Gene-for-Gene_Theory解读
基因与基因组的结构与功能讲课文档

内含子存在意义:保证或增强基因的稳定表达;内含子中包含有其他 基因的编码序列(即基因内基因)。
GT-AG法则:真核生物基因内含子高度保守的一致序列:内含子5`端大 多数以GT开始,3`端大多数以AG结束,称为GT-AG法则。此法则 可用作真核基因中RNA剪接的识别信号。
基因与基因组的结构与功能
第一页,共75页。
3.1. 基因概念的变迁
3.2. 基因概念的多样性
3.3. 基因组
第二页,共75页。
3.1 基因概念的变迁
Theory of the gene (192实体
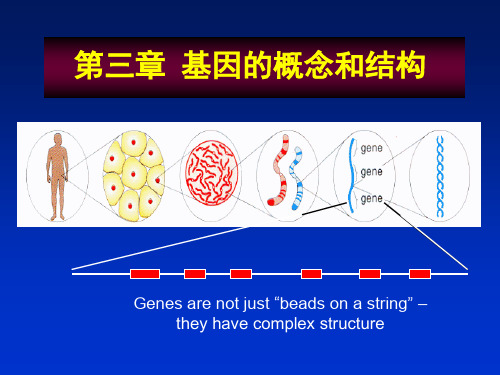
• 基因象链珠(bead)一样,孤立地呈 线状地排列在染色体上
OriC
0
4000K
大肠杆菌 1000K
C-Value: 4.6×106bp
3000K
2000K
TerC
大肠杆菌染色体DNA
第三十八页,共75页。
2. 结构基因大多组成操纵子
po z
y
at
promoter operator structural gene terminator
ß-galactosidase半乳糖苷酶 z ß-galactoside permease透酶 y
的结构基因的表达, 如:启动基因,操纵基因不转录 、不翻译
不是所有的基因都能为蛋白质编码
第七页,共75页。
✓ 基因命名无严格统一标准
1. 一般:三个小写英文斜体字母表示,lac、leu
2. 三个小写英文斜体字母后面加一个斜体大写字母表示
基因座, LacZ、 LacY、 LacA
《基因论》读后感

Reflections on "The Theory of the Gene"Reading "The Theory of the Gene" by Thomas Hunt Morgan was a profound experience that left me with a deeper understanding of the intricate and fascinating world of genetics. Morgan's comprehensive exploration of the principles and mechanisms of genetics provided a comprehensive and engaging overview of the subject.One of the most striking aspects of the book was Morgan's focus on the gene as the fundamental unit of heredity. His exploration of the particle theory of inheritance, chromosome-gene relationships, and the origin of mutations offered a clear and concise understanding of the complexities of genetic inheritance. The author's explanation of how genetic information is passed down from generation to generation through chromosomes was particularly enlightening.Morgan's writing style was engaging and accessible, making the complex concepts of genetics accessible to a wide range of readers. His use of examples and anecdotes throughout the book added a personal touch, making the subject matter more relatable and interesting.The book also provided a fascinating insight into the ethical and social implications of genetics. Morgan's discussion on the role of genes in gender determination and the potential for genetic engineering to impact society was particularly thought-provoking. It made me consider the ethical dilemmas that arise when we have the ability to manipulate genetic information and the potential consequences for future generations.Overall, "The Theory of the Gene" was an excellent introduction to the field of genetics. It provided a solid foundation of knowledge and understanding that will serve as a valuable resource for further exploration of this fascinating subject.I highly recommend this book to anyone interested in genetics or the science of heredity.阅读托马斯·亨特·摩尔根的《基因论》是一次深刻而富有启发性的体验,让我对遗传学这个复杂而迷人的领域有了更深入的理解。
遗传学第三章 基因的概念和结构
基因重叠方式
• Mis-reading for stop codon
( Q RNA virus 1973. A. Weiner )
400Nt
800Nt
AUG----------------------UGA-----------------------UAA
设有两个独立起源的隐性突变,具有类似的表现型。判断是属于同一个基因 突变,还是属于两个基因突变?即判断是否属于等位基因? ①建立双突变杂合二倍体; ②测定突变间有无互补作用。
• 顺反测验:顺式排列为对照(是两个突变座位位于同一条染色 体上),其表现型野生型。实质上是进行反式测验(反式排列是 两个突变座位位于不同的染色体上)。
① 反式排列为野生型:突变分属于两个基因位点; ② 反式排列为突变型:突变分属于同一基因位点。
Complementary assay
rII47 0
rII106 0
rII 47
rII106
rII106 0 rII51 0
rII106 rII51
Why?
