第七章 糖代谢 第二次课
第七章 糖代谢

糖代谢第一节糖的分解代谢一.酵解(glycolysis , Embden-Meyerhof Pathway EMP):酵解小结:1.调节 (1) 己糖激酶(葡萄糖激酶):受G—6—P反馈抑制(2)磷酸果糖激酶:ATP抑制、F—2 .6—2P激活F—2.6—2P对磷酸果糖激酶的激活作用(3)丙酮酸激酶:ATP、乙酰CoA抑制2. ATP的生成:酵解总反应Glu+2Pi+2ADP+2NAD+→2丙酮酸+2ATP+2NADH+2H2O+2H+3. 底物水平磷酸化:二.丙酮酸的去路三.三羧酸循环(TCA循环, 柠檬酸循环,Krebs循环)1.丙酮酸的氧化脱羧2.TCA途径及参加的酶3.TCA小结(1)TCA途径部位(2)TCA总反应式(3)生成的ATP(4)TCA的调节A.丙酮酸脱氢酶系: ATP、NADH、乙酰辅酶A↓NAD+ 、CoA ↑B. 柠檬酸合成酶:ATP、NADH、琥珀酰CoA↓C.异柠檬酸脱氢酶系: ATP↓ ADP、NAD+↑D.α—酮戊二酸脱氢酶系: NADH、琥珀酰CoA↓ AMP↑4. TCA的意义5. TCA的回补反应:A. 丙酮酸→ 草酰乙酸B. 磷酸烯醇式丙酮酸→ 草酰乙酸B. 一些氨基酸可回补TCA :Glu , Asp 等四.葡萄糖彻底氧化为CO2和H2O需经历几个阶段1.几个阶段?2.1分子葡萄糖彻底氧化生成的ATP分子数?五.乙醛酸循环1.途径2.意义六.磷酸戊糖途径(pentose phosphate pathway) 细胞内葡萄糖的主要利用途径1.磷酸戊糖途径及参加的酶2.磷酸戊糖途径的调节:3.途径的意义:糖的异生作用节什么是糖的异生作用及生理意义1.糖异生作用:由非糖物质合成葡萄糖的过程2. 生理意义:(1)补充糖供应的不足,维持血糖稳定(80—120 mg / 100ml血)(2)消除肌肉中乳酸和丙酮酸等的积累糖异生途径1.丙酮酸→ 磷酸烯醇式丙酮酸(PEP)2.PEP → 1.6-2P-F ( EMP逆反应 )3. 1.6-2P-F + H2O → 6-P-F + Pi(果糖1.6-二磷酸酶)4.6-P-F → 6-P-G5. 6-P-G → G + Pi糖异生的前体: 丙酮酸、乳酸、甘油、各种生糖氨基酸及TCA中间代谢物等乳酸的再利用和Cori循3. 糖异生与酵解的协同调控(1)三个不可逆反应(2)2.6--二磷酸果糖的协同调控糖原的合成与分解节糖原的合成与分解都是从糖原的非还原端开始 糖原的分解代谢1.糖原的结构2.糖原主链的断裂(糖原磷酸化酶限速酶)3. 去分枝作用(去分枝酶)4.G-1-P → G-6-P (磷酸葡萄糖变位酶)5.G-6-P + H2O → G + Pi (葡萄糖-6-磷酸酶 肝脏) 二.糖原分解的调节: 糖原磷酸化酶的调节1.别构调节:AMP↑ ATP↓ G-6-P↓2.共价调节 : 磷酸化 / 脱磷酸化胰高血糖素和肾上腺素对糖原磷酸化酶的激活三.糖原的生物合成(Glycogen Synthesis)1.UDP—葡萄糖的合成(UDP—葡萄糖焦磷酸化酶)2.糖原的合成(糖原合酶 限速酶)糖原合酶的调节 :别构调节 G-6-P ↑共价调节:磷酸化 / 脱磷酸化3.糖原的分枝作用(糖原分枝酶)四.糖原代谢的调节1.糖原合成与分解的协同调控(1)葡萄糖对糖原合成与分解的调节(2)磷酸化调节 2. 激素对糖代谢的调节。
生物化学 第七章 糖类与糖类代谢

β -淀粉酶
两种淀粉酶性质的比较
