从Y染色体看日本人、韩国人、越南的来源
从Y染色体单倍型论证南北汉族的单一起源m

从Y染色体单倍型论证南北汉族的单一起源第一部分:以Y染色体单倍体类型研究和人类非洲起源论我先讲讲Y染色体单倍体类型的方式分析的原理和方法。
人有46条染色体(Chromosome),其中44条为常染色体,X、Y两条为性染色体。
XY组合的为男性,XX组合的为女性,所以,Y染色体只能父子相传,我们研究Y染色体,可以比较清晰的发现人群的迁徙和发展。
为了研究Y染色体的单倍型(Y-Chromosome Haplotype),我们绘制了人类Y染色体谱系树,我们按着Y染色体单倍型的不同把全人类(包括非洲)分为18个类型(用从A到R十八个字母代表),出现频率高,数量多的类型单独列出(如O、N),把小概率出现的类型列入上级母类(如F、P),这也就是类型上有逻辑上重合的缘故,这样做是为了使分类清晰,分析方便!人类Y染色体谱系树(图)此主题相关图片如下:非洲人是人类的大本营,今天,所有人类的Y染色体单倍体类型都能在非洲找到(指M168\YAP\M130三种基本类型)。
而非洲以外的居民,他们都是一支出非洲居民的后裔,他们的Y染色体上都带有M168的突变点。
(图)人类刚出非洲,都带有M168的变异点,此后,M168又发展出三个支系,C—M130、D—YAP、F—M89,它们下又有不同的子类型。
世界各地人群谱系关系树(图)此主题相关图片如下:F—M89是现代人中占压倒性的类型,大概全世界80%以上的人都有这个变异点。
F—M89有很多子类型,其中,K—M9是最重要的一个支系,亚欧大陆上,除了西亚地区和西伯利亚,世界岛上绝大部分民族都由K—M9的子孙构成,今天中国人(汉族)中,96%的人都是K—M9类型。
K—M9下面也有很多子类型,最重要的有两个,一个是ON—M214,另一个是P—M45。
前者(ON—M214)主要分布在东亚、东南亚、北亚东部、部分太平洋岛屿(以上都是O—M175子类型)和乌拉尔山两侧、北欧北部、东欧北部和北极圈内的爱斯基摩人、中国云南地区、华南和华北部分地区(以下都是N—M231子类型)。
关于黄种人来源和东亚人种迁徙再探讨

关于黄种人来源和东亚人种迁徙再探讨(2011-12-29 16:03:21)转载▼关于黄色人种或蒙古人种,其人种体质特征与产生来源等观点有很多。
一般认为黄种人主要体质特征有:- 肤色在世界所有人种中居中,呈黄色或黄褐色- 头发黑且直- 面部平坦,圆润,较宽几年前复旦李辉先生在他那篇有名的《走向远东的两大现代人种》文章中根据当时已经有的粗略数据给出了一些“惊人”结论,归纳起来我以为他主要认为黄种人在缅甸产生论,黄种人(NO)屠杀驱赶棕色人种论(CD),或叫新老亚洲论。
这几年有了更深入的分子人类学数据,回头再看,我认为李辉先生这两大观点都是十分错误的。
而新老亚洲论更是给很多人怕被测出是“老亚洲”或者已经被贴上“老亚洲”标签的人背上了心理包袱。
没有时间做很多精细研究和给出精细数据分布,但大致理出本人观点。
1 黄种人产生地应该是在中亚下图是一张人类肤色深浅分布图可以看出,纬度越高,皮肤越白,越接近赤道,皮肤越黑。
说明肤色与纬度关系很大。
可能其他环境要素也会产生一定影响,但是显然纬度会是最主要影响因素。
从肤色分布看,最初的黄种人不太可能在东南亚或者中国南方产生。