plane E.coli K12
依据; One gene
2、假基因(pseudo gene)
• 假基因:同已知的基因相似,处于不同的位点,因缺失或突变而 不能转录或翻译,是没有功能的基因。
第五节、外显子和内含子
• 内含子(intron):DNA序列中不出现在成熟mRNA的片段; • 外显子(extron):DNA序列中出现在成熟mRNA中的片段。
Ovalbumin DNA X cDNA
5387 bp 11 genes 3 mRNA 9 peptides
遗传学Genetics

现代定义:
(1)在生物的群体、个体、细胞和基因等层次上研究生命信息 (基因)的结构、组成、功能、变异、传递(复制)和表达规 律与调控机制的一门科学--基因学。 (2)研究基因和基因组的结构与功能的学科。
Charles Darwin与进化论:
Charles Darwin (1809-1882) 1859年出版The Origin of Species 认为现存的物种是由古老的物种 渐变(modification )来的。 生存斗争与自然选择的进化理论。 用以解释他的进化原因的理论 支柱是natural selection
可观察到或检测到的生物的形态结构、生理或行为特征。
什么是生命的本质特征?繁殖与自身相似的同类(无 性生殖、有性生殖)
(自我繁殖=传递性状:种群的、群体的、家族的、个体的)
遗传与变异是生命自我繁殖过程中产生的两个现象。
What is Genetics/Genics
heredity, inheritance 遗传
Mendel以前的遗传学说
泛生论 (theory of pangenesis):
公元前5世纪希波克拉底Hippocrates提出。
希波克拉底学派认为子代之所以具有亲代的特性,是因 为在精液或胚胎里集中了来自身体各部分的微小代表元 素(active humors, element )。相信后天获得 (acquired)的性状是能遗传(inherit)的。
遗传使物种得以延续,使物种相对稳定; 变异使物种得以发展和进化,使世界丰富多彩,充满活力与希望。 变异是物种进化、新物种形成的基础和资本。
《gene的概念》幻灯片

Diplococcum pneumonice
DNA as genetic material 1948. retired, The Nobel committee has been criticized for not recognizing Avery’s achievement before his death ( 1877-1955 )
1953 Watson和 Crick提出了DNA构造 的右手双螺旋模型~double helix pattern。
大约30多年, 认为gene指导专一的
2.2.3 顺反子学说 1955年,S.Benzer研究了T4噬菌体rⅡ区的 精细构造,提出了顺反子学说(Theory of cistron):
一个顺反子就是一个基因,这个基因或 者编码蛋白质,或者编码RNA分子。
顺反子理论 Theory of cistron
(S.Benzer 1955)
对经典的基因概念的 修正与开展
1941. Beadle & Tatum
理论基础
Nuraspora crassa One gene ----one enzyme
1944 Avery O.T. (Canada)
Bachelor
物种 加强和完善对环境的适应
逐渐转变为新种
获得的性状是由环境影响 (非遗传物质的改变)
新性状一旦获得, 便能遗传给后代
2.1.3. 泛生论假说 (Hypothesis of the
Pangenesis)
C. Darwin 1868
决定单位性状的遗传
Panger
泛生子
1744 莫珀图斯(P.Maupertuis法国)提 出“泛生论〞(theory of pangenesis) 、 于1868年达尔文(Darwin) 承受“泛生论〞 ,并正式命名。
基因定位PPT课件

A BC
A1 B1 C1 A2 B2 C2
基因组学 Genomics
§4.1 基因的鉴定
§4.1.2 遗传分析
判断某一性状是否由主基因控制和受多少对基因控 制,首先需要对该性状作遗传分析。遗传分析的基本方 法是:
(1)选择具有相对性状的两个亲本杂交; (2)在F1中观察该性状属于完全显性,还是不完全 显性;
基因组学 Genomics
第四章 基因定位
基因组学 Genomics
§4 基因定位
§4.1 基因的鉴定 §4.2 主基因定位 §4.3 QTL定位 §4.4 分子标记辅助选择
基因组学 Genomics
§4 Mapping of genes
§4.1 基因的鉴定
基因定位( Mapping of genes )是指通过遗传作图的 方法,确定基因与遗传标记之间的关系。
