膜蛋白ppt
合集下载
《乙酰胆碱受体》课件

《乙酰胆碱受体》PPT课 件
乙酰胆碱受体是一类重要的膜蛋白,媒介着乙酰胆碱的信号传导,并在神经 系统中发挥关键作用。本课件将介绍乙酰胆碱受体的定义、分类、结构、功 能、与神经系统的关系、研究进展以及在药物研发中的应用。
乙酰胆碱受体的定义
乙酰胆碱受体是一种膜蛋白,可结合并传递乙酰胆碱信号,调节神经递质的 释放和神经传导。
乙酰胆碱受体与神经系统的关 系
乙酰胆碱受体与神经系统密切相关,影响着神经递质的平衡和神经信号的传 导,对神经系统的正常功能起着重要作用。
乙酰胆碱受体的研究进展
对乙酰胆碱受体的研究不断深入,揭示了其结构、功能调节以及与各种神经 系统疾病的关联,为新药研发提供了重要的理论基础。
乙酰胆碱受体在药物研发中的 应用
乙酰胆碱受体作为药物的靶点,在治疗神经系统疾病、药物依赖性和疼痛等 领域具有广泛的研究和应用价值。
乙酰胆碱受体的分类
乙酰胆碱受体分为两种类型:毛细胞膜相关的离子通道型受体(nAChR)和G 蛋白偶联受体型受体(mAChR)。
乙酰胆碱受体的结构
乙酰胆碱受体由多个蛋白亚单位组成,包括α亚单位、β亚单位、γ、δ和ε等亚 单位,形成复杂的结构。
乙酰胆碱受体的功能
乙酰胆碱受体在神经系统中调节神经递质的释放和ห้องสมุดไป่ตู้经传递,参与感觉、运 动、学习和记忆等生理过程。
乙酰胆碱受体是一类重要的膜蛋白,媒介着乙酰胆碱的信号传导,并在神经 系统中发挥关键作用。本课件将介绍乙酰胆碱受体的定义、分类、结构、功 能、与神经系统的关系、研究进展以及在药物研发中的应用。
乙酰胆碱受体的定义
乙酰胆碱受体是一种膜蛋白,可结合并传递乙酰胆碱信号,调节神经递质的 释放和神经传导。
乙酰胆碱受体与神经系统的关 系
乙酰胆碱受体与神经系统密切相关,影响着神经递质的平衡和神经信号的传 导,对神经系统的正常功能起着重要作用。
乙酰胆碱受体的研究进展
对乙酰胆碱受体的研究不断深入,揭示了其结构、功能调节以及与各种神经 系统疾病的关联,为新药研发提供了重要的理论基础。
乙酰胆碱受体在药物研发中的 应用
乙酰胆碱受体作为药物的靶点,在治疗神经系统疾病、药物依赖性和疼痛等 领域具有广泛的研究和应用价值。
乙酰胆碱受体的分类
乙酰胆碱受体分为两种类型:毛细胞膜相关的离子通道型受体(nAChR)和G 蛋白偶联受体型受体(mAChR)。
乙酰胆碱受体的结构
乙酰胆碱受体由多个蛋白亚单位组成,包括α亚单位、β亚单位、γ、δ和ε等亚 单位,形成复杂的结构。
乙酰胆碱受体的功能
乙酰胆碱受体在神经系统中调节神经递质的释放和ห้องสมุดไป่ตู้经传递,参与感觉、运 动、学习和记忆等生理过程。
蛋白质分离技术全ppt课件

第十节
蛋白质 的分离与纯化
一、 引言
二、 蛋白质(酶)分 离纯化的前处理三、蛋白质(酶来自分离 与纯化四、层析技术
五、电泳技术
六、离心技术
1
一、 引言
• 蛋白质(酶)存在于一切生物体中,是 非常重要的生物大分子。蛋白质是生物 功能的执行者,担负着生物催化、物质 运输、运动、防御、调控及记忆、识别 等多种生理功能。
化膜,暴露出疏水区域,同时又中和了电荷, 破坏了亲水溶胶,蛋白质分子即聚集而形成沉 淀。
26
Salting-in
Salting-out
溶 解 度
盐浓度
27
水化膜
++ + +
碱
+
+
++ +
酸
带正电荷蛋白质 (亲水胶体)
脱水
水化膜 碱
酸
等点电时的蛋白质 (亲水胶体)
脱水
带负电荷蛋白质 (亲水胶体)
脱水
• 盐析法应用最广的还是在蛋白质领域,已有八 十多年的历史,其突出的优点是:
• ①成本低,不需要特别昂贵的设备。 • ②操作简单、安全。 • ③对许多生物活性物质具有稳定作用。
25
⑴ 盐析的基本原理
• 蛋白质溶液为亲水溶胶体系,其稳定因素:水 化膜和电荷。
• 中性盐的亲水性大于蛋白质分子的亲水性。 • 加入大量中性盐后,夺走了水分子,破坏了水
• 3) 酶解法:利用各种水解酶,如溶菌酶、纤维素酶、蜗牛 酶和酯酶等,于37℃,pH8,处理15分钟,可以专一性地将 细胞壁分解。
• 4) 有机溶剂处理法:利用氯仿、甲苯、丙酮等脂溶性溶剂或 SDS(十二烷基硫酸钠)等表面活性剂处理细胞,可将细胞 膜溶解,从而使细胞破裂,此法也可以与研磨法联合使用。
