重组的类型、机制与转座(self)
DNA重组和转座

第一节 DNA重组(recombination)一)、DNA重组1、概念:是指由于不同DNA链的断裂和连接而产生的DNA片段的交换和重新组合,形成新的DNA分子的过程。
2、意义:重组是遗传学的灵魂,没有重组就没有生物的进化;没有重组也就没有现代的分子克隆技术二)、DNA重组的类型1、同源重组(Homologous Recombination)2、位点特异性重组(Site-specific Recombination)3、DNA的转座(transposition)同源重组一、概述1、定义:两个DNA分子同源序列之间进行的重组2、条件:(1)两个DNA分子有同源序列(相关的酶可以用任何一对同源序列为底物)(2)两个DNA分子必须紧密接触3、发生:真核生物: 非姐妹染色单体交换相对应的区域。
原核生物: 依赖recA蛋白,并形成 Holliday 结构Holiday模型(1964年)二、大肠杆菌同源重组的分子基础(一)RecA1、作用:可促进单链同化或单链吸收RecA具有使DNA单链置换双链中同源链的能力2、单链同化发生的三个条件(1)其中一个DNA必须存在单链区(2)其中一个DNA必须有一个自由3’末端(3)此单链区和3’末端必须位于两分子之间互补的区域内(二)RecBCD复合体1、酶的活性:(1)核酸酶(2)解旋酶(3)ATPase2、作用:在CHi位点处产生含3`游离末端单链。
(三)CHi位点——RecBCD识别的靶位点5`GCTGGTGG3`3`CGACCACC5`是重组频率较高的部位E.coli每隔约5~10kb有一个拷贝,Holliday 结构的形成:1. 同源序列排列在一起;2. 酶切。
通过核酸酶和RecBCD蛋白复合体的作用在一对同源DNA上产生切口;3.入侵。
含有3’端切口的ssDNA被recA蛋白包裹形成recA蛋白-ssDNA细丝;RecA-ssDNA细丝寻找相对的DNA双螺旋上的相应序列。
分子生物学:重组和转座

转座子的特点
转座子是不必借助同源序列就可以移动的片断,即转座作用与供体 和受体的序列无关;
原核生物和真核生物都有转座子; 转座序列可沿染色体移动,甚至在不同染色体间跳跃(跳跃基因) 转座子对基因组而言是一个不稳定因素,它可导致宿主序列删除、
倒位或易位,并且其在基因组中成为“可移动的同源区”。位于 不同位点的两个拷贝转座子之间可以发生交互重组,从而造成基 因组不同形式的重排。 有些转座子与基因组的关系犹如寄生,它们的功能只是为了自身 的扩增与繁衍,因此被称为是自私的DNA。
转座中涉及的机制依赖于DNA 链的切割和重接,因此 (branch migration)
Cro蛋白抑制C I基因的转录,它占优势噬菌体即进入繁殖周期,并导致宿主细胞裂解。
与重组过程联系起来。 二者含有共同的核心序列15bp(O区)。
DNA转座子上携带作为重组位点的DNA序列,以及参与重组的蛋白质的基因(转座酶基因)。 同源重组 (homologous recombination )或普遍性重组(generalized recombination )
复制式转座 如 Tn3
非复制式转座 如 Tn10
类病毒反转录转座子/反转录病毒
带有反向终端重复序列.反向终端 重复序列嵌入在较长的正向排列的 重复序列(长末端重复序列, long terminal repeats LTR)中;
带有靶位点重复序列; 类病毒反转录转座子编码两种移位所需的蛋白:转座酶和反转录酶。 类病毒反转录转座子和反转录病毒的区别在于:反转录病毒的基因
径的歧化做好准备。
位点特异性重组与同源重组的区别
带有2个基因ORF1和ORF2;
同源重组 (homologous recombination )或普遍性重组(generalized recombination )
06.第六章-遗传重组的分子机理
• Mu的DNA是线型的,两端没有粘性末端,而是类 似于IS的序列,并有与转座有关的基因A和基因B , 其整合方式与λ噬菌体不同,不是位点专一性的整 合和切除,而是类似于转座因子,其末端常常带 有一小段宿主DNA,因而可以引起转导。
model): DNA双链的断裂与重接
3)Holliday模型:异源双链
(heteroduplex)的断裂与重接
4)Meselon-Radding 模型:
1.Holliday 模型
a) 同源染色体联会
b) 内切酶切割非姊妹染 色单体DNA
c) 交换重接形成交联桥 结构(cross-bridge
structure)
A: phe- try- tyr- × B: met- his-
苯丙AA 色AA 酪AA
甲硫AA 组AA
↓
原养型菌落
phe+ try+ tyr+ met+ his+
问题:是接合引起的?是转化引起的?