(2)如果不知道要定位的基因与已定位的基因之间的 关系,但它们的表现型相同,则存在它们是同一个基因的 可能性,需要确定它们之间的关系。
基因组学 Genomics
§4 Mapping of genes
§4.2 主基因定位
基因定位通常指的是主基因的定位。而微效基因的 定位通常用专有名词QTL定位。
基因组学 Genomics
基因组学 Genomics
§4.2 主基因定位
§4.2.3 分子标记的分析
已定位标记的利用:
通过遗传图谱的构建,很多分子标记在染 色体上的位置已经确定,因此只要找到与基因 连锁的分子标记,就可以确定基因在染色体上 的位置。这类标记有RFLP标记和SSR标记等。
已定位标记的利用,通常是从现有的遗传 图谱中,按一定距离均匀选取分子标记。
基因组学 Genomics
基因的概念及发展
果蝇眼面大小遗传的剂量效应和位置效应
779
358
68
45
25
Illustration of the different sizes of compound eyes of the female Drosophila melanogaster as caused by the varying numbers of facets. The size of the eye is influenced by the position effect 果蝇的X染色体上16A区段
杂种内的镶嵌特征是亲本胚芽混合所致。
他认为在生活周期的任何阶段细胞都可放出胚芽, 胚芽随血液循环到达生殖细胞,并传递给后代。
4.种质论(Theory of germplasm, Weismann,1885)
多细胞生物的细胞可分为“体质”和“种质” 后天获得性只能改变体质(Somatoplasm,体细胞),
遗传性状由遗传因子决定 遗传因子成对存在 生殖细胞中具有成对因子中的一个 每对因子分别来自雌雄亲代的生殖细胞 形成生殖细胞时,成对因子相互分离 生殖细胞的结合是随机的 遗传因子有显隐性之分
孟德尔定律:
分离定律(Law
of segregation) 自由组合定律(Law of independent assortment) 否定了融合遗传、泛生论及获得性遗传理论 为颗粒遗传理论(particulate theary of inheritance )奠定 了基础 1900年,孟德尔定律被重新发现
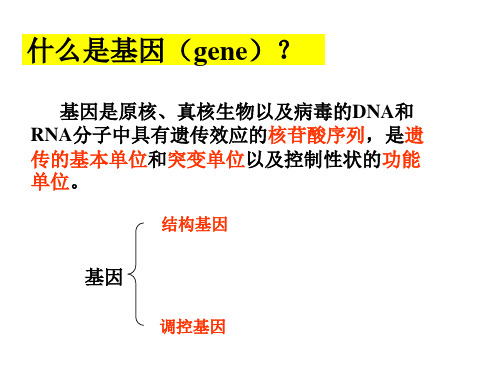
什么是基因(gene)?
基因是原核、真核生物以及病毒的DNA和 RNA分子中具有遗传效应的核苷酸序列,是遗 传的基本单位和突变单位以及控制性状的功能 单位。
gene-for-gene

2.1 细菌的无毒基因
无毒基因存在与否决定着病原菌能否侵入 含相应抗病基因的寄主植物或侵入后细菌能 否大量增殖。典型的诱导抗病性反应是过敏 反应(HR),它引起一种快速局部的细胞死 亡,伴随着受侵染植物组织中的细菌生长受 抑制。 迄今为止已有超过40种细菌的无毒基因被 克隆、测序,主要分为两种:avrBs3基因家 族和avrRxv/yopJ家族
其基本原理是利用转座子或t-DNA插入到抗病 基因的内部或邻近位点,获得抗病功能丧失的突变 体,然后利用插入的片段(转座子或t-DNA)作探针, 通过前提条件是被 操作的植物有现成的遗传转化操作系统或有成熟的 根癌农杆菌转化系统以及有效的大规模突变体筛选 的方法和手段。
3.4 亮氨酸拉链(Leucine zipper,LZ )
LZ存在于一些寡聚蛋白中,许多DNA结合蛋白 就含有LZ。LZ在真核生物转录因子的同源及异源二 聚体形成中起着重要作用,相似的卷曲螺旋结构域 (Coiled-coil domains)促进了蛋白之间的相互作用 ,并导致许多其他功能的产生,但它们在抗病蛋白 功能发挥中所起的作用目前还知之甚少。推测它可 能在抗病反应信号的传导过程中与病原物产生的配 体结构特异性结合相关。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
R Plant cell becomes diseased
Avr allele Virulent pathogen
No R allele; plant cell becomes diseased
Virulent pathogen
No R allele; plant cell becomes diseased
,如玉米Hm1 基因和大麦mlo 基因,其抗性机制与病原菌的亲
和因子有关,而不需要病原菌无毒因子的激活。
2.1符合基因对基因关系的互作模式
2.1.1 受体-配体模式 (Receptor ligand model)