蛋白质 的分离与纯化
一、 引言
二、 蛋白质(酶)分 离纯化的前处理三、蛋白质(酶来自分离 与纯化四、层析技术
五、电泳技术
六、离心技术
1
一、 引言
• 蛋白质(酶)存在于一切生物体中,是 非常重要的生物大分子。蛋白质是生物 功能的执行者,担负着生物催化、物质 运输、运动、防御、调控及记忆、识别 等多种生理功能。
化膜,暴露出疏水区域,同时又中和了电荷, 破坏了亲水溶胶,蛋白质分子即聚集而形成沉 淀。
26
Salting-in
Salting-out
溶 解 度
盐浓度
27
水化膜
++ + +
碱
+
+
++ +
酸
带正电荷蛋白质 (亲水胶体)
脱水
水化膜 碱
酸
等点电时的蛋白质 (亲水胶体)
脱水
带负电荷蛋白质 (亲水胶体)
脱水
• 盐析法应用最广的还是在蛋白质领域,已有八 十多年的历史,其突出的优点是:
• ①成本低,不需要特别昂贵的设备。 • ②操作简单、安全。 • ③对许多生物活性物质具有稳定作用。
25
⑴ 盐析的基本原理
• 蛋白质溶液为亲水溶胶体系,其稳定因素:水 化膜和电荷。
• 中性盐的亲水性大于蛋白质分子的亲水性。 • 加入大量中性盐后,夺走了水分子,破坏了水
• 3) 酶解法:利用各种水解酶,如溶菌酶、纤维素酶、蜗牛 酶和酯酶等,于37℃,pH8,处理15分钟,可以专一性地将 细胞壁分解。
• 4) 有机溶剂处理法:利用氯仿、甲苯、丙酮等脂溶性溶剂或 SDS(十二烷基硫酸钠)等表面活性剂处理细胞,可将细胞 膜溶解,从而使细胞破裂,此法也可以与研磨法联合使用。
高中生物-细胞膜课件

第十七页,编辑于星期五:十点 二十三分。
②化学突触是存在于可兴奋细胞之间的连接方式,它通 过释放神经递质来传导神经冲动。化学突触是相对于电 突触而言的,它们共同完成可兴奋细胞之间的通讯。
第十八页,编辑于星期五:十点 二十三分。
③间隙连接 两个细胞的质膜之间有2—3 nm的间隙又 称缝隙连接。 构成间隙连接的根本单位称连接小体或连接子。连接 子是一种颗粒结构,由6个跨膜蛋白分子构成中央为 1.5—2 nm的孔道,一些小分子可通过。 相邻细胞膜连接子两两相对,形成一个间隙连接,为 细胞间物质交换、信息传递提供直接通道。 间隙连接主要分布于上皮、平滑肌及心肌等组织细胞 间。
⑶通讯连接 通讯连接包括间隙连接、植物细胞中的胞间 连丝、神经细胞间的化学突触。
①胞间连丝是植物细胞间特有的连接方式。如图1— 1—20所示,在胞间连丝连接处的细胞壁不连续,相 邻细胞的细胞膜共同形成20 nm ~40nm的管状结构, 是物质从一个植物细胞进入到另一个植物细胞的通路, 它在植物细胞间的物质运输和信息传递中具有重要作 用。
由细胞膜上受体、G蛋白以及PIP2磷酸二酯酶〔PDE) 组成
第二十三页,编辑于星期五:十点 二十三分。
③具有酪氨酸蛋白激酶活性的受体信号通路 一些生长因子和靶细胞膜的相应受体结合后,受体 本身的酪氨酸蛋白激酶活性被激活,将细胞内靶蛋 白的酪氨酸残基磷酸化,从而引起细胞反响。
第二十四页,编辑于星期五:十点 二十三分。
第十五页,编辑于星期五:十点 二十三分。
⑵锚定连接 锚定连接在机体组织内分布很广泛。通过锚定
连接可将相邻细胞的骨架系统或将细胞与基质相连,形成 一个坚挺、有序的细胞群。
锚定连接具有两种不同形式:与中间纤维相连接的锚定连接 主要包括桥粒和半桥粒;与肌动蛋白纤维相连接的锚定连
②化学突触是存在于可兴奋细胞之间的连接方式,它通 过释放神经递质来传导神经冲动。化学突触是相对于电 突触而言的,它们共同完成可兴奋细胞之间的通讯。
第十八页,编辑于星期五:十点 二十三分。
③间隙连接 两个细胞的质膜之间有2—3 nm的间隙又 称缝隙连接。 构成间隙连接的根本单位称连接小体或连接子。连接 子是一种颗粒结构,由6个跨膜蛋白分子构成中央为 1.5—2 nm的孔道,一些小分子可通过。 相邻细胞膜连接子两两相对,形成一个间隙连接,为 细胞间物质交换、信息传递提供直接通道。 间隙连接主要分布于上皮、平滑肌及心肌等组织细胞 间。
⑶通讯连接 通讯连接包括间隙连接、植物细胞中的胞间 连丝、神经细胞间的化学突触。