对比一下现在中国南方与同纬度的其他地方人种皮肤颜色,当肤色更深的澳美人种从接近赤道地方进入到缅甸或华南北纬30度线附近的时候,应该不足以产生黄种人突变,说明该地不太可能产生肤色更浅的黄色人种。
而现在中国南方人种较浅的皮肤颜色主要是很晚期的黄色人种从北向南扩散与本地混合为主而形成的。
李辉说在缅甸产生黄种人,这没有任何逻辑或其他证据证明,这是根据当时猜想的NO迁徙路径纯粹想当然地凭空而说。
相反肤色分布图的逻辑是可以看做对这种看法的否定。
实际上更高纬度的中亚与所处地理位置则应是诞生黄种人的最佳理想场所。
最初的黄种人最有可能就是在中亚产生的。
2 东亚父系人种的南北两线迁移路径(注:以下单倍群名称以Y-开头表示是父系单倍群名称,以mt-开头表示是母系单倍群名称)根据中科院昆明所文章,认为东亚中国人种93%的父系(主要指Y-NO系,包含Y-CD系), 来自于南线的迁移。
Investigative Genetics:从Y染色体解析东亚人群历史

Investigative Genetics:从Y染色体解析东亚人群历史作者:王传超李辉来源:科学网2013-6-17 17:14:19分享到:1关键词:东亚人群性染色体东亚是亚洲的一部分,面积广阔、风光旖旎。
东亚有着占世界22%的人口,主要分为四种体质类型:新石器时代东亚人,即蒙古人种;旧石器时代大洋洲人,即澳大利亚人种;旧石器时代东南亚人,即尼格利陀人种;还有欧洲人即高加索人种。
东亚有阿尔泰、南亚、南岛、侗傣、苗瑶、汉藏和印欧等七个语系的200多种语言。
这使得东亚成为世界上研究人类进化、遗传多样性和基因与文化相互作用的最重要区域之一。
在过去数年中,分子人类学的研究者们通过使用常染色和X染色体、父系Y染色体、母系线粒体等等各类遗传标记体系来解析东亚人群的遗传多样性。
常染色体和X染色体遗传自父母双方,会被重组所打乱,而Y染色体上主干的非重组区呈严格父系遗传,并且Y染色体的“有效群体大小”理论上至多为常染色体的四分之一,X染色体的三分之一,对漂变非常敏感,容易形成群体特异性多态标记,从而包含更多的关于群体历史的信息。
Y染色体的这些特点使其成为研究人类进化和迁徙最强有力的工具之一。
Y染色体进入人们的视野,开始于其在追溯现代人起源上的应用。
自上世纪90年代以来,人类学界争论最激烈的话题,是东亚地区现代人的起源问题。
由于东亚出土了大量的古人类化石,一些人类学家认为东亚地区的人类是本土连续进化的,支持全球现代人的多地区起源。
然而,1999年宿兵等人采用Y染色体非重组区的19个SNP来研究东亚人群,得出东亚地区现代人起源于非洲,并由南方进入东亚,而后向北方迁。
随后,2000年柯越海等人的对东亚地区12127份男性随机样本的Y染色体进行SNP分型研究。
Y染色体突变M168被认为约是7.4万年前现代人走出非洲时所产生的突变,其原始型仅出现在东非人群中,除非洲以外的人群都是突变型。
柯越海等人的研究虽然没有直接检测M168这个突变,但他们检测了M89、M130和YAP这三个M168下游的突变。
人类源流——人类Y染色体4
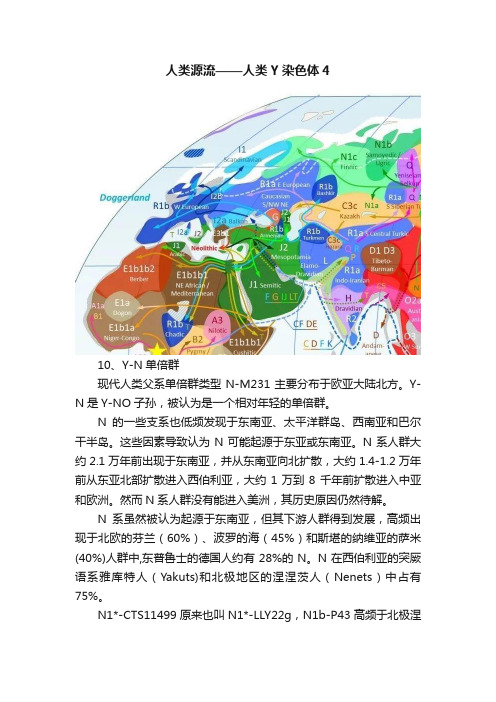
人类源流——人类Y染色体410、Y-N单倍群现代人类父系单倍群类型N-M231主要分布于欧亚大陆北方。
Y-N是Y-NO子孙,被认为是一个相对年轻的单倍群。
N的一些支系也低频发现于东南亚、太平洋群岛、西南亚和巴尔干半岛。
这些因素导致认为N可能起源于东亚或东南亚。