U型管实验
• 但在这里,结果却获 得了原养型菌株,说 明有一种可通过滤膜 的过滤性因子(FA), 细菌不必直接接触即 可进行基因转移
遗传学讲义 第六章 遗传重组的分子基础
中国海洋大学 生命学院 汪小龙 xiaolong@
1、遗传重组的类型
(1) 同源重组(homologous recombination)
又叫普遍性重组(generalized recombination) ,大范围同源序 列对等交换。真核生物减数分裂中同源染色体联会,非姊妹 染色单体之间的交换就是同源重组。需要重组蛋白因子参与, 如E.coli. 的RecA.参与重组,又叫依赖于RecA的重组(RecAdependent recombination);
重组和转座32ppt课件

转座酶:transposase 解离酶:Resolvase
2. 非复制型转座 (nonreplicative transposition)
Nonreplicative transposition allows a transposon to move as a physical entity from a donor to a recipient site. This leaves a break at the donor site, which is lethal unless it can be repaired.
“We are sadly ignorant of the organization of the chromosome and of the possible types of changes in this that may occur to the chromosome as a whole or at the locus level”.
四、模板选择(copy choice)性重组
适用于RNA病毒,在这种重组中,聚合酶 从一个模板转换到另一个模板来合成RNA, 结果新合成的分子将含有两个不同亲本的遗 传信息
第二节 同源重组
第三节 转座(transposition)
一、原核转座子的类型
1.插入序列(insertion sequences , IS): IS家族的结构:
靶DNA,再将转座子连接到靶DNA的凸出单链上,最后填 补空缺完成转座。
The direct repeats of target DNA flanking a transposon are generated by the introduction of staggered cuts whose protruding ends are linked to the transposon.
第二十三章重组和转座
第七节 真核生物的转座因子
一、玉米中的控制因子
1938年Marcus Rhoades首次发现不稳定突变等
位基因(unstable mutant allele),即一种回 复突变率很高的等位基因。不稳定是取 决于不连锁的Dt基因的存在。
McClintock。1940~1950描述了大量的控制因子
A1 A1 dt dt
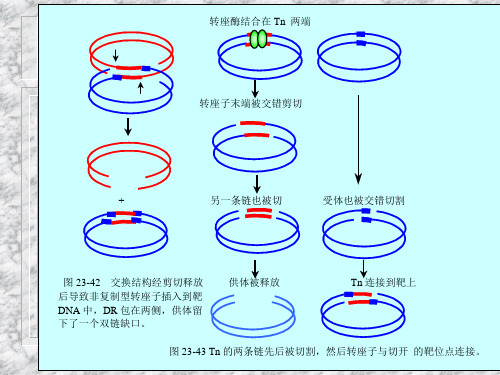
转座酶结合在 Tn 两端 转座子末端被交错剪切
+
另一条链也被切
受体也被交错切割
图 23-42 交换结构经剪切释放 后导致非复制型转座子插入到靶 DNA 中,DR 包在两侧,供体留 下了一个双链缺口。
供体被释放
Tn 连接到靶上
图 23-43 Tn 的两条链先后被切割,然后转座子与切开 的靶位点连接。
复制型转座模型解释了 (1) 复制性转座在转座后原来的位置上保留原
LTR 反转录病毒 gag
LTR pol int env
δ gag int
pol
δ
Ty
TyA
