免疫补体激活的三条途径的比较
医学免疫学三条激活途径记忆方法

医学免疫学三条激活途径记忆方法医学免疫学是研究机体免疫系统的科学,其中激活途径是免疫系统中非常重要的一环。
免疫系统具有三条主要的激活途径,包括经典途径、替代途径和MBL途径。
本文将围绕这三条激活途径展开,介绍它们的特点和作用。
一、经典途径经典途径是免疫系统中最早被发现的激活途径,也是最重要的一条途径。
它的激活是通过抗原-抗体复合物与C1q结合来启动的。
C1q是经典途径的起始蛋白,它能够识别并结合到抗原-抗体复合物上,从而激活C1r和C1s两个酶。
接下来,C1s酶会切割C4和C2蛋白,生成C4b和C2a,在C4b和C2a的作用下,形成C3转化酶C4b2a。
C3转化酶能够切割C3蛋白,生成C3a和C3b。
C3b 能够与C4b2a结合,形成C5转化酶C4b2a3b,进而切割C5蛋白,引发免疫应答。
二、替代途径替代途径是免疫系统中另一条重要的激活途径,它与经典途径不同,不依赖于抗原-抗体复合物。
替代途径的激活是通过C3蛋白的水解来实现的。
在正常情况下,C3蛋白会随机水解,生成C3b。
C3b 能够与主要组织相容性复合物(MHC)上的补体受体结合,形成C3转化酶C3bBb。
C3转化酶能够切割更多的C3蛋白,进一步放大免疫应答。
替代途径的激活速度较快,能够迅速启动免疫反应。
三、MBL途径MBL(mannose-binding lectin)途径是一条与免疫系统中的糖蛋白Mannose结合的途径。
MBL是一种能够识别并结合到病原体表面Mannose的蛋白,它能够激活免疫系统。
MBL途径与经典途径有相似的激活方式,也是通过Mannose-结合的MBL与Mannose 结合的抗原结合来启动的。
MBL与Mannose结合的抗原结合后,激活MASP(MBL-associated serine protease)酶,进而切割C4和C2蛋白,生成C4b和C2a,最终形成C3转化酶。
三条激活途径在免疫系统中起着不可或缺的作用。
试述补体活化的三条途径

1. 试述补体活化的三条途径。
答:1、经典激活途径:(1)激活物:主要是由IgG或IgM类抗体与相应抗原结合形成的免疫复合物。
(2)参与成分:C1、C2、C3、C4、C5、C6、C7、C8、C9。
(3)激活过程:分为识别阶段、活化阶段、膜攻击阶段。
(4)激活顺序:依次为C1、C4、C2、C3、C5--C9。
(5)转化酶:C3转化酶:C4b2b;C5转化酶:C4b2b3b。
(6)生物学作用:可在特异性免疫的效应阶段发挥作用。
C5转化酶裂解C5后形成膜攻击复合物,最终溶解靶细胞;补体裂解形成的小片段C4a、C2a、C3a 在血清等体液中可发挥多种生物学效应。
2、旁路激活途径:(1)激活物:主要是病原体胞壁成分,如脂多糖、肽聚糖、磷壁酸等。
(2)参与成分:除C3、C5、C6、C7、C8、C9外,还有B因子、D因子、P因子等。
(3)激活过程:首先激活C3,然后完成C5--C9的活化过程。
(4)激活顺序:依次为C1、C4、C2、C3、C5--C9。
(5)转化酶:C3转化酶:C3bBb;C5转化酶:C3bBb3b或C3bnBb(6)生物学作用:参与非特异性免疫,在感染早期发挥重要作用。
其生物学效应与经典途径相似。
3、MBL激活途径:(1)激活物:表面具有甘露糖、葡萄糖的病原微生物。