N系人群大约2.1万年前出现于东南亚,并从东南亚向北扩散,大约1.4-1.2万年前从东亚北部扩散进入西伯利亚,大约1万到8千年前扩散进入中亚和欧洲。
然而N系人群没有能进入美洲,其历史原因仍然待解。
N系虽然被认为起源于东南亚,但其下游人群得到发展,高频出现于北欧的芬兰(60%)、波罗的海(45%)和斯堪的纳维亚的萨米(40%)人群中,东普鲁士的德国人约有28%的N。
N在西伯利亚的突厥语系雅库特人(Yakuts)和北极地区的涅涅茨人(Nenets)中占有75%。
N1*-CTS11499原来也叫N1*-LLY22g,N1b-P43高频于北极涅涅茨人群。
N1a-M128高频于乌拉尔地区。
N1c1-M46大约有1.4万岁,是Y-N系统中最高频的支系。
他可能起源于今天的中国,随后在西伯利亚经历了严重的瓶颈,高频于雅库特人中,并向东欧方向进行第二次扩张。
N1c1主要发现于东北欧人群,尤其是芬兰 (61%), 拉普兰(斯堪的纳维亚半岛的最北端地区,53%), 爱沙尼亚 (34%), Latvia (38%), 立陶宛 (42%)和北俄罗斯 (30%), 也中低频分布于中俄罗斯 (15%), 白俄罗斯 (10%), 东乌克兰 (9%), 瑞典 (7%), 波兰 (4%) 和土耳其 (4%)。
N1c 也突出分布于说乌拉尔语种族的伏尔加-乌拉尔地区,包括Udmurts (67%)、Komi (51%)、 Mari (50%) 和Mordvins (20%),也存在于突厥语邻居中,如楚瓦什人(28%)、伏尔加鞑靼人(21%) 和巴什基尔人(17%),以及俄罗斯南部Nogais(9%) 。
(2)解读日本人的基因和起源

(2)解读日本人的基因和起源遗传学人类基因Y染色体单倍体技术近年有了很大的发展,各方对此期待甚高,希望就此解读人类进化、变迁与人种之谜。
编者对此也满怀期待,但是网上对这些研究数据的解读却是误解甚多。
最近网上有人发表了东邻日本人的基因数据,Y染色体数据如下:日本人和族的Y染色体类型中,其中O2(东北亚黄种人)--M95占31.7%,O3(华夏黄种人)--M122占20.1%,O1(华夏黄种人)--M119占3.4%;D(小黑种人)占34.7%,D1(小黑种人)占4%;C1(褐种人)-M8占8.5%;N(北欧黄种人)占1.6%。
阿伊努人的Y染色体类型中,C1(褐种人)--M8占90%,D(小黑种人)占10%。
网上的解读大多是:小黑种人成分占有41.7%,日本人是小黑种人;东北亚黄种人占31.7%,日本人是东北亚黄种人;华夏黄种人占23.5%,日本人是华夏黄种人;甚至还有说是动物基因,等等,不一而足。
这里有个对数据解读的误区,这些数据无疑是有根据的,即使有偏差也不会太大。
但是数据的解读方法与统计方法一致才不会出现缪误。
这组数据是在一定数量(如10000个)个体抽查的结果,正确的解读如下:10000个日本和族人中,具有小黑种人成分的人占有41.7%,具有东北亚黄种人成分的人占31.7%,具有华夏黄种人成分的人占23.5%,具有褐种人成分的人占8.5%;具有北欧黄种人成分的人占1.6%。
同样解读阿伊努人的Y染色体类型:10000个阿伊努人中,具有褐种人成分的人占90%,具有小黑种人成分的人占10%。
这样的解读结果只能是:日本和族是多个人种的混血民族;而由于褐种人本就是具有小黑种人的基因,所以阿伊努人基本是纯正的褐种人。
而历史的资料表明,日本人的主体是和族,另外有少数民族阿伊努族、琉球族,现今琉球族已基本被和族同化。