TyB
LTR gag int
pol LTR
Copia
ORF
pol
L1
ORF1
ORF2
图 23-72 病毒家族的反转录子末端有重复顺序内部有 开放读框
无LTR的反转录转 座子通过切开靶位 点双链,提供了引 物末端。反转录转 座子作为模板合成
Copia(果蝇)
Pol Ⅲ 转录 的加 工假 基
因
LINES L1(哺乳动物)
末端
长末端重复序列
无末端重复序列
靶重复序列
4~6 bp
7~12bp
阅读框
反转录酶和/或整合酶 无(不编码和转座有关
DNA的重组

有些细菌在自然条件下不发生转化或转化 效率很低,但在实验室中可以人工促使转化。 例如大肠杆菌,用高浓度Ca2+处理,可诱导细 胞成为感受态,重组质粒得以高效转化。
转化过程涉及细菌染色体上10多个基因编码的功能。 转化过程涉及细菌染色体上10多个基因编码的功能。如:感受态因、 10多个基因编码的功能 感受态因、 与膜结合的DNA结合蛋白、自溶素、多种核酸酶等 DNA结合蛋白 与膜结合的DNA结合蛋白、自溶素、多种核酸酶等。感受态因子诱导与 感受态有关的蛋白质表达,自溶素使与膜结合的DNA结合蛋白、 DNA结合蛋白 感受态有关的蛋白质表达,自溶素使与膜结合的DNA结合蛋白、核酸酶 裸露,当游离DNA与膜结合的DNA结合蛋白结合后, DNA与膜结合的DNA结合蛋白结合后 裸露,当游离DNA与膜结合的DNA结合蛋白结合后,核酸酶使其中一条 链降解,另一条链被吸收,与染色体DNA重组。 DNA重组 链降解,另一条链被吸收,与染色体DNA重组。
3.细菌的转导 转导(transduction)是通过噬菌体将细菌基因 从供体转移到受体细胞的过程。 转导有两种类型 普遍性转导(generalized transduction):是指宿 主基因组任意位置的DNA成为成熟噬菌体颗 DNA 粒DNA的一部分而被带入受体菌。 局限性转导(specialized transduction):某些温 和噬菌体在装配病毒颗粒时将宿主染色体整合 部位的DNA切割下来取代病毒DNA。
小鼠: J λ C λ3 小鼠:Vλ2 人: Vλ~ 300 J λ C λ6 λ 小鼠重链基因族( 号染色体 号染色体) 小鼠重链基因族(12号染色体)
细菌的结合作用
traS和traT基因编码表面排斥蛋白,阻止F+细胞之间的转移,F+细胞的性菌毛与F-细胞 基因编码表面排斥蛋白,阻止F 细胞之间的转移, 细胞的性菌毛与F
第二十三章重组和转座

•A1: 控制色素形成 •Dt: 控制产生斑点
第二十三章重组和转座
Marcus Rhoades分析了一种墨西哥玉米的穗, 此穗来自于一种籽粒有颜色的纯种自花受粉的玉米, 但它在后代中表现出一种意外的双因子杂种修饰性 孟德尔分离比 的植原物来,的自品交系产可生能了为上A1A述1d比td例t,。突变后产生A1a1Dtdt
第二十三章重组和转座
2020/12/9
第二十三章重组和转座
u 复制型转座模型解释了 u (1) 复制性转座在转座后原来的位置上保留原
有的Tn; u (2) 在新位置上转座子的两端出现正向重复靶
序列; u (3)转座过程中出现共合体。
第二十三章重组和转座
六.非复制型转座 七.Tn10的转座具有多项控制 u 1.“ 多拷贝抑制”(multicopy inhibition)
第二十三章重组和转座
u 这样a1成为首次发现的不稳定突变等位 基因(unstable mutant allele)的例子。即一 种回复突变率很高的等位基因。然而这种 等位基因的不稳定是取决于不连锁的Dt基 因的存在。一旦回复突变的发生,它们就 变得稳定了;即Dt基因能离开A1基因,这时 A1表型不再改变。这样a1表型是由一个缺陷 型的转座因子的插入而产生也就顺理成章 了(缺陷的转座因子自己并不能移动)。 Dt的缺乏使得表型保持稳定。