①胞间连丝是植物细胞间特有的连接方式。如图1— 1—20所示,在胞间连丝连接处的细胞壁不连续,相 邻细胞的细胞膜共同形成20 nm ~40nm的管状结构, 是物质从一个植物细胞进入到另一个植物细胞的通路, 它在植物细胞间的物质运输和信息传递中具有重要作 用。
由细胞膜上受体、G蛋白以及PIP2磷酸二酯酶〔PDE) 组成
第二十三页,编辑于星期五:十点 二十三分。
③具有酪氨酸蛋白激酶活性的受体信号通路 一些生长因子和靶细胞膜的相应受体结合后,受体 本身的酪氨酸蛋白激酶活性被激活,将细胞内靶蛋 白的酪氨酸残基磷酸化,从而引起细胞反响。
第二十四页,编辑于星期五:十点 二十三分。
第十五页,编辑于星期五:十点 二十三分。
⑵锚定连接 锚定连接在机体组织内分布很广泛。通过锚定
连接可将相邻细胞的骨架系统或将细胞与基质相连,形成 一个坚挺、有序的细胞群。
锚定连接具有两种不同形式:与中间纤维相连接的锚定连接 主要包括桥粒和半桥粒;与肌动蛋白纤维相连接的锚定连
第五章 蛋白质的三维结构(二)(共45张PPT)

包括: 膜内在蛋白质: 脂锚定蛋白质:
1. 膜内在蛋白质: 第一种类型:血型糖蛋白 第二种类型: 7个跨膜肽段的膜蛋白 〔细菌紫膜红质〕
第一种类型
第二种类型---7个跨膜肽段的膜蛋白
第二种类型 ---β桶型膜蛋白
2. 脂锚定蛋白质: A 酰胺——连接豆蔻酰锚钩 B 硫酯——连接脂肪酰锚钩 C 硫醚——连接异戊烯基锚钩 D 酰胺——连接糖基磷脂酰肌醇锚钩。
反平行螺旋束
蛋白质的结构--二级结构 三.亚基相互作用的方式 序列上存在七残基重复序列。
Often found in DNA- and RNA-binding proteins. 一级结构上相邻的两个螺旋采取接近相互垂直的取向。 由4—10个β折叠股构成。 Dihedral(两面角) 三.亚基相互作用的方式 Dihedral(两面角) 序列上存在七残基重复序列。 上下型β桶:β曲折闭合而成,配 相同的,形成 封闭的二聚 反平行β片〔露面夹心〕:含3—15个β折叠股的单层反平行β折叠片,但不闭合成桶。 2.结构域类型:(根据二级结构种类和组合方式) 体结合部位位状蛋白质疏水侧链埋藏在分子内部,亲水侧链暴露在 分子外表:
疏水核心几乎全部由β折叠片和α螺旋组成。多数α螺旋 是两亲性螺旋。 5.球状蛋白质分子的外表有一个空穴〔裂光、凹槽、口袋 〕:
它常是结合底物、效应物等配体并行使生物功能的活性 部位。空穴周围分布着许多疏水侧链。
第八节 膜蛋白的结构 膜周边蛋白质:
单绕平行β桶
双绕平行β片
〔马鞍形扭曲片〕
3.全β—结构〔反平行β折叠片〕蛋白质:
由4—10个β折叠股构成。
希腊钥匙β桶:由希腊钥匙维成
反平行β桶
果冻卷β桶:类似希腊钥匙,更复杂。
类型:
1. 膜内在蛋白质: 第一种类型:血型糖蛋白 第二种类型: 7个跨膜肽段的膜蛋白 〔细菌紫膜红质〕
第一种类型
第二种类型---7个跨膜肽段的膜蛋白
第二种类型 ---β桶型膜蛋白
2. 脂锚定蛋白质: A 酰胺——连接豆蔻酰锚钩 B 硫酯——连接脂肪酰锚钩 C 硫醚——连接异戊烯基锚钩 D 酰胺——连接糖基磷脂酰肌醇锚钩。
反平行螺旋束
蛋白质的结构--二级结构 三.亚基相互作用的方式 序列上存在七残基重复序列。
Often found in DNA- and RNA-binding proteins. 一级结构上相邻的两个螺旋采取接近相互垂直的取向。 由4—10个β折叠股构成。 Dihedral(两面角) 三.亚基相互作用的方式 Dihedral(两面角) 序列上存在七残基重复序列。 上下型β桶:β曲折闭合而成,配 相同的,形成 封闭的二聚 反平行β片〔露面夹心〕:含3—15个β折叠股的单层反平行β折叠片,但不闭合成桶。 2.结构域类型:(根据二级结构种类和组合方式) 体结合部位位状蛋白质疏水侧链埋藏在分子内部,亲水侧链暴露在 分子外表:
疏水核心几乎全部由β折叠片和α螺旋组成。多数α螺旋 是两亲性螺旋。 5.球状蛋白质分子的外表有一个空穴〔裂光、凹槽、口袋 〕:
它常是结合底物、效应物等配体并行使生物功能的活性 部位。空穴周围分布着许多疏水侧链。
第八节 膜蛋白的结构 膜周边蛋白质:
单绕平行β桶
双绕平行β片
〔马鞍形扭曲片〕