α-淀粉酶 不耐酸,pH3时失活 耐高温,70C时15分 钟仍保持活性 广泛分布于动植物和 微生物中。 -淀粉酶 耐酸,pH3时仍保持活性 不耐高温,70C15分钟 失活 主要存在植物体中
3、R-酶(脱支酶)
水解α-1,6糖苷键,将α及β-淀粉酶作用于 支链淀粉最后留下的极限糊精的分支点或支链淀粉 分子外围分支点水解,产生短的只含α-1,4糖苷 键的糊精,使之可进一步被淀粉酶降解。
ATP CH2 OH H O H OH H OH OH H OH 葡萄糖
CH2 O H OH
1,6-二磷酸果糖
(
2)第二阶段:1, 6-二磷酸果糖 3-磷酸甘油醛
CH2OPO3H2 C O 96%
H2O3PO
CH2 O H OH
CH2OPO3H2 OH OH H 醛缩酶
CH2OH 磷酸二羟丙酮
第二节
双糖和多糖的酶促降解
一、双糖的酶促降解
二、多糖的酶促降解
一、双糖的酶促降解
蔗糖+H2O 蔗糖+UDP
蔗糖酶
蔗糖合酶
葡萄糖+果糖 果糖+UDPG
2 葡萄糖
麦芽糖+H2O
-乳糖 +H2O
麦芽糖酶
β-半乳糖苷酶
葡萄糖+半乳糖
二、多糖的酶促降解
淀粉的酶促降解 糖原的酶促降解
(一)、淀粉的酶促降解
1、磷酸化酶
催化淀粉非还原末端的葡萄糖残基转移给P,生成G-1-P, 同时产生一个新的非还原末端,重复上述过程。 直链淀粉
支链淀粉
G-1-P
G-1-P + 磷酸化酶极限糊精
糖代谢优秀PPT课件学习PPT教案

丙酮酸的去路
糖酵解途径
(无氧)
葡萄糖 (有氧或无氧) 丙酮酸
(有氧)
乙酰 CoA
乳酸 乙醇
三羧酸 循环
丙酮酸的无氧还原
(1) 乳酸发酵(lactic fermation)
动物,藻类、乳酸菌
G +2ADP+ 2Pi 2ATP+2H2O
2ห้องสมุดไป่ตู้酸 +
NAD
(2)酒精发酵(alcoholic fermation)
异柠檬酸脱氢酶
CHCOO H 异柠檬酸脱氢酶
CH2
HOO CHCOOH
O CHCOOH O CHCOOH
H 异柠檬酸
草酰琥珀酸
α-酮戊二酸
这是三羧酸循环的第一次氧化脱羧反应,
异柠檬酸脱氢酶是第二个限速酶。
(4)α-酮戊二酸氧化脱羧反应
COOH
CH2COOH
O
CH2 +
CCOOHH
NAD+
HSCoA
( (一一))糖糖酵的解途无径氧(g酵ly解col及ys生is)醇发酵
(Embden Meyerhof Parnas EMP)
1.定义:在无氧的条件下,葡萄糖或糖原分解成丙酮酸,并释 放少量能量的过程称为糖的无氧分解。这一过程与酵母菌使 糖发酵的过程相似,又称为糖酵解。 1897年发现酵母提取液可以使蔗糖发酵生成乙醇,打开了 现代生物化学的大门…直到1940年,糖酵解的全过程才被 全面揭示,在这项研究中,有许多科学家作出了巨大贡献, 其中德国生物化学家G.Embden O.Meyerhof的贡献最大, 因此糖酵解途径又叫Embden-Meyerhof(EMP途径)。
• 2、β淀粉酶只能从非还原端开始水解。 • 3、水解淀粉中的α-1,6糖苷键的酶是α-1,6
第七章 糖代谢
K2=3250
在植物光合组织中蔗糖磷酸合酶的活性较高,而非光合组 织中蔗糖合酶的活性较高。这是目前认为可能在光合组织中合 成蔗糖的主要途径。
(二)淀粉的合成:
存在于植物体内,尤其是谷类、豆类、薯类 作物的籽粒和贮藏组织都含丰富的淀粉。
淀粉合成中的糖基供体有ADPG、UDPG, 主要是ADPG。
合成分两阶段进行,先合成直链淀粉,然后 分支形成支链淀粉。