•
第二十三章重组和转座
第二十三章重组和转座
第二十三章重组和转座
第八节 反转录病毒和反转录子
u 一.反转录病毒(retroviruses) u (一) 反转录病毒的生活史 u 反转录子(retroposons) u 反转录转座子(retrotransposons) u 1. 反转录病毒基因组的结构与功能 u 2. 反转录病毒的整合模型 u 3. 反转录病毒可转录细胞的序列 u 二.酵母的Ty因子(transponson yeast)
重组的类型机制与转座

对进化生物学的影响
揭示物种进化的机
制
重组和转座是物种进化的重要驱 动力之一,研究它们有助于深入 了解物种进化的机制和过程。
解析生物多样性的
形成
重组和转座在生物多样性形成中 发挥重要作用,通过研究它们可 以更好地解析生物多样性的形成 机制和过程。
探索生命起源和演
化
通过研究重组和转座,可以更深 入地探索生命起源和演化的奥秘 ,了解生命的本质和演化规律。
。
转座酶
转座子移动需要转座酶的作用 ,这些酶能够切割转座子与 DNA的连接点,并将其插入
新的位置。
转座机制
转座子可以插入到基因内部或 基因间,影响基因的表达和功 能,也可能导致基因突变和重
排。
03
转座
转座的分类
转座子
可自主复制的转座元件,能在基因组内不同位置复制 和移动。
逆转座子
需依赖病毒或质粒进行复制的转座元件,通常从RNA 转录本中转录出cDNA并插入基因组。
THANKS
感谢观看
对医学和生物技术的意义
疾病发生机制的研究
重组和转座与许多疾病的发生和发展密切相关,研究它们有助于深 入了解疾病的发生机制和发展过程。
药物设计和治疗策略的制定
通过研究重组和转座,可以发现新的药物靶点和治疗策略,为药物 设计和治疗提供新的思路和方法。
生物技术的开发和利用
重组和转座是生物技术研究和应用的重要领域之一,通过研究它们 可以促进生物技术的开发和利用,推动相关领域的发展。
重组的类型、机制与转座
目 录
• 重组的类型 • 重组的机制 • 转座 • 重组与转座的关系 • 重组与转座的研究意义
01
重组的类型
同源重组
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
基因转变分为两种:染色单体转变和 半染色单体转变 减数后分裂:该异常分离时减数分裂后 有丝分裂的产物,故又称减数后分离 怎么解释这种现象呢?突变是不可能的, 因为他们的频率比这些基因的正常突变 频率高很多;
(2)Holliday模型
同源染色体联会(图 中只显示四条染色单 体中的二条非姐妹染 色体单体) 两条方向相同的单链 被切断; 游离端的氢键断裂, 离开互补单链 游离端交叉地和断裂 单链连接
(gene conversion)
真菌类减数分裂的正常分裂:重组通常是交互 的,例如一个杂合体中,如果以染色体把基因 A交给他的同源染色体,则同源染色体也会把a 基因交回给他。则一个位点上的两等位基因分 离时,子囊应该呈现 4:4分离
基因转变现象
真菌类的异常分离现象:1930年德国遗传学家 Winkler在链孢霉的重组产物中发现异常的5: 3(约0.06%)和6:2(约0.05%)的分离比。
链孢霉的生活史
分生孢子(n) 菌丝 (n) 子实体 交配型A 核融合 合子核(2n) 减数分裂I 减数分裂II
有丝分裂
交配型B
萌发(n) 子囊孢子(n)
脉孢霉减数分裂
非姐妹染色单 体未发生交换
+ + + + -
+
+ + + +
-
+
-
MI
M II
-
+ + +
+ + -
+ +
-
-
-
+
+
+ -
n
2、3非姐妹染色 单体发生交换
转座作用的机制