3.全β—结构〔反平行β折叠片〕蛋白质:
由4—10个β折叠股构成。
希腊钥匙β桶:由希腊钥匙维成
反平行β桶
果冻卷β桶:类似希腊钥匙,更复杂。
类型:
纤维蛋白和膜蛋白构象

(存在Lys和Hyl间醇醛交联或吡啶啉结构,二硫键)
资料仅供参考,不当之处,请联系改正。
2. 胶原的形成:以I型胶原为例
羟化酶
① 羟化 : 发生在Pro ,Lys
Hyp ,Hyl (90%)
② 糖化 : Hyl-OH (葡萄糖、半乳糖) 糖苷
③ 前原胶原的形成:三条肽链交接在一起
端肽
④ 原胶原形成: 去处C-端和N-端端肽
片层之间(范德华力) 纵向有韧性,无伸展性
彩色电镜下观察从蜘蛛吐丝器吐出的丝心蛋白束(蓝色)
NH -CH-COO +
-
资料仅供参考,不当之处,请联系改正。
3
NH3+
(CH2)3
NH3+
(二) 弹性蛋白(elastin)CH: -(CH2)2— — (CH2)2 -CH
COO
N1-
存在:韧带、声带、主动脉等
资料仅供参考,不当之处,请联系改正。
(一)角蛋白 (keratin):
-角蛋白, β-角蛋白, 羽毛角蛋白, 无定形角蛋白
1. -角蛋白:头发角蛋白 构成:
-helix(0.45nm)原纤维(2nm) 微纤维(8nm)大纤维(200nm) 特性:
a 有较强的伸缩性,在热水或 碱溶液中可伸展为β-sheet
b 水不溶性(含大量的疏水aa)
c 韧性,多螺旋结构紧密
2. β-角蛋白:
资料仅供参考,不当之处,请联系改正。
例如:各种丝蛋白 代表:蚕丝中的丝心蛋白
( fibroin )。 含大量-Gly-Ser-Gly-Ala-
Gly-Ala-重复单位 不能形成-helix,多以
反平行β-sheet存在 维持力:链间(氢键);
资料仅供参考,不当之处,请联系改正。
2. 胶原的形成:以I型胶原为例
羟化酶
① 羟化 : 发生在Pro ,Lys
Hyp ,Hyl (90%)
② 糖化 : Hyl-OH (葡萄糖、半乳糖) 糖苷
③ 前原胶原的形成:三条肽链交接在一起
端肽
④ 原胶原形成: 去处C-端和N-端端肽
片层之间(范德华力) 纵向有韧性,无伸展性
彩色电镜下观察从蜘蛛吐丝器吐出的丝心蛋白束(蓝色)
NH -CH-COO +
-
资料仅供参考,不当之处,请联系改正。
3
NH3+
(CH2)3
NH3+
(二) 弹性蛋白(elastin)CH: -(CH2)2— — (CH2)2 -CH
COO
N1-
存在:韧带、声带、主动脉等
资料仅供参考,不当之处,请联系改正。
(一)角蛋白 (keratin):
-角蛋白, β-角蛋白, 羽毛角蛋白, 无定形角蛋白
1. -角蛋白:头发角蛋白 构成:
-helix(0.45nm)原纤维(2nm) 微纤维(8nm)大纤维(200nm) 特性:
a 有较强的伸缩性,在热水或 碱溶液中可伸展为β-sheet
b 水不溶性(含大量的疏水aa)
c 韧性,多螺旋结构紧密
2. β-角蛋白:
资料仅供参考,不当之处,请联系改正。
例如:各种丝蛋白 代表:蚕丝中的丝心蛋白
( fibroin )。 含大量-Gly-Ser-Gly-Ala-
Gly-Ala-重复单位 不能形成-helix,多以
反平行β-sheet存在 维持力:链间(氢键);
存在着三种类型的膜蛋白

许多生物学上重要的分子通过简单 的扩散就可进入和离开细胞,其中 包括非极性的气体(例如O2和CO2) 和疏水性的分子(例如类固醇激素、 脂溶性维生素和某些药物等)。
吲哚 甘油 尿素 色氨酸 葡萄糖
7.15 跨膜转运系统对 细胞是非常必要的
转运蛋白通常都是对某些 分子或结构上类似的分子 的基团特异的。最简单的 一类转运蛋白执行单向转 运(图a),即它们只携带 单一一种类型的溶质跨膜 转运。而许多转运蛋白可 进行两种溶质的同一方向 的同向转运(图b)或协 同转运(图c)。
7.11 存在着三种类型的膜蛋白
三种类型的膜蛋白:内在膜蛋白、外周膜蛋白和锚脂蛋白。
由于内在膜蛋白是嵌入在膜的脂双层中,所以这类蛋白的 分离需要破坏蛋白和膜脂之间的疏水相互作用。
通常都是使用一些去污剂破坏疏水相互作用。因为去污剂 在溶解膜的过程中可以取代大多数膜脂,形成可溶性的微团。