和族亦称“大和人”,是日本民族的主体,约占总人口的98% 以上,属蒙古人种东亚类型,其系统特征是头较短,黄皮肤,黑头发,体毛和胡须较少,身材普遍较矮。
我们对亚太地区大量群体的 Y 染色体调查研究画出了图 1 的民族

百越遗传结构的一元二分迹象李 辉【提 要】最新的分子人类学材料———Y染色体DNA能对民族系统进行精细的分析。
百越系统被发现与中国的其他系统差异很大,而与南岛语系民族(马来系统)特别是台湾语族群体相当接近。
已经研究的百越群体显示出遗传发生关系和语言文化类型的差距,这与百越的整体认同和地域分化有关。
根据数据的主成分分析得到百越民族系统遗传结构的三个特点。
11百越有单起源的遗传学迹象,可能大约三、四万年前发源于广东一带,而后慢慢扩散开来;21百越二分为以浙江为中心的东越和以版纳为中心的西越;31百越群体在发展过程中曾经由广东向东北、西北、西南三个方向迁徙。
百越接触过的许多族群也涵入了部分百越的遗传类型。
百越从广东经江西向浙江的扩散可能发生于很早以前,福建与浙江的越族群体是不同路线迁来的,在台湾原住民遗传结构中同样表现出这种二元性。
西部傣族类群是很晚从广东迁出的。
【关键词】百越 遗传结构 主成分分析 单起源迹象。
【作 者】李辉,复旦大学现代人类学研究中心博士。
上海,200433【中图分类号】Q988 【文献标识码】A 【文章编号】1004-454X(2002)04-0026-006一、遗传研究的百越范围民族学在界定民族系统的范围时,依据的是民族系统的独有特征。
长期以来,对民族进行的遗传学研究也期望找到这样的特征,但是从血型等早期的民族遗传指标中找到系统特征无异于天方夜谭。
近十来年人类基因组研究的兴起才使民族遗传研究有了希望。
线粒体DNA最先展现了全人类的系统结构①,但是由于它构建的群体系统稳定性较差而遭到了许多骂名。
对于精细的民族研究,现有的线粒体标记更是无能为力。
随后研究热点转到了Y染色体DNA上②。
这是一种纯父系遗传、信息量大、稳定、特异的绝佳遗传标记,它不但构建了稳定的发生于非洲的全人类谱系树③,在民族之间更是体现出不同层次的差异④。
我们对亚太地区大量群体的Y染色体调查研究画出了图1的民族系统亲缘树⑤。
34万年前的人类“祖父”
◆ 佟 庆/文34万年前的人类“祖父”《美国国家地理》杂志有个“基因地图项目”,让世界各地的人们寄送人体DNA样本给他们,由他们来对样本进行检测,从而确定人们之间的亲缘关系。
最近,一份奇特的DNA样本的检测结果让人们目瞪口呆。
奇怪的Y染色体来自古老的人种这份DNA样本的主人是生活在美国南卡罗来纳州的一位非洲裔美国男性,名叫佩里,最近刚刚去世。
他在世的时候,家属就把他的DNA样本送来检测。
结果发现,佩里的Y染色体上的一些遗传标记竟然与任何其他男性的Y染色体的标记都不一样!科学家认为,佩里的基因非常非常古老。
我们知道,与其他人类的常染色体不同,绝大多数Y染色体上的基因在遗传过程中不会发生变化,基因突变发生的概率很低,这使得我们可以很容易根据当代人类的DNA中有限的突变基因,去追寻我们祖先之间的相互关系。
如果两个Y染色体携带有相同的突变,那就说明它们在过去有着共同的父系祖先。
两个Y染色体携带的不同突变越多,那么它们的共同祖先就离现在越遥远,这个共同祖先就冷核聚变是可行的冷核聚变确实很难让人理解,就像木柴可以燃烧,但是我们需要先把木柴加热到燃点才行,哪有不加热就自动燃烧的木柴呢?同样,核聚变也需要高温高压才能进行核反应。