转座时发生的插入作用有一个普遍的特征, 那就是受体分子中有一段很短的(3-12bp)、 被称为靶序列的DNA会被复制,使插入的转 座子位于两个重复的靶序列之间。 不同转座子的靶序列长度不同,但对于一 个特定的转子来说,它所复制的靶序列长 度都是一样的,如IS1两翼总有9个碱基对的 靶序列,而Tn3两端总有5bp的靶序列。
(二)位点特异性重组
(site-specific recombination)
重组发生在特殊的位点上:只涉及特定位 置的短同源区或特定的碱基序列之间; 由特定重组蛋白因子识别专一性DNA序列: 重组时发生精确的切割和连接反应; 属于非同源特定片段间的重组。最典型的 如:λ噬菌体的att位点整合到大肠杆菌 的基因组中;
一、重组的概念及类型
1.重组概念 广义/细胞学水平上:由于独立分配或交换而在 后代中出现的新基因的组合过程; 狭义/分子水平上:基因的交换或重排而导致的 新基因的组合过程,即2个DNA分子间或一个DNA 分子的不同部位间,通过断裂和重接,交换DNA片 段从而改变基因的组合和序列. 2. 重组类型: (1)同源重组(Homologous recombination) (2)位点特异性重组 (3)异常重组
(Homologous recombination)
1.同源重组的概念:指DNA同源序列间的重 组.常涉及大片段同源DNA序列之间的交换。 是重组中的普遍形式,又称普遍性重组 (generalized recombination) (1)交互重组:多数真核生物减数分裂过程 中,同源染色体的非姐妹染色体单体或DNA分 子间相互交换对等的部分; (2)单向重组:在细菌转化、接合和普遍性 转导中,仅受体发生重组,供体并未发生重 组。(依赖RecA重组)
Ac-Ds系统调节色素基因C的机制
C基因位于玉米的九号染色体短臂上, AcDs位于C基因附近; 当C基因附近有Ac而无Ds时,C基因有活性, 玉米籽粒有色素形成; 当Ac不存在,而Ds因子插入、固定在C基因 处, C基因失活,玉米籽粒无色素形成; 当Ac存在时,虽有Ds因子插入、固定在C基 因处, 但胚胎发育过程中,有些细胞中的 Ds因子会因Ac的活化而发生切离转座,C 基因恢复活性,这些细胞仍能合成色素;
(三)异常重组 ( Illegitimate recombination)
完全不依赖于序列间的同源性而使一段 DNA序列插入到另一段中。 形成重组分子时往往依赖于DNA复制而 完成重组过程,因此又称复制性重组 转座因子(transposable element): 细胞中能改变自身位置的一段DNA 序列
根据该模型重组始终是交互的,如在一 个杂合体,如果一条染色体把基因A交 给同源染色体,则同源染色体必然把基 因a反过来交给他;
要求两染色单体在相应的位点上断裂, 然后以新的组合连接起来,或者从分子 水平上要求两DNA分子在同一碱基对间 断裂,然后互相以新的组合连接起来, 否则重组产物的长度就会改变;
连接成半交叉,称为 holliday结构 分支点移动呈 “十字 型” “十字型”的两臂旋 转形成中空的十字结 构
不 同 解 离 方 式
•经修补后连接形成重组和非重组的异源DNA链
Holliday模型 对基因转变机制的解释
Holliday模型中无论哪种解离方式,都 会产生两个异源的DNA双链 异源双链有关的核苷酸链分别来自不同 的亲本,如 g+ X g-,两者有一对碱基 之差 g+为:A C A G T g-为 A C A T T T G T C A T G T A A
见书p206-207 由于重组出现了完全野生型的孢子对++, 但没有重组的对应产物——双突变型的 孢子对pdx pdxp,但是,尽管pdxp位点出 现异常分离,其右侧紧密连锁的标记基 因pdx位点显示正常分离 Winkler把真菌中不规则分离的现象解释 为同源染色体联会时,一个基因使其相 对的位置上的基因发生相应的变化所致, 称为基因转变;