二、反应过程 反应可分为两个阶段: 第一阶段:氧化阶段,生成NADPH+H+和 CO2;由6-磷酸葡萄糖直接脱氢脱羧生 成磷酸戊糖; 第二阶段:非氧化阶段,一系列基团转 移反应;磷酸戊糖分子再经重排最终又 生成6-磷酸葡萄糖。
第一阶段:氧化阶段
1、脱氢反应:6-磷酸葡萄糖脱氢酶以NADP+ 为辅酶,催化6-磷酸葡萄糖脱氢生成6-磷酸 葡萄糖酸δ内酯,不可逆。
生物合成的供氢体
脂肪酸、胆固醇和类固醇化合物 的生物合成,均需要大量的NADPH。
0 R-CH2-C-R’
=
OH R-CH2-CH-R’ NADP+
R-CH2-CH2-R’
NADPH + H+
H R-C=C-R’
3、磷酸戊糖途径与疾病
神经精神病
(neuropsychiatric disorder)
第六节 糖的合成代谢
一、光合作用 二、糖异生途径 三、蔗糖和多糖的生物合成
一、光合作用
• 光合作用是糖合成代谢的主要途径。 • 绿色植物、光合细菌或藻类等将光能转变成化学 能的过程,即利用光能,由CO2和H2O合成糖类化 合物并释放出氧气的过程,称为光合作用。 • 光合作用的总反应式可表示如下: 光能 • n CO2 + n H2O (CH2O)n + n O2 叶绿体 糖类化合物
07第七章糖代谢

新陈代谢包括生物体内所发生的一切合成和分解作用。一般说来, 生物体把从外界摄取的含低能量的较简单的化合物,转化成高能量的复杂 的细胞结构的化合物。也就是说在新陈代谢所包括的合成代谢与分解代谢 中,前者是吸能反应,后者是放能反应,合成与分解代谢既表现着生物体 内物质分子的改变,又体现出生物体在生命活动中能量的变化。
2020/6/8
(三)新陈代谢的研究方法
代谢研究的方法很多。代谢研究方法的选择,要考虑 研究的对象和所要解决的问题。常用的方法有以下几种。 1、同位素示踪法
同位素示踪法(isotopic tracertechnique)也称为体内(in vivo) 水平的代谢研究。原子序数相同,化学性质相同,但质量不 同的元素叫做某元素的同位素,即同位素的质子数相同,中 子数不同。同位素有稳定同位素和放射性同位素两种;天然 同位素都是稳定同位素。放射性同位素的核能够自己发生变 化,放出带有电荷的粒子或不带电荷的射线。稳定同位素和 放射性同位素都可用于代谢研究,但放射性同位素要比稳定 同位素应用方便些。
在机体的生命过程中合成代谢(同化作用)与分解代谢(异化 作用)的主次关系也是相互转化的,由于这种转化就使生物个体的 发展呈现出生长、发育和衰老等不同的阶段。可见,生命机体通过 新陈代谢获得它所必需的能量;通过新陈代谢建造和修复生物体; 通过新陈代谢完成遗传信息的贮存、传递和表达过程,使得生物物 种世代繁衍、生生不息。总之,合成为分解准备了物质前提,外部 物质变为内部物质;同时,分解为合成提供必需的能量,内部物质 又能转变为外部物质。
第七章糖类代谢案例
麦芽糖分子结构(葡萄糖α-1,4-葡萄糖苷)
CH2OH O H H OH H OH
CH2OH O OH H 1 H OH H H H OH
H O
4
H OH
β-半乳糖
α-葡萄糖
乳糖分子结构(葡萄糖β,α-1,4-半乳糖苷)
乳糖和麦芽糖有半缩醛羟基,因此具有还原性。 • 蔗糖没有游离的半缩醛羟基,是非还原糖。
•
三、多糖
多糖属于非还原性糖。 按生物来源:植物多糖、动物多糖和微生物多糖; 按生物功能:贮存多糖和结构多糖; 按组成的不同:同多糖和杂多糖。