(2)参与成分:MBL、C反应蛋白、C2、C3、C4、C5、C6、C7、C8、C9。
(3)激活过程:A、MBL:MBL激活起始于炎症急性蛋白与病原体的结合。
MBL 与病原体表面的甘露糖等糖类配体结合后,激活与之相连的MBL相关的丝氨酸蛋白酶(MASP)。
MASP2与C1s活性相似,其激活补体的过程与经典途径相似;MASP1具有C3转化酶活性,其激活补体过程与旁路途径相似。
B、C反应蛋白:C反应蛋白与C1q结合使之活化,激活过程与经典途径相似。
(5)转化酶:C3转化酶、C5转化酶与经典途径相同。
(6)生物学作用:参与非特异性免疫,在感染早期发挥重要作用。
补体系统的三条激活途径的异同点

补体系统的三条激活途径的异同点
补体系统啊,那可是免疫系统里超级厉害的一部分呢!它有三条激活途径,经典途径、旁路途径和凝集素途径,这三条途径既有相同点,又有各自独特的地方,就像三胞胎一样,有相似也有不同。
先来说说相同点吧。
它们不都是为了保护我们的身体嘛,都是免疫系统的重要防线呀!不管是经典途径、旁路途径还是凝集素途径,最终不都是为了对抗病原体,让我们保持健康嘛。
它们就像一群勇敢的战士,在身体里时刻准备着战斗呢!
但它们的不同点也很明显呀!经典途径就像是正规军,得有特定的“敌人”出现才会被激活,是不是很严谨呢?旁路途径呢,就像一群机灵的游击队员,随时都能快速反应,管他什么情况,先冲上去再说!而凝集素途径呢,就像是有特殊情报的特工,能识别一些特别的“信号”然后行动起来。
经典途径的激活一般需要抗体的参与呀,这就好像要有上级的命令才能行动。
而旁路途径可不需要这些,它自己就能找到“战机”,随时准备出击,多牛啊!凝集素途径则是凭借对某些糖类的识别来启动,就像有一双特别的眼睛能发现别人发现不了的东西。
再想想看,要是没有这三条途径的相互配合,我们的身体得面临多大的危险呀!它们就像是一个团队,各自发挥着自己的优势,共同守护着我们的健康。
这难道不神奇吗?这三条途径就像是三道坚固的防线,让病原体难以突破。
补体系统的三条激活途径真的是太重要啦!它们的存在让我们的免疫系统更加完善,让我们能更好地抵御疾病的侵袭。
我们真应该庆幸身体有这么厉害的保护机制呀!它们就是我们健康的守护者,没有它们可不行呢!。
补体活化的三种途径的异同点

补体活化的三种途径的异同点。
补体系统是一组广泛存在于血清、组织液和细胞膜表面的经激活后具有酶活性的蛋白质,参与机体抗微生物的防御反应和免疫调节,也可介导某些免疫病理性组织损伤和炎症反应。
在生理情况下,多数血清补体组分以无活性的酶原形式存在;但在特定条件下,如在某些活化物的作用和特定的反应表面,补体各成分才依次被激活,发挥生物学作用。
根据起始物和激活顺序的不同,补体的活化分为经典途径、甘露聚糖结合凝集素途径和旁路途径。
经典途径凝集素途径旁路途径不同点参与组分C1q、 C1r 、 C1s、MBL 、 MASP-1 、 B 因子、 D因子、 PC2、 C4MASP-2起始物抗原抗体复合物病原微生物表面甘细菌、 G-内毒素、酵露糖残基母多糖、葡聚糖活化顺序依次活化 C1q、依次活化 MASP、C2、活化 C3b、 B 因子、 DC1r 、C1s、C2、C4、C4、C3,形成 C3 与因子、备解素P、 C3,C3,形成 C3 与 C5C5 转化酶形成旁路途径C3转化转化酶酶,同时存在C3b 的级联放大机制是否依赖抗体是否否参与的免疫反特异性的体液免天然免疫天然免疫应疫相同点参与组分共同的固有成分C3,共同的末端反应成分C5、 C6、 C7、C8、 C9共同的末端效形成 C5 转化酶把C5裂解成 C5a和 C5b,若补体激活发生在脂质应双层上,则 C5b与 C6、 C7、 C8、 C9 形成膜攻击复合物(MAC);若发生在血清中,则与S 蛋白结合形成无溶细胞活性的物质。