转座可被分为复制性和非复制性两大类。 在复制性转座中,所移动和转位的是 原转座子的拷贝。转座酶(transposase) 和解离酶(resolvase)分别作用于原始 转座子和复制转座子。TnA类转座主要是 这种形式。 在非复制性转座中,原始转座子作为 一个可移动的实体直接被移位,IS序列、
Mu及Tn5等都以这种方式进行转座。
(2)转座子(transposon, Tn)
转座子(transposon, Tn)是一类较大的 转座因子,除了含有与它的转座作用有关的 基因外,还带有某些抗药性基因(或其他宿 主基因)。因此又称复合式转座因子,转座 往往使宿主菌获得有关基因的特性; 两翼往往是两个相同或高度同源的IS序列, 表明IS序列插入到某个功能基因两端时就可 能产生复合转座子。一旦形成复合转座子, IS序列就不能再单独移动,因为它们的功能 被修饰了,只能作为复合体移动。
Duplication of the DNA sequence at a target site,when a transposon is inserted. The duplicated sequences are shown in red. These sequences are generally only a few base pairs long, so their size (compared with that of a typical transposon) is greatly exaggerated in this drawing.
-
-
异常分裂比6:2
+ +
+ + + +
+ + -
+ + -
+
+
-
MI
M II
-
异常分裂比5:3
+
+ + + +
+ + -
+ + -
+
-
+
-
MI
M II
-
异常分裂比 3: 1 : 1 : 3
+ + + + -
+
+ + + +
+
-
-
MI
M II
-
基因转变的显著特点
子囊中所形成的4个孢子对中,同一孢子 对的两个孢子基因型不同; 异常分离的子囊中,约有30%与其相邻的 两侧标记连锁基因之间发生交换和重组; 在有基因转变的子囊中,基因转变和遗传 重组都发生在同样两条染色单体的子囊比 例竟高达90%。即基因转变和重组是密切 相关的。
第十二章 重组与转座
引言
可遗传的变异是生物适应和进化的基础; 遗传物质发生变异的来源包括两个方面: 突变:基因结构或染色体的数目、结构改变; 重组:染色体或基因序列发生重排及新的组合; 基因重组是遗传的基本现象; 1. 无论高等真核生物,还是细菌、病毒都存在; 2. 不仅在减数分裂,而且在高等生物体细胞中; 3. 不只在核基因,而且在细胞质基因之间; 重组是遗传学的灵魂; 1. 没有重组就没有生物的进化; 2. 没有重组就没有现代的分子克隆技术
二.同源重组的分子机制
交叉学说与断裂重接模型
Holliday模型
(1)交叉学说与断裂重接模型
1909年 janssens 提出交叉型假说:每 次交叉表明父母本的一条染色单体接触、 断裂和重接,形成一个新的组合,其他 两条染色单体仍保持完整状态; 1937年Darlington提出重组的断裂和重 接模型:减数分裂中同源染色体相互分 离就像将绳子的两股分开一样会产生扭 曲,为消除张力,两姐妹染色体单体在 对应位点发生断链,然后非姐妹染色体 单体的“断头”相互重接,产生重组。
异源双链DNA区应为:A C A G T T G T A A 异源的DNA是不稳定的 不同异常分离比的形成,见书p213。
G G A C A
野生型g+
丢失
G
T A 突变型g-