(一) 同多糖
• 1. 淀粉(分为直链 淀粉和支链淀粉)
• 直链淀粉以(14) 糖苷键聚合而成。呈螺 旋结构,遇碘显紫蓝色。
• 支链淀粉中除了 (1→4)糖苷键构成糖链以外,在支点 处存在 (1→6)糖苷键,分子量较高。遇碘显紫红色。
糖的分类
根据其水解产物的情况,糖主要可分为: • 单糖 (monosacchride):仅包含一个多羟基醛或多羟基 酮单位;是构成寡糖和多糖的基本单位。 • 寡糖 (oligosacchride):由2-20多个单糖通过糖苷键连接 而成的糖类物质; • 多糖 (polysacchride):由多个单糖(20个以上)单位通 过糖苷键连接而构成的糖类物质; • 结合糖 (glycoconjugate):糖和非糖物质共价结合而成 的复合物。
二、糖酵解的生物化学过程
糖酵解的全过程在细胞质(cytoplasm)中进 行,共10步,可分为两大阶段: • 酵解准备阶段(第1-5步); • 产能阶段(第6-10步)。
糖酵解途径(1)
将ATP上的磷酸基团转移到受体上的酶称为激酶(kinase)。 己糖激酶(肝外组织)与葡萄糖激酶(肝脏)为同工酶,对葡萄糖的 Km值分别为0.1mmol/L、5~10mmol/L;葡萄糖激酶是一种诱导酶, 葡萄糖浓度较高时才起作用。 己糖激酶是糖酵解过程中第一个调节酶,受产物的别构抑制。 该反应为糖酵解途径的第一个限速步骤。
第七章 糖代谢—糖的合成

糖异生的生理意义
(一)维持血糖浓度恒定 (二)补充肝糖原,回收乳酸能量(乳酸循环) 补充肝糖原,回收乳酸能量(乳酸循环) (三)协助氨基酸代谢 (四)调节酸碱平衡(乳酸异生为糖) 调节酸碱平衡(乳酸异生为糖)
五、糖代谢的紊乱
糖尿病:体内胰岛素缺乏,血糖分解缓慢, 糖尿病:体内胰岛素缺乏,血糖分解缓慢,导致血糖过高由尿 排出。患者的肝糖原合成和糖的氧化能力均降低,脂代谢增加 排出。患者的肝糖原合成和糖的氧化能力均降低, 产生过多的酮体(乙酰乙酸、 羟丁酸 丙酮), 羟丁酸、 ),并发酮血症 ,产生过多的酮体(乙酰乙酸、β-羟丁酸、丙酮),并发酮血症 和酮尿症。 和酮尿症。
G-1-P
UTP (a) ADP ATP
G-6-P
Pi
G
糖原的合成与分解代谢
第六节 糖的异生
* 概念
糖异生(gluconeogenesis)是指从非糖化 糖异生(gluconeogenesis)是指从非糖化 (gluconeogenesis) 合物转变为葡萄糖或糖原的过程。 合物转变为葡萄糖或糖原的过程。
合成具有α 糖苷键的有分支的糖原, 3. 合成具有α-1,6糖苷键的有分支的糖原,反应由 分支酶催化 催化: 分支酶催化:
糖原 n+1
UDP (c)
Pi
(d)
UDPG UDPG
PPi (b)
(a) 磷酸葡萄糖变 位酶 UDPG焦磷酸 (b) UDPG焦磷酸 化酶 (c) 糖原合成酶 分支酶) (分支酶) (d) 磷酸化酶 脱支酶) (脱支酶)
2、UDPG在糖原合成酶催化下合成新糖原 UDPG在糖原合成酶催化下合成新糖原 新的葡萄糖残基加在糖原引物的非还原末端的葡萄糖 残基的第四碳的羟基上, 糖苷键。 残基的第四碳的羟基上,形成α-1,4糖苷键。UDP被 , 糖苷键 被
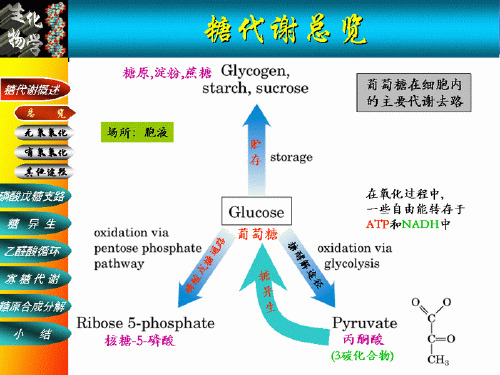
第七章 糖代谢

1 第七章 糖代谢 第一节 糖代谢概述 1.1、糖代谢的类型 p311