受体-配体模型是从基因对基因假 说发展而来的。该模型认为:病原体 的 avr 基因直接或间接地编码一种配 体(激发子),它与抗病基因(R)编码的 产物(受体)相互作用,从而触发受侵 染部位细胞内的信号传递过程,激活 其他防卫基因的表达,产生超敏反应 (Hypersensitive Response, HR) , 在病原体侵染部位出现枯死斑点症状, 使植物获得抗性。
பைடு நூலகம்
(Plant Responses to Internal and External Signals Chapter 39)
该学说认为,植物对某种病原物 的特异抗性取决于它是否具有相应 抗性基因,而同时病原物的专一致病
性取决于病原物是否具有无毒基因,
也就是说寄主分别含有感病基因(r) 和抗病基因(R),病原物分别含有毒性 基因(Vir)和无毒基因(Avr),只有当 具有相应抗病基因的植物(R)与具有 无毒基因(Avr)的病原物相遇时,才 会激发植物的抗病反应,其他情况下 二者表现亲和,即寄主感病。
2.1.3 已提出的其他相关模式
(a)受体-配体模式
(b)共受体模式:无毒蛋白(A)先
与 共受体(C)上的高亲和位点结合, R蛋白再与 C之间发生相互作用启动防 御反应。
(c)警戒模式
(d)蛋白水解酶依赖型防御启动:介
于一些细菌、真菌和病毒的特点,预测 这些基因可能会编码蛋白质水解酶。该 论点认为 Avr蛋白会对寄主中的某些蛋 白的水解作用的处理后寄主内的R蛋白 识别并与部分水解状态的蛋白结合,引 起信号传导,引发植物的防御反应。
3.1 三型分泌系统(TTSS / T3SS )
A transmission electron-microscope image of isolated T3SS needle complexes from Salmonella typhimurium
2.2 非R/Avr基因互作的植物抗病模式
寄主植物中存在促进亲和反应的基因,即感病基因,其编码产物是一种 “亲和性因子”。通过对这类基因的修饰甚至诱发隐性突变,即可赋予植 物对某一种甚至多种病原菌的广谱抗性。这类基因有大麦抗白粉菌( Erysiphe graminis f sp, Hordei)基因mlo,豌豆抗白粉菌(E.pisi) 基因er-1,番茄抗白粉病(Oidium lycopersici) 基因ol-2 和拟南芥抗 白粉病基因 PMR 等。
3 不同Avr与R蛋白互作的定位
不同寄主与病原物Avr和R蛋白的互作定位。对一些细菌Avr和R蛋白的 匹配性分析,揭示了普遍的空间相互依赖性。如上图,植物的R蛋白用深绿 色标出,其他与植物抗性相关蛋白用浅绿色标出。细菌、真菌和预测的线虫 Avr蛋白分别用红色、紫色和粉红色标出。微生物侵染结构用蓝色标出。
1 植物基因对基因假说
植物与病原微生物的侵袭,两者 在自然生态系统中长期并存,相互影 响乃至协同进化,使得植物的抗病性 与病原物的致病性之间形成一种动 态平衡。
1971年,Flor 根据亚麻对锈菌小 种特异抗性的研究提出了基因对基 因假说(Gene-for-gene theory) 。
Harold H. Flor
(/course/pp315/gene/)
2.1.2 警戒模式 (Guard model )
模式认为植物寄主中存在着病原菌 效应蛋白(Avr-effector)致毒性靶标 (Virulence target),效应蛋白与 之互作来调节植物活性,以利于病原 菌生长和繁殖;NB-LRR 类 R 蛋白起 警戒作用,一旦监测到靶蛋白被Avr 改变,即可激活 R 蛋白与靶蛋白互作 ,以避免病原菌效应蛋白对靶蛋白的 操纵,或引发一系列信号传导,启动 植物的防御反应。
2 R与Avr基因互作的模式
植物与病原菌的分子交流决定了它们互作的最终结果。病原菌 效应蛋白被送入寄主植物细胞后,能否被侵染的植物组织识别, 决定了植物是感病还是抗病。由专化 R 基因介导的抗性符合“基 因对基因”的互作模式。然而,从已克隆的 R 基因及其产物结构 和功能看,有些 R 基因的抗病机制并不符合“基因对基因”模式
Signal molecule (ligand) from Avr gene product
Avr allele Avirulent pathogen
Receptor coded by R allele R
Plant cell is resistant
No Avr allele; virulent pathogen
Gene-for-Gene Theory
拓庆荣 21014032
目录
• 1 植物基因对基因假说 • 2 抗病基因的作用机理 • 3 不同Avr与R蛋白互作的定位 • 4 病原物无毒基因及无毒蛋白 • 5 植物抗病基因 • 6 R基因编码R蛋白的结构特点 • 7 植物抗病基因的克隆 • 8 植物抗病基因的应用