许多内在膜蛋白与它周围的脂的结合是非常紧密的,可以 利用可溶性蛋白纯化使用过的技术(如凝胶过滤、离子交换等) 将去污剂溶解的膜萃取物中的膜蛋白分离出来,纯化的内在膜 蛋白一般都保存在去污剂中,一旦除去去污剂,将导致不可逆 的蛋白凝聚和沉淀。
7.15.1通道蛋白和孔蛋 白使溶质扩散过膜
通道蛋白和孔蛋 白是带有中央亲水通 道的跨膜蛋白。一般 来说术语孔常用于细 菌,而通道常用于动 物。大小、电荷和几 何形状都合适的溶质 都可顺着浓度梯度经 通道快速地扩散。
7.15.2 被动转运是自发进行的
被动转运不需要能量驱动,被动转运也称为易化扩散。 转运蛋白的作用是加快反应的平衡,如果没有转运蛋白,单 靠扩散达到平衡非常慢。
外周膜蛋白与内在膜蛋白不同,它们与膜的 作用弱得多,通常都是通过离子键和氢键与膜脂 的极性头部或与内在膜蛋白结合的。
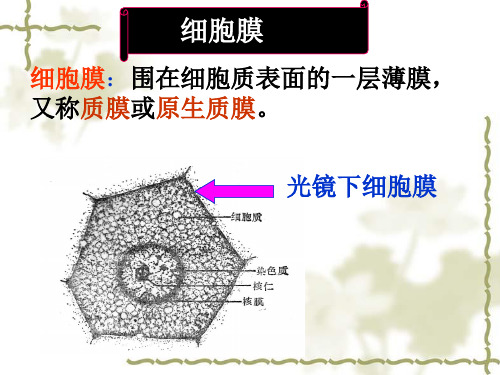
细胞膜(共162张PPT)

第二节 细胞膜及其表面
细胞膜(cell memberane):是包围在细胞外周的 一层薄膜,又称质膜(plasma membrane). 细胞膜是原始的非细胞生物演化为细胞生物的
一个转折点
单位膜(unit membrane): “二暗一明”的膜 式结构叫三层夹板式结构。
生物膜
细胞膜
细胞膜
细胞质
细胞内膜(internal membrane):除了细胞膜以外 的细胞内所有膜性结构。
1
2
3
胞质面 5
1.单 次 穿 膜: 单条a-螺旋贯穿脂质双层。
2.多 次 穿 膜: 数条a-螺旋几次折返穿越脂质双层。
跨膜 蛋白
3.非穿越性共价结合:不穿越脂质双层的全部,而与胞质侧单层 脂质的烃链结合。
4.与磷脂酰肌醇结合: 镶嵌蛋白通过自己的一个寡糖链与磷脂酰肌 醇(在非胞质面的单层)共价结合。
上提出了修正模型,认为 膜上还具有贯穿脂双层的 蛋白质通道,供亲水物质 通过。
Unit membrane modle
4. J. D. Robertson 1959
用超薄切片技术获得了清
晰的细胞膜照片,显示暗-
明-暗三层结构,它由厚约
的双层脂分子和内外表面
各厚约2nm的蛋白质构 成。单位膜模型的不足 之处在于把膜的动态结 构描写成静止的不变的。
而推测细胞膜由双层 家族性高胆固醇血症临床特点:
信号分子及其信号传导方式
脂分子组成。 2.
(二)降血脂药物:当小孩的LDL-C超过160mg/dl(正常<110 mg/dl)须要小心评估,防止心血管疾病危险性。 多附在膜的内表面,非共价地结合在镶嵌蛋白上。 NO对血管的效应可以很好地解释硝化甘油的作用,早在100年前就使用硝化甘油处理心绞痛的病人(这种绞痛是由血液不适当地流向心肌 引起的)。 斑上有中间纤维相连,中间纤维的性质因细胞类型而异,桥粒中间为钙粘素(desmoglein及desmocollin)。
细胞膜(cell memberane):是包围在细胞外周的 一层薄膜,又称质膜(plasma membrane). 细胞膜是原始的非细胞生物演化为细胞生物的
一个转折点
单位膜(unit membrane): “二暗一明”的膜 式结构叫三层夹板式结构。
生物膜
细胞膜
细胞膜
细胞质
细胞内膜(internal membrane):除了细胞膜以外 的细胞内所有膜性结构。
1
2
3
胞质面 5
1.单 次 穿 膜: 单条a-螺旋贯穿脂质双层。
2.多 次 穿 膜: 数条a-螺旋几次折返穿越脂质双层。
跨膜 蛋白
3.非穿越性共价结合:不穿越脂质双层的全部,而与胞质侧单层 脂质的烃链结合。
4.与磷脂酰肌醇结合: 镶嵌蛋白通过自己的一个寡糖链与磷脂酰肌 醇(在非胞质面的单层)共价结合。
上提出了修正模型,认为 膜上还具有贯穿脂双层的 蛋白质通道,供亲水物质 通过。
Unit membrane modle
4. J. D. Robertson 1959
用超薄切片技术获得了清
晰的细胞膜照片,显示暗-
明-暗三层结构,它由厚约
的双层脂分子和内外表面
各厚约2nm的蛋白质构 成。单位膜模型的不足 之处在于把膜的动态结 构描写成静止的不变的。
而推测细胞膜由双层 家族性高胆固醇血症临床特点:
信号分子及其信号传导方式
脂分子组成。 2.