1.2、糖代谢的生物学意义 p311 1、提供能量; 2、提供合成其它物质的碳源; 3、作为生物体的结构物质; 4、作为信息分子。 第二节 糖的分解代谢 2.1、糖的无氧分解 p311 1、概况:
2、反应历程:3 阶段共11步或12步反应,均在胞液中进行。其中第1、2阶段共10步反应称为糖酵解途径(EMP途径)。P311-316 Ⅰ、第一阶段:葡萄糖 → 磷酸丙糖,共5步反应。 P312-313
糖的有氧分解 糖的无氧分解 磷酸戊糖途径 糖异生作用 由单糖合成寡糖及多糖
糖的分解代谢
糖的合成代谢 糖代谢 寡糖及多糖的降解 2 磷酸葡萄糖异构酶 3
此阶段共需耗2分子ATP。⑴、⑶两步反应均为不可逆反应,为EMP途径中的重 要调控位点。催化酶均为别构酶。其中ATP虽然是磷酸果糖激酶Ⅰ的底物,但对该 酶具有别构抑制作用(反馈抑制)。 经此阶段,1C6 → 2C3。 注意:书写反应式时,应写明底物、产物、酶、辅助因子、可逆性等。 Ⅱ、第二阶段:3-磷酸甘油醛 → 丙酮酸,共5步反应,为EMP途径的重要阶段。 P313-314 4 此阶段经历了一次脱氢,产生1分子NADH+H+,2次底物水平磷酸化,共产生2 Mg2+或Mn2+ 5
分子ATP。第⑽步反应可视为不可逆反应,由丙酮酸激酶(别构酶)催化,为EMP途径的重要调控位点。1,6-二磷酸果糖(磷酸果糖激酶Ⅰ的产物)对丙酮酸激酶具有别构激活作用(前馈激活)。 Ⅲ、第三阶段: 丙酮酸的还原。 P316
整个无氧分解途径如下: p315-316 乳酸脱氢酶 6 7 3、糖无氧分解的特点: p314-316 ⑴、无氧参与 ⑵、产能少,共生成4分子ATP,消耗2分子ATP ,即1分子G经EMP途径净生 成2分子ATP 。 ⑶、经一次脱氢,1分子G可产生2分子NADH+H+。 ⑷、调控部位:反应⑴、⑶、⑽均为不可逆反应,催化酶己糖激酶、磷酸果糖激酶(限速酶)、丙酮酸激酶均为别构酶。 4、生物学意义 为机体供能,由于产能少,只能应付急需(厌氧生物除外)。 2.2、糖的有氧分解 p311-316,p321-325 1、概述:
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
乙酰CoA+草酰乙酸 草酰乙酸 乙酰
TCA循环 循环
⑵ 柠檬酸异构化生成异柠檬酸 H2O
H2C COOH HO C COOH H CH COOH
H2C COOH C COOH HC COOH
H2C COOH H C COOH
COOH HO CH
柠檬酸
顺乌头酸 异柠檬酸
顺乌头酸酶 柠檬酸 异柠檬酸
异柠檬酸脱氢酶
异柠檬酸+NAD+ 异柠檬酸
α-酮戊二酸 +CO2+NADH+H+ 酮戊二酸
TCA循环 循环
酮戊二酸氧化脱羧生成琥珀酰辅酶 ⑷ α-酮戊二酸氧化脱羧生成琥珀酰辅酶 酮戊二酸氧化脱羧生成琥珀酰辅酶A HSCoA NAD+ H2C COOH
H2C COOH CH2
O C SCoA
CH2 O C COOH
6-磷酸 磷酸 果糖激 酶-2
1,6-二磷酸果糖 二磷酸果糖
2,6-二磷酸果糖 二磷酸果糖
丙酮酸激酶
原料增多 时,促进 转化 * 别构调节 别构激活剂: 别构激活剂:1,6-双磷酸果糖 双磷酸果糖 别构抑制剂: 别构抑制剂:ATP, 丙氨酸
丙氨酸 丙酮酸, 丙酮酸, 相当于产物, 相当于产物,产物 抑制酶活性
称之未-科综合征,乙醇会妨碍 称之未 科综合征,乙醇会妨碍VB1的 科综合征 吸收及促进其排泄? 吸收及促进其排泄?
Why? ?