作用形成 MAC,介导靶细胞溶解,发挥抗感染免疫的作用;活化过程中,产生多种具有炎性介质活性的片段,参与炎症反应;维持机体内环境稳定。
总之,补体的三种活化途径有各自不同的启动机制,但可以产生共同的末端效应,同时,这三种活化途径间存在密切的联系,如经典途径产生的C3b 可以触发旁路途径,旁路途径的C3 转化酶对经典途径的补体激活也有放大效应,这使得补体在免疫效应的不同阶段参与机体的抗感染免疫反应及其他多种生物学功能。
3条补体激活途径

3条补体激活途径三条补体激活途径补体是一种重要的免疫系统组分,它能够识别并攻击入侵人体的病原体。
补体激活是指补体分子在识别到病原体后,通过一系列的反应,最终导致病原体被摧毁。
补体激活途径有三种,分别是经典途径、替代途径和凝集素依赖性途径。
经典途径经典途径是补体激活途径中最早被发现的一种。
它的激活需要依赖于抗原-抗体复合物的存在。
当抗原-抗体复合物与补体分子结合时,会激活C1分子,从而引发一系列的反应。
这些反应包括C1激活后,C4和C2分子结合形成C4b2a复合物,也称为C3裂解酶。
C3裂解酶能够切割C3分子,形成C3a和C3b。
C3b分子能够与其他补体分子结合,形成C5裂解酶,最终导致病原体的溶解。
替代途径替代途径是补体激活途径中最早被发现的一种。
它的激活不需要抗原-抗体复合物的存在,而是依赖于补体分子与病原体表面的结合。
当补体分子与病原体表面结合时,会激活C3分子,形成C3裂解酶。
C3裂解酶能够切割C3分子,形成C3a和C3b。
C3b分子能够与其他补体分子结合,形成C5裂解酶,最终导致病原体的溶解。
凝集素依赖性途径凝集素依赖性途径是补体激活途径中最后被发现的一种。
它的激活需要依赖于凝集素分子的存在。
凝集素是一种能够识别病原体表面糖类的分子。
当凝集素分子与病原体表面糖类结合时,会激活Mannose 结合凝集素(MBL)分子,从而引发一系列的反应。
这些反应包括MBL 激活后,C4和C2分子结合形成C4b2a复合物,也称为C3裂解酶。
C3裂解酶能够切割C3分子,形成C3a和C3b。
C3b分子能够与其他补体分子结合,形成C5裂解酶,最终导致病原体的溶解。
总结补体激活途径是免疫系统中非常重要的一部分。
经典途径、替代途径和凝集素依赖性途径是三种不同的补体激活途径。
它们的激活方式不同,但最终都能够导致病原体的溶解。
对于研究免疫系统和疾病治疗都有重要的意义。
三条补体激活途径

三条补体激活途径补体是一种重要的免疫系统组成部分,它在机体的防御和清除病原体方面发挥着重要的作用。
补体系统可以通过多条途径激活,包括经典途径、替代途径和凝集素途径。
本文将依次介绍这三条补体激活途径的特点和作用。
一、经典途径经典途径是补体激活的主要途径之一,它最早被发现并得到了广泛研究。
经典途径的激活需要抗体的参与,当机体感染病原体时,免疫系统会产生特异性抗体来识别和结合病原体。
当抗体与病原体结合后,经典途径的第一组分C1q会结合到抗体的Fc部分上,从而激活经典途径。