(二)降血脂药物:当小孩的LDL-C超过160mg/dl(正常<110 mg/dl)须要小心评估,防止心血管疾病危险性。 多附在膜的内表面,非共价地结合在镶嵌蛋白上。 NO对血管的效应可以很好地解释硝化甘油的作用,早在100年前就使用硝化甘油处理心绞痛的病人(这种绞痛是由血液不适当地流向心肌 引起的)。 斑上有中间纤维相连,中间纤维的性质因细胞类型而异,桥粒中间为钙粘素(desmoglein及desmocollin)。
大学医学细胞生物学细胞膜课件
呼吸窘迫综合征 又称新生儿肺透明膜病。指新生儿出生后不久即出现进行 性呼吸困难和呼吸衰竭等症状,主要是由于缺乏肺泡表面 活性物质所引起,导致肺泡进行性萎陷,患儿于生后4~ 12小时内出现进行性呼吸困难、呻吟、发绀、吸气三凹征 (吸气时胸骨上窝、锁骨上窝、肋间隙出现明显凹陷), 严重者发生呼吸衰竭。发病率与胎龄有关,胎龄越小,发 病率越高,体重越轻病死率越高。
1.脂肪酸链的长度及不饱和程度 2.胆固醇与磷脂的比值 3.卵磷脂与鞘磷脂的比值 4.膜蛋白 5.温度,离子强度,PH
细胞膜的流动性异常与疾病 新生儿呼吸窘迫症是由于质膜中卵磷脂/鞘磷脂比值 过低,限制了膜流动性,影响了O2/CO2的交换. 衰老,动脉硬化是由于质膜中卵磷脂/鞘磷脂比值过 低,限制了膜流动性.
四.细胞膜的功能
(一)保护 (二)物质运输
重点
小分子和离子的穿膜运输 大分子和颗粒物质的膜泡运输
(三) 信息的跨膜传递
难点
1.小分子和离子的穿膜运输 根据是否需要ATP的参入,分为: 被动转运:不消耗细胞的代谢能(ATP), 顺浓度梯度的运输。 主动转运:消耗细胞的代谢能(ATP), 逆浓度梯度的运输。
3.1935年J.F.Danielli和H.Davson发现细胞的表面张力显著低于 油水界面的表面张力而提出片层结构模型 4.20世纪50年代末J.D.Robertson用电镜观察细胞膜发现都呈 “两暗一明”而提出单位膜模型
5.20世纪60年代,红外光谱等技术证明膜蛋白不是β折叠而是α螺旋 而提出液态镶嵌模型
将以下化合物按膜通透性递增次序排列: 核糖核酸、钙离子、葡萄糖、乙醇、氮分子、水 氮分子(小而非极性) >乙醇(小而略有极性)>水(小 而极性)>葡萄糖(大而极性)>钙离子(小而带电荷) >核糖核酸(很大而带电荷)
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
2.1 基于去垢剂的结晶研究(in surfo methods) 去垢剂也称表面活性剂(surfactant),一般情况下其亲水性质要比脂类分子强,因此形成的微团类型 属于 Type 1,其结构特征为微团的烷基尾巴聚集在一起,被亲水的头部所包裹.在膜蛋白结晶方法 学中, 以去垢剂结晶为代表的 In surfo 结晶法是最为经典也最为传统的方法.
1.1 两亲性分子自组装原理 两亲性分子的亲水基与疏水基与水有相反的作用倾向:亲水基倾向于进入水中,而疏水基则倾向于远离 水相.二者平衡的结果是:两亲性分子将在水的表面聚集并覆盖在水面上,其中每个分子的亲水基都伸 入水中,而疏水基则指向水相之外. 但当两亲性分子的浓度进一步增大,水表面完全被单层两亲性分子覆盖.此时继续增加浓度,将迫使这 些分子不得不在水中开始发生自组装,形成亲水基与疏水基隔离的胶团结构. 形成这种胶团的最低浓度被称为临界胶束浓度(critical micelle concentration,CMC).超过了 CMC 后,两亲 性分子通常能进行分子自组装形成圆柱状、圆锥状、锲子状等结构,这些组装成的微团按照形状的不同 可分为三种类型
2.1.1 去 垢 剂 微 团 结 晶 法 (detergent micellesmethod). In surfo 结晶法中去垢剂微团结晶法是最常用的方法,Michel 等应用此方法获得了世界上第一个膜蛋 白结构———紫细菌光反应中心结构. 该方法的原理简单来说是利用去垢剂提取膜蛋白,使膜蛋白与其形成的复合物 PDC 可以像可溶性蛋 白那样直接用气相扩散(悬滴法和坐滴法)和透析等方法来结晶.PDC 是一个很复杂的网络结构,包括 膜蛋白分子、去垢剂和水缓冲液,其中去垢剂分子会贯穿在晶格中形成网络结构,而膜蛋白分子间 的相互作用是形成晶格的最重要因素.晶格对于PDC 的大小和形状有严格的要求,小的 PDC 一般可 以形成有序的膜蛋白晶格堆积,因此去垢剂在结晶过程中的选择至关重要.非离子型去垢剂和两性 去垢剂已成功应用于膜蛋白结晶中,其中前者的应用更为广泛.装配 PDC 过程中也可添加一种或几 种去垢剂,有时根据靶蛋白的性质也可以添加一些脂类分子或者其他化学分子.