但如果有催化剂的参与,冷核聚变还是可以发生的。
通常很多难以进行的化学反应,在催化剂的作用下就变得容易多了,例如要想让异常稳定的氮气与氢气进行反应很难,没有催化剂的话,在上千摄氏度的高温和几百个大气压的高压下,都不会反应,要想让它们反应生成氨,需要在电弧的作用下才行。
但是工业上合成氨,采用铁催化剂,氮气与氢气的反应就变得容易多了,反应条件仅是400℃的温度和200个大气压。
所以,化肥的生产成本就很低了。
为什么催化剂能够让反应变得更容易?这是因为参加反应的分子之间存在静电排斥,而催化剂参与后,就消除了它们之间的排斥力。
就像两个陌生人中有个介绍人就可以消除陌生感一样。
其实在恒星的核聚变反应中就有催化剂,在恒星中,存在大量的质子,质子与质子相撞生成氦,这个几率比较小,反应速度并不快。
《中国人所带的Y染色体 从DNA上找源头》A 作者温哥华宅男
中国人所带的Y染色体 从DNA上找源头转自贴库ຫໍສະໝຸດ 作者:温哥华宅男此主题相关图片如下:
从DNA上找源头1">“国际Y染色体命名委员会”把全世界的Y染色体分为从A到R的若干大的类型,而中国人所带的,主要是O型、D型,其中D型比较古老,是棕色人种留下的基因,而O型分为O1,O2和O3三种,O1型从越南、广西方向进入中国,沿着海岸线往东北走,形成了百越民族。在距今两万年的时候,O3型中分化出了O3a3b型的一支,即是苗瑶语族的祖先。而O3型其他的人,继续向北走,成为汉、羌、藏等人的共同祖先。(耶鲁大学李辉供图)9月初,是新学期开学的日子。复旦大学校园里,到处是报到的新生,复旦大学生命学院现代人类学研究中心的谭婧泽老师则忙着带几个助手给新生做体检。这不是普通意义上的健康检查,她是要配合课题,对学生们进行体质人类学的测量,取得一批数据。“学生们来自全国各地,正好是不错的数据来源”,而建立庞大的全国范围的人群数据库,是复旦现代人类学研究中心近年来一直孜孜以求的工作。复旦的新生们以来自江浙一带的为多,但也有许多学生从云南、江西、河南、山东等省考来。而现代人类学研究中心的研究团队,主任金力是上海人,李士林是吉林人,其他同事分别来自新疆、甘肃、江苏、湖北……谭婧泽自己是宁夏人。谭婧泽是18岁就离开宁夏的。到上海之前,她先到北京,在那里上大学和读研究生。毕业后,她又到了上海自然博物馆,再到复旦。从宁夏到上海,相隔“八千里路云和月”的故乡,已成为户口本或登记表上的“籍贯”,而对于她的下一代,宁夏只是母亲的老家,是自己的上一代向上海迁徙的一个源头。那么再上一代呢?谭婧泽的父母都是宁夏人。父母的父母呢?他们怎么到宁夏的?再一代代追上去,他们的祖先又是哪里来的呢?——这个问题,总有孩子向自己的父母问起,一代一代问了好多年。我们在学校读过的历史教科书,告诉我们人是从猿进化而来,进化是从数百万年前开始的。我们也都知道,北京的周口店曾出土了生活在60万年前的“北京猿人”,似乎顺理成章的,我们都是“北京猿人”的后代。如果问今天的中国人是从哪里来的,我们也都知道黄帝大战蚩尤的传说,知道中华民族有五千年的文明史。但是,具体来说,上古的猿人是怎么变成今天的中国人的?我自己又怎么成为今天的我的,会呆在这个地方?这中间的环节又是怎样一环环连接起来的?从前的人们,有几种途径来回顾自己的来路:或是从洪荒年代口口相传下来神话传说;或是古墓遗址中挖掘出的文物器物及化石;或是前人留存下来的古籍记录。古籍记录非常有限,神话传说又太玄虚,而文物出土常常支离破碎,使我们对于我们的前世总是感觉扑朔迷离。最近20多年,人们又多了一条认识自己的路径:分子遗传学,从DNA上找自己的源头。