激活后的经典途径会依次激活C1r、C1s、C4、C2等组分,形成C3裂解酶,最终导致病原体的溶解和清除。
二、替代途径替代途径是补体激活的另一条重要途径,它与经典途径不同,不需要抗体的参与。
替代途径的激活主要依赖于宿主细菌表面的一些特殊结构和病原体的特异抗原。
当宿主细菌或病原体感染机体时,它们的表面会暴露出一些特殊结构,例如脂多糖、甘露糖等。
这些结构可以直接激活替代途径的第一组分C3,进而激活后续的组分,形成C3裂解酶,最终引发病原体的溶解和清除。
三、凝集素途径凝集素途径是补体激活的第三条途径,它与经典途径和替代途径有一些相似之处,但也有一些独特的特点。
凝集素途径的激活依赖于一类特殊的蛋白质,称为凝集素。
凝集素在机体中广泛存在,可以与病原体表面的一些特定糖基结构结合,从而激活凝集素途径。
激活后的凝集素途径会依次激活MASP-1、MASP-2等组分,形成C3裂解酶,最终引发病原体的溶解和清除。
补体系统可以通过经典途径、替代途径和凝集素途径来进行激活。
经典途径依赖于抗体的参与,替代途径依赖于病原体的特异结构,而凝集素途径则依赖于凝集素与病原体表面糖基的结合。
这三条途径相互补充,共同参与机体的免疫防御和病原体的清除。
研究补体激活的途径对于我们深入了解免疫系统的调控机制,发展新的免疫治疗策略具有重要的意义。
补体激活的原理

补体激活的原理
补体激活是一个复杂的生物学过程,涉及多个补体成分和反应。
它主要通过三条途径进行:经典途径、替代途径和MBL途径。
1.经典途径:由特异性抗体的反应启动,激活后,某些补体蛋白酶例如C1酶、C4酶、C3酶等会被激活。
这些酶的活性可以迅速地催化级联反应,从而释放高能化合物如C3b、C4b等在体液中。
2.替代途径:这一途径直接由微生物或其他非特异性免疫因素激活,不依赖于抗体。
与经典途径相似,替代途径也会产生C3b等补体成分,引发级联反应。
3.MBL途径:主要由与甘露糖结合蛋白(MBL)的结合活化。
在补体激活过程中,级联反应会产生一些多肽片段,如C3a和C5a,它们被称为过敏毒素。
这些多肽片段会招募并激活免疫细胞,放大免疫响应。
同时,C3a和C5a通过与相应的受体(如C3aR和C5aR)结合,进一步触发下游信号通路,调节免疫反应。
异常的补体激活与多种疾病的发生有密切联系,例如免疫缺陷、自免疫紊乱等。
因此,对补体激活原理的理解有助于深入探究这些疾病的发病机制,并为开发新的治疗方法提供理论基础。
以上内容仅供参考,建议查阅专业生物学书籍或文献以获取更准确的信息。
补体激活的三条途径特点的比较
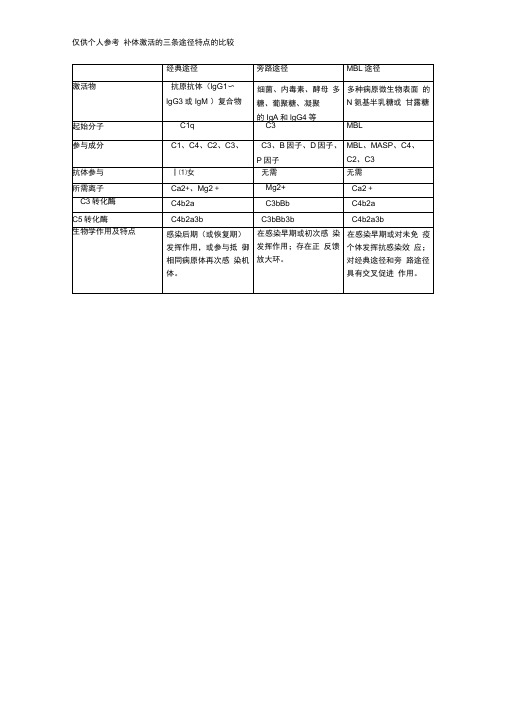
C2、3
抗体参与
丨⑴女
无需
无需
所需离子
Ca2+、Mg2+
Mg2+
Ca2+
C3转化酶
C4b2a
C3bBb
C4b2a
C5转化酶
C4b2a3b
C3bBb3b
C4b2a3b
生物学作用及特点
感染后期(或恢复期) 发挥作用,或参与抵 御相同病原体再次感 染机体。
在感染早期或初次感 染发挥作用;存在正 反馈放大环。
在感染早期或对未免 疫个体发挥抗感染效 应;对经典途径和旁 路途径具有交叉促进 作用。
仅供个人参考
仅供个人用于学习、研究;不得用于商业用途
For personal use only in study and research; not for commercial use.