丙酮酸的脱氢酶复合体 多酶复合体:是催化功能上有联系的几种酶通过非 多酶复合体:是催化功能上有联系的几种酶通过非
连接彼此嵌合形成的复合体。 共价键连接彼此嵌合形成的复合体 共价键连接彼此嵌合形成的复合体。
糖酵解途径
三羧酸循环
胞浆
细胞质
CO2+H2O+ATP
糖的有氧氧化
第一阶段:丙酮酸的生成(胞浆) 第一阶段:丙酮酸的生成(胞浆)
第二阶段: 第二阶段:丙酮酸氧化脱羧生成乙酰 CoA(线粒体) (线粒体)
NAD+ , HSCoA
CO2 , NADH + H+
丙酮酸
丙酮酸脱氢酶复合体
乙酰CoA 乙酰
HSCoA
丙酮酸脱氢酶复合体的组成
NAD+
酶 E1:丙酮酸脱氢酶
辅酶
TPP(来自 B1) (来自V S 硫辛酸( 硫辛酸( L ) E2:二氢硫辛酰胺转乙酰酶 HSCoA 长期酗酒者:缺乏V 糖有氧氧化抑制, S 长期酗酒者:缺乏 B1 →糖有氧氧化抑制,糖酵解增 糖有氧氧化抑制 机体酸中毒→严重猝死 强→机体酸中毒 严重猝死 机体酸中毒 E3:二氢硫辛酰胺脱氢酶 FAD, NAD+
TCA循环 循环
异柠檬酸氧化脱羧生成α-酮戊二酸 ⑶ 异柠檬酸氧化脱羧生成 酮戊二酸
H2C COOH
NAD+ HC COOH H2C COOH
H2C COOH CH2 O C COOH
α-酮戊二酸
HO C COOH H
异柠檬酸 NADH+H+
HC COOH O C COOH
草酰琥珀酸
CO2 关键酶 关键酶 关键酶 关键酶
延胡索酸 (fumarate) 延胡索酸酶
H2C COOH HO CH COOH
苹果酸 (malate)
延胡索酸 + H2O
苹果酸
苹果酸脱氢生成草酰乙 脱氢生成草酰乙酸 ⑻ 苹果酸脱氢生成草酰乙酸
TCA循环 循环
NAD+
NADH+H+
H2C COOH HO C COOH H
苹果酸脱氢酶
H2C COOH
NAD GDP+Pi GTP NADH+H+
NAD+ NADH+H+
③
⑥
FAD
CO2 ④
⑤ CoASH CO2 CoASH
小 结
三羧酸循环的要点(1,2,3,4): : 三羧酸循环的要点
经过一次三羧酸循环, 经过一次三羧酸循环, 消耗1分子乙酰 发生1次底物水平磷酸化 生成GTP) 次底物水平磷酸化(生成 消耗 分子乙酰CoA; 发生 次底物水平磷酸化 生成 分子乙酰 2次脱羧 生成 分子 次脱羧,生成 分子CO2 次脱羧 生成2分子 3个关键酶催化 个不可逆反应 个关键酶催化3个不可逆反应 个关键酶催化 4次脱氢 分子 次脱氢(1分子 分子NADH+H+) 次脱氢 分子FADH2和3分子 分子 关键酶有: 关键酶有: 柠檬酸合酶 α-酮戊二酸脱氢酶复合体 酮戊二酸脱氢酶复合体 异柠檬酸脱氢酶
FAD
FADH2
H CH COOH H CH COOH
琥珀酸 (succinate) 琥珀酸脱氢酶
HOOC CH HC COOH
延胡索酸 (fumarate)
琥珀酸 + FAD
延胡索酸 +FADH2
TCA循环 循环
延胡索酸水化生成苹果酸 生成苹果 ⑺ 延胡索酸水化生成苹果酸
H2O
HOOC CH HC COOH
课后作业:记住糖酵解的反应途径。 课后作业:记住糖酵解的反应途径。 预习案例7-1 预习案例
Glu
E1
G-6-P
F-6-P
ATP ADP E1:己糖激酶 己糖激酶
E2 F-1, 6-2P ATP ADP
磷酸
丙酮
3-磷酸 磷酸
糖 酵 解 的 代
E2: 6-磷酸果糖激酶 磷酸果糖激酶-1 磷酸果糖激酶 E3: 丙酮酸激酶
TCA循环 循环 速度降低
乙酰-CoA 乙酰 浓度增加
