 当于一旦底物一酶(ES)中间复合物形成后,酶将底物转换为产物 的效率。
第三节
酶促反应动力学
酶促反应动力学研究的是酶促反 应速度及其影响因素。这些因素包括 底物浓度、酶浓度、pH、温度、激活 剂、抑制剂等。酶促反应动力学的研 究具有重要的理论和实践意义。
第二节
酶的活力测定
一、酶活力与酶反应速度
1.酶活力 酶活力(Enzyme Activity)是指酶催化化学反应的能 力,也即酶催化某一化学反应的反应速度。检查酶的含量 及存在,不能直接用重量或体积来表示,通常用它催化某 一特定反应的速度来表示,即用酶的活力来表示。由于酶 活力受温度、pH等环境因素的影响很大,在进行酶活力测 定时,必须使这些因素保持恒定。
酶的最适温度不是酶的特征性常数,它与反应进行的时间有关。酶可 以在短时间内耐受较高的温度。相反,延长反应时间,最适温度便降低。
四、pH对反应速度的影响
酶分子中的许多极性基团在不同的pH条件下解离状态不同,所
带电荷的种类和数量也各不相同,酶活性中心的某些必需基团往往
仅在某一解离状态时才具有最大的催化作用。此时的环境pH称为酶 促反应的最适pH。动物体内多数酶的最适pH接近中性。
1.米-曼氏方程式
1913年,L.Michaelis和M.L.Menten提出了反应速度与底物浓度关系 的数学方程式,即著名的米一曼氏方程式,简称米氏方程式,即:
式中υmax为最大反应速度,[S]为底物浓度,Km为米氏 常数,υ是在不同[S]时的反应速度。
2.Km值和υmax值的测定
由于底物浓度对反应速度影响的关系曲线是矩形双曲线,因此很难
2.可逆性抑制作用
可逆性抑制作用的抑制剂通常以非共价键与酶或酶-底物复合物可
逆性结合,使酶活性降低或消失。采用透析或超滤的方法可将抑制剂除 去,使酶恢复活性。可逆性抑制作用的类型大体可分为以下三种:
(1)竞争性抑制作用:抑制剂与酶的底物结构相似,可与底物 竞争酶的活性中心,从而阻碍酶与底物结合形成中间产物。这类抑 制作用使酶的Km增大,但不改变酶的υmax 。
(3)等电点(pI)沉淀法:
酶蛋白在等电点处溶解度最小,易于沉淀,因此可以通过调节介质的pH
值对酶进行提纯。但由于蛋白质在pI附近一定范围的pH值溶液中都可发生沉 淀,而且相当多蛋白质的pI很接近,所以该法的提纯效果及回收率均不理想,
一般只用在酶的粗分离阶段。
(4)有机溶剂沉淀法: 常用的有机溶剂有乙醇、丙酮和甲醇等。该法是一种比较有效的分离 提纯方法。为了避免酶蛋白变性,须在低温(-5~-10℃)下进行分离提纯。
准确地测得Km值和υmax值。若将米氏方程式进行变换,将曲线作图改为 直线作图,便可容易地用图解法准确求得Km值和υmax值。最常用的是 双倒数作图法,又称为林-贝氏(Lineweaver-Burk)作图法,它将米氏方 程式等号两边取倒数,再以1/υ 对1/[S]作图,可得一直线,它在纵轴 上的截距为1/υmax,在横轴上的截距为-1/Km。