人类Y染色体DNA单倍型类群
目录 [隐藏]∙ 1 树形图∙ 2 单倍型类群A 和 B∙ 3 有M168 (CT)变异的单倍型类群 ∙ 4 单倍型类群 F (G, H & IJK) ∙ 5 单倍型类群 K (M9) ∙ 6 单倍型类群 NO (M214) ∙ 7 单倍型类群 P (M45) ∙ 8 单倍型类群在欧洲的时间发展 ∙ 9 同见 ∙ 10参考文献 ∙11 外部链接[编辑]树形图单倍型类群 S [编辑]单倍型类群A 和B单倍型类群A是非洲人的子单倍型类群,现代的所有单倍型类群起源点。
BT是单倍型类群A的分支。
它有两个主要谱系,单倍型类群B和CT。
定义突变分离CT(除A和B的所有单倍型类群)为M168和M294。
这些突变早于“走出非洲”的迁移。
DE突变的定义可能发生在非洲东北部,大约65,000年前。
[1] P143突变定义了单倍型类群CF。
可能发生在那时,将现代人类带至亚洲南部海岸。
亚, 密克罗尼西亚, 和玻利尼西亚▪单倍型类群C3分布▪单倍型类群D2 (M55, M57, M64。
1, M179, P12, P37。
1, P41。
1 (M359。
1), 12f2。
2)▪单倍型类群D3 (P47)▪单倍型类群E(Y-DNA)分布单倍群F和其后代分布▪单倍型类群F*分布于南部印度, 斯里兰卡, 云南, 朝鲜半岛▪单倍型类群G分布▪单倍型类群G2c1▪单倍型类群H分布45000年前分离▪单倍型类群I分布岛 I2B1 (m223)主要分布于西部, 中部,和北欧。
▪单倍型类群J分布▪单倍型类群L (M20) 分布于南亚, 中亚, 西南亚,地中海▪单倍型类群T分布单倍型类群O分布单倍型类群Q分布Q被定义由SNP M242。
认为出现在大约35000-40000年前的中亚。
单倍型类群Q的亚类型和定义变异,根据2008 ISOGG树[4]在下文提供。
ss4 bp, rs41352448, 不出现在ISOGG 2008树因为STR。
人类Y染色体DNA单倍型类群介绍2018
Y染色体DNA单倍群介绍1、Y-DNA单倍群人类Y染色体DNA单倍群由非重组DNA的Y染色体突变进行定义。
这种由许多人共享的突变称为单核苷酸多态性(SNP)。
人类Y染色体每一代大约积累两次突变。
Y-DNA单倍群的分支结构组成一个Y染色体进化树,有数百甚至数千的突变由这些不同的单倍群共享。
Y染色体的最近的共同祖先(most recent common ancestor MRCA),也被称为Y染色体亚当,是目前活着的男性的最近的男性共同祖先。
Y染色体亚当估计生活在大约236000年前的非洲。
通过研究其他瓶颈,所有欧亚大陆的人都是69000年前的一个男人的后裔。
之后一个主要的遗传瓶颈期发生在大约5000年前,今天大多数欧亚大陆的人可以追溯到5000年前的十二个祖先。
Y-DNA单倍群进化树单倍群 A & B 单倍群 A(M91)单倍群A是所有单倍群起源点。
现代所有单倍群都是单倍群A的后代,稀疏分布在非洲,主要集中在西南部的科伊桑人和尼罗河谷东北部人群。
单倍群 BT (M42,M94,M139,M299)约55000年前分,BT是单倍群A的分支单倍群B(M60)单倍群B主要分布于非洲,主要集中于俾格米人群。