高水平的乙酰CoA激活 激活 高水平的乙酰
产生更多的草酰乙酸
丙酮酸羧化酶
草酰乙酸主要来自糖,糖供应不足时, 草酰乙酸主要来自糖,糖供应不足时,草 酰乙酸缺乏, 会怎么样? 酰乙酸缺乏,TAC会怎么样?例如糖尿病 会怎么样
O C COOH
草酰乙酸 苹果酸 (malate)
(oxaloacetate)
苹果酸 + NAD+
草酰乙酸 + NADH+H+
H2O
H2O
② ①
NADH+H+ NAD+ CoASH H2O
② ⑧
⑦
H2O FADH2
①柠檬酸合酶 ②顺乌头酸酶 ③异柠檬酸脱氢酶 ④α-酮戊二酸脱氢酶复合体 酮戊二酸脱氢酶复合体 琥珀酰CoA合成酶 ⑤琥珀酰 合成酶 ⑥琥珀酸脱氢酶 ⑦延胡索酸酶 ⑧苹果酸脱氢酶 +
* 反应部位
所有的反应均在线粒体中进行。 所有的反应均在线粒体中进行。 线粒体中进行
TCA 简 图
1953年 汉斯·克雷布斯 克雷布斯, 1953年 汉斯 克雷布斯,李普曼 因发现柠檬酸循环而获奖
汉 斯 · 克 雷 布 斯 ( Hans Adolf Krebs,1900.8.25——1981.11.22 ) 生于德国希尔德斯海姆, 1918——1923年在德国多所大 年在德国多所大 学学医, 学学医 , 1926 —— 1930年间 年间 成为瓦尔堡的助手, 成为瓦尔堡的助手 , 因纳粹上 台而移居英国, 台而移居英国 , 在谢菲尔德大 学主持生物化学研究所。 1945 学主持生物化学研究所 。 年任该校生化学教授, 年任该校生化学教授 , 1954年 年 去牛津执教
TCA循环 循环
乙酰CoA与草酰乙酸 缩合形成柠檬酸 ⑴ 乙酰 与草酰乙酸 缩合形成柠檬酸
乙酰辅酶A 乙酰辅酶A
关键酶 CH3CO~SCoA ~
柠檬酸合成酶
H2C COOH HO
HSCoA 柠檬酸 (citrate) 柠檬酸 + CoA-SH
H2O
C COOH H2C COOH
O C COOH H2C COOH
大肠肝菌中 其中每一个酶都有其特定的催化功能, 其中每一个酶都有其特定的催化功能,都有其催化 丙酮酸脱氢酶系 的电镜图 活性必需的辅酶。 活性必需的辅酶。
大肠肝菌中 丙酮酸脱氢酶系 的示意图
转乙酰酶
丙酮酸脱氢酶 二氢硫辛酰胺 脱氢酶
1. α-羟乙基 羟乙基-TPP的生成 的生成 羟乙基 CO2 2.乙酰硫辛酰 乙酰硫辛酰 胺的生成 NADH+H+ 5. NADH+H+ 的生成 NAD+ CoASH 3.乙酰 乙酰CoA 乙酰 的生成 4.硫辛酰胺的生成 硫辛酰胺的生成 E1丙酮酸脱氢酶 E2二氢硫辛酰胺转乙酰酶 E3二氢硫辛酰胺脱氢酶
H2C COOH CH2 O C SCoA GDP+Pi
琥珀酰CoA 琥珀酰 ATP 琥珀酰CoA + GDP + Pi 琥珀酰
HSCoA
H2C COOH H2C COOH
琥珀酸
GTP
ADP
底物水平 磷酸化反应
琥珀酸+ 琥珀酸 GTP + CoA-SH
⑹ 琥珀酸氧化脱氢生成延胡索酸
TCA循环 循环
α-酮戊二酸 酮戊二酸
+ NADH+HCO 2 关键酶
琥珀酰CoA 琥珀酰
α-酮戊二酸脱氢酶系 α-酮戊二酸 + CoA-SH+ NAD+ 酮戊二酸 琥珀酰CoA + C O2 + NADH+H+ 琥珀酰
TCA循环 循环
琥珀酰CoA转变为琥珀酸 ⑸ 琥珀酰 转变为琥珀酸 琥珀酰CoA合成酶 合成酶 琥珀酰
糖的有氧氧化与糖酵解 糖的有氧氧化与糖酵解
葡萄糖→→……→→丙酮酸 乳酸(糖酵解 丙酮酸 丙酮酸→乳酸 糖酵解 糖酵解) 葡萄糖
丙酮酸 CO2+H2O+ATP
线粒体 胞浆 细胞
(糖的有氧氧化) 糖的有氧氧化) 糖的有氧氧化