Nur fuden pers?nlichen fur Studien, Forschung, zu kommerziellen Zwecken verwendet werden.
仅供个人参考补体激活的三条途径特点的比较经典途径旁路途径mbl途径激活物抗原抗体lgg1?lgg3或igm复合物细菌内毒素酵母多糖葡聚糖凝聚的iga和lgg4n氨基半乳糖或甘露糖起始分子c1qc3mbl参与成分c1c4c2c3c3b因子d因子p因子mblmaspc4c2c3抗体参与无需无需所需离子ca2mg2mg2ca2c3转化酶c4b2ac3bbbc4b2ac5转化酶c4b2a3bc3bbb3bc4b2a3b生物学作用及特点感染后期或恢复期挥作用或参与抵御相同病原体再次感染机体
仅供个人参考 补体激活的三条途径特点的比较
经典途径
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
调节分子 体液中可溶性调节因子 C1抑制物 180 105 使 C1s 失去活性,不能裂解C4、C2 血清浓度 μg/ml 分子量(KDa) 生物学活性
C4结合蛋白(C4bp)
I因子(C3b灭活因子) H因子(C3b灭活促进因子) S蛋白(MAC抑制物) SP40/40 膜结合蛋白 CR1(C3b受体1, CD35) MCP(膜辅助蛋白,CD46)
DAF(促衰变因子)
HRF(同源限制因子/C8 bp) MIRL 膜反应性溶解抑制物 CD59
75
分布于多数血细胞表面加速
的降解
分布在血细胞膜表面抑制旁观细胞溶解 分布于红细胞、淋巴细胞、单核细胞、血小板, 防止C7、C8与 结合,防止MAC形成
分子量(KDa)
补体的生物学活性
生物学活性 补体分子
250
50 400 500 50
590
90 150 80 0.08 200 45~70
使C4b裂解,加速 C4B2B 衰变
灭活C3b、C4b 置换 C3bBb 中的Bb,使其失去活性 阻止MAC插入到靶细胞 抑制MAC的形成,防止MAC溶细胞作用 分布于多数血细胞上,加速C3转化酶的降解 分布于血细胞(除红细胞)和上皮细胞,辅助I 因子介导C3b、C4b降解
增强血管通透性
与肥大细胞或嗜碱性粒细胞结合后,释 放组胺等血管活性介质,使血管扩张 吸引中性粒细胞及单核细胞 与IC结合后,粘附于红细胞、血小板等 细胞上,易被吞噬与清除 与免疫细胞结合后,调节细胞的功能 增加抗体对病毒的中和作用促进某些包 膜病毒的溶解
C1、C2、C3、C4
C1~C9
C3b、C4b
作用机理
细胞(病毒感染细胞,肿瘤细胞和红细 胞)或细菌溶解 与理作用 炎症介质作用
激肽活性
过敏毒素 趋化作用 免疫粘附及清除IC 免疫调节作用 中和溶解病毒
C2a
C5a 、C3a、C4a C3a、C5a 、 C3b、C4b C3b、C1q