膜蛋白三维结晶
Three-dimensional Crystallization of Membrane Protein
C目录 ONTENTS 第一章 膜蛋白三维结晶原理 第二章 膜蛋白结晶方法研究进展
01 膜蛋白三维结晶原理
The principle of three-dimensional membrane protein crystallization
膜蛋白结构的特殊性,使其一旦脱离天然磷脂双分子结构就会发生聚集变性,导致其三维结构改变和功 能的丧失.因此,要保证膜蛋白的结晶,首先必须找到一种能够高效地从天然环境中提取膜蛋白并能稳 定其结构功能的方法.使用两亲性分子诸如去垢剂、脂类分子是目前最主要的手段,其中去垢剂应用最 为广泛,但近年来脂类分子也逐渐受到了研究者越来越广泛的关注.
单击此处输入标题
Type 0 (Lamellar phase)、 Type 1 (Normal micelle)、 Type 2(Inverse micelle). 为了满足能量最小原理,这些类
型的微团分别将其分子的疏水部 分与水相隔离起来.
Type 1、Type 2 分别是以去垢剂 和脂类分子为典型代表,Type 0 则两者都可以装配形成.
体结构研究内容,而且为光合
作用捕光机制和光保护机制的 研究奠定了重要的结构基础.
02 膜蛋白结晶方法研究进展
2 膜蛋白结晶方法研究进展
基于两亲性分子自组装以及能与膜蛋白分子装配成复合物结晶的原理,膜蛋白结晶研究目前主要有 两种方法———基于去垢剂的 in surfo 结晶法和基于脂类的 in cubo 结晶法.
三维晶体Ⅰ 具有典型的层状结构,
主要通过基于类分子的方法获
得,其膜蛋白分子间的相互作用
靠疏水表面来介导,因此增加了
膜蛋白间的相互接触而可以产生
蛋白质分子的密集堆积,通常能 得到很好的衍射数据. 对于 X 射线单晶衍射来说需要膜 蛋白的三维晶体,最通常的晶体 类型是三维晶体Ⅱ
单击此处输入标题
除上述膜蛋白三维晶体基本类 型外,Liu 和Chang 等在研究菠
菜主要捕光复合物(LHC-Ⅱ)结
构中发现了膜蛋白三维晶体堆 积的第三种方式:首先由 60 个膜蛋白 LHC-Ⅱ分子、脂分子 及色素分子组装成一个正二十
面体的空心球体结构,然后将
该二十面体结构作为基本单位
进行晶体的周期排列.这种全
新的堆积方式完全不同于前面 所述的Ⅰ型和Ⅱ型膜蛋白晶体, 可作为第三类膜蛋白三维晶体 类型(3-D crystal type Ⅲ).该重大 研究发现不但丰富了膜蛋白晶
1.2 膜蛋白提取原理及其晶体分类
去垢剂分子的结构特征是具有一 个亲水的头部和一个疏水的尾 巴. 当把去垢剂加入到含有脂双层的 溶液中后,去垢剂分子首先进入 膜表面和水相之间,导致局部膜 组分变形.当越来越多的去垢剂 进入后,膜组分被去垢剂饱和最 终崩塌与去垢剂分子一起形成蛋 白质 - 去垢剂混合微团复合物 (proteindetergent complex, PDC), 膜蛋白包裹于 PDC 之中后被提取 出来.
单击此处输入标题
根据后续结晶实验中使用的方法 可将膜蛋白晶体分为 3 类: 二维晶体(2-D crystal)、 三维晶体Ⅰ(3-D crystal type Ⅰ) 三维晶体Ⅱ(3-D crystal type Ⅱ).
二维晶体 实质上是在脂类分子重
新装配成脂双层的膜平面上形成 的,因此只能通过2-D 电子显微镜 观察.
其中 Type1 去垢剂分子包围的区 域是被去垢剂烷基尾巴所充满而
不是真空的区域,去垢剂分子通
常有一个单链烷基,其含有大量 的亲水基促进了去垢剂组装成. Type 1 微团,反之脂类分子含有 单链或者双链烷基,因此有比去
垢剂更强的疏水性质,这样促使 其自组装成 Type 2 微团.