详细树形图:见B单倍群文件夹单倍群 CT (P143)标识单倍群 CT的突变标记是M168和M294.包含单倍群D、E、C、F,可能88000年前在亚洲或非洲出现。
单倍群 C (M130)历史起源:C单倍群携带M130突变,来源于CF单倍群。
中国境内的C单倍群主要是C2(携带M217突变),占中国总人口比例大约为5%—10%。
其下游又可分为南北两大支,北支C2b(携带F1396突变),主要分布于蒙古族和满族等民族;南支C2c(携带F1067突变),几乎遍及全中国。
详细树形图:见C单倍群文件夹Haplogroup C (M130, M216) 分布在亚洲、大洋洲和北美等o Haplogroup C1 (F3393/Z1426)▪Haplogroup C1a (CTS11043)▪Haplogroup C1a1 (M8, M105, M131) 日本低频分布▪Haplogroup C1a2 (V20) 欧洲和尼泊尔低频分布▪Haplogroup C1b (F1370, Z16480)▪Haplogroup C1b1 (AM00694/K281)▪Haplogroup C1b1a (B66/Z16458)▪Haplogroup C1b1a1 (M356) 印度低频分布, 阿拉伯半岛和中国北部▪Haplogroup C1b2 (B477/Z31885)▪Haplogroup C1b2a (M38) 分布在印度尼西亚,新几内亚岛,美拉尼西亚,密克罗尼西亚,和玻利尼西亚▪Haplogroup C1b2b (M347, P309) 澳洲土著o Haplogroup C2 (M217, P44) 分布在欧亚大陆和北美,特别是在蒙古人,哈萨克人,通古斯人,西伯利亚人,和Na-Dené-speaking语民族单倍群DE(M1,M145,M203)约65000年前分离单倍群D (M174)详细树形图:见D单倍群文件夹∙Haplogroup D (M174) 分布在日本、中国(特别分布于西藏)和安达曼岛o D1 (CTS11577)▪D1a (Z27276, Z27283, Z29263)▪Haplogroup D1a1 (M15) 主要分布在西藏、羌族、彝族和苗瑶语人群▪Haplogroup D1a2 (P99) 主要分布在西藏、羌族、纳西族、突厥部落▪Haplogroup D1b (M55, M57, M64.1, M179, P12, P37.1, P41.1 (M359.1), 12f2.2) 主要在日本o D2 (L1366, L1378, M226.2) 菲律宾、麦克坦岛Haplogroup E (M96)详细树形图:见E倍群文件夹∙Haplogroup E (M40, M96) 分布在非洲、中东和欧洲o Haplogroup E1 (P147)▪Haplogroup E1a (M33, M132) 旧称E1▪Haplogroup E1b (P177)▪Haplogroup E1b1 (P2, DYS391p); 旧称E3▪Haplogroup E1b1a (V38) 非洲尼日尔-刚果语人群; 旧称E3a▪Haplogroup E1b1b (M215) 非洲之角,北非、中东和欧洲地中海地区; 旧称E3bo Haplogroup E2 (M75)Haplogroup F (M89)单倍群F和后代迁徙图单倍群F和后代构成了目前世界人口的90%,几乎都分布在撒哈拉以南非洲地区之外。