单击此处输入标题
1.1 两亲性分子自组装原理 两亲性分子的亲水基与疏水基与水有相反的作用倾向:亲水基倾向于进入水中,而疏水基则倾向于远离 水相.二者平衡的结果是:两亲性分子将在水的表面聚集并覆盖在水面上,其中每个分子的亲水基都伸 入水中,而疏水基则指向水相之外. 但当两亲性分子的浓度进一步增大,水表面完全被单层两亲性分子覆盖.此时继续增加浓度,将迫使这 些分子不得不在水中开始发生自组装,形成亲水基与疏水基隔离的胶团结构. 形成这种胶团的最低浓度被称为临界胶束浓度(critical micelle concentration,CMC).超过了 CMC 后,两亲 性分子通常能进行分子自组装形成圆柱状、圆锥状、锲子状等结构,这些组装成的微团按照形状的不同 可分为三种类型
2.1.1 去 垢 剂 微 团 结 晶 法 (detergent micellesmethod). In surfo 结晶法中去垢剂微团结晶法是最常用的方法,Michel 等应用此方法获得了世界上第一个膜蛋 白结构———紫细菌光反应中心结构. 该方法的原理简单来说是利用去垢剂提取膜蛋白,使膜蛋白与其形成的复合物 PDC 可以像可溶性蛋 白那样直接用气相扩散(悬滴法和坐滴法)和透析等方法来结晶.PDC 是一个很复杂的网络结构,包括 膜蛋白分子、去垢剂和水缓冲液,其中去垢剂分子会贯穿在晶格中形成网络结构,而膜蛋白分子间 的相互作用是形成晶格的最重要因素.晶格对于PDC 的大小和形状有严格的要求,小的 PDC 一般可 以形成有序的膜蛋白晶格堆积,因此去垢剂在结晶过程中的选择至关重要.非离子型去垢剂和两性 去垢剂已成功应用于膜蛋白结晶中,其中前者的应用更为广泛.装配 PDC 过程中也可添加一种或几 种去垢剂,有时根据靶蛋白的性质也可以添加一些脂类分子或者其他化学分子.
膜蛋白三维结晶
Three-dimensional Crystallization of Membrane Protein
C目录 ONTENTS 第一章 膜蛋白三维结晶原理 第二章 膜蛋白结晶方法研究进展
01 膜蛋白三维结晶原理
The principle of three-dimensional membrane protein crystallization
膜蛋白结构的特殊性,使其一旦脱离天然磷脂双分子结构就会发生聚集变性,导致其三维结构改变和功 能的丧失.因此,要保证膜蛋白的结晶,首先必须找到一种能够高效地从天然环境中提取膜蛋白并能稳 定其结构功能的方法.使用两亲性分子诸如去垢剂、脂类分子是目前最主要的手段,其中去垢剂应用最 为广泛,但近年来脂类分子也逐渐受到了研究者越来越广泛的关注.
单击此处输入标题
Type 0 (Lamellar phase)、 Type 1 (Normal micelle)、 Type 2(Inverse micelle). 为了满足能量最小原理,这些类
型的微团分别将其分子的疏水部 分与水相隔离起来.
Type 1、Type 2 分别是以去垢剂 和脂类分子为典型代表,Type 0 则两者都可以装配形成.
体结构研究内容,而且为光合
作用捕光机制和光保护机制的 研究奠定了重要的结构基础.
02 膜蛋白结晶方法研究进展
2 膜蛋白结晶方法研究进展
基于两亲性分子自组装以及能与膜蛋白分子装配成复合物结晶的原理,膜蛋白结晶研究目前主要有 两种方法———基于去垢剂的 in surfo 结晶法和基于脂类的 in cubo 结晶法.
三维晶体Ⅰ 具有典型的层状结构,
主要通过基于类分子的方法获
得,其膜蛋白分子间的相互作用
靠疏水表面来介导,因此增加了
膜蛋白间的相互接触而可以产生
蛋白质分子的密集堆积,通常能 得到很好的衍射数据. 对于 X 射线单晶衍射来说需要膜 蛋白的三维晶体,最通常的晶体 类型是三维晶体Ⅱ
单击此处输入标题
除上述膜蛋白三维晶体基本类 型外,Liu 和Chang 等在研究菠
菜主要捕光复合物(LHC-Ⅱ)结
构中发现了膜蛋白三维晶体堆 积的第三种方式:首先由 60 个膜蛋白 LHC-Ⅱ分子、脂分子 及色素分子组装成一个正二十
面体的空心球体结构,然后将
该二十面体结构作为基本单位
进行晶体的周期排列.这种全
新的堆积方式完全不同于前面 所述的Ⅰ型和Ⅱ型膜蛋白晶体, 可作为第三类膜蛋白三维晶体 类型(3-D crystal type Ⅲ).该重大 研究发现不但丰富了膜蛋白晶
1.2 膜蛋白提取原理及其晶体分类
去垢剂分子的结构特征是具有一 个亲水的头部和一个疏水的尾 巴. 当把去垢剂加入到含有脂双层的 溶液中后,去垢剂分子首先进入 膜表面和水相之间,导致局部膜 组分变形.当越来越多的去垢剂 进入后,膜组分被去垢剂饱和最 终崩塌与去垢剂分子一起形成蛋 白质 - 去垢剂混合微团复合物 (proteindetergent complex, PDC), 膜蛋白包裹于 PDC 之中后被提取 出来.
单击此处输入标题
根据后续结晶实验中使用的方法 可将膜蛋白晶体分为 3 类: 二维晶体(2-D crystal)、 三维晶体Ⅰ(3-D crystal type Ⅰ) 三维晶体Ⅱ(3-D crystal type Ⅱ).
二维晶体 实质上是在脂类分子重
新装配成脂双层的膜平面上形成 的,因此只能通过2-D 电子显微镜 观察.
其中 Type1 去垢剂分子包围的区 域是被去垢剂烷基尾巴所充满而
不是真空的区域,去垢剂分子通
常有一个单链烷基,其含有大量 的亲水基促进了去垢剂组装成. Type 1 微团,反之脂类分子含有 单链或者双链烷基,因此有比去
垢剂更强的疏水性质,这样促使 其自组装成 Type 2 微团.
单击此处输入标题