细胞生物学教程8
合集下载
细胞生物学教程8
• 第二信使:起信号转换和放大的作用,如 cAMP、cGMP、IP3、DAG、Ca2+。
三、受体(receptor)
• 能够识别和选择性结合某种配体(信号分 子),多为糖蛋白,分子上具有配体结合 区域和产生效应的区域。
• 特点:①特异性;②饱和性;③高度的亲 和力。
• 类别:细胞内受体和细胞表面受体。
第八章 细胞通信
CELL COMMUNICATION
FUNCTIONS OF CELL COMMUNICATION
Gene transcription Cell proliferation Cell survival Cell death Cell differentiation Cell function Cell motility Immune responses
• 作用:通过磷酸化调节蛋白质的活性。
Protein kinases
五、胞间通信的主要类型
• (一)细胞间隙连接 • 连接子中央为直径1.5nm的亲水性孔道,允
许小分子如Ca2+、cAMP通过。 • 作用:协同相邻细胞对外界信号的反应,
如可兴奋细胞的电耦联现象(电紧张突触)。
connexon
(二)膜表面分子接触通讯
Three conformation of the acetylcholine receptor
Ion-channel linked receptors in neurotransmission
二、G蛋白耦联型受体
• 7次跨膜蛋白,胞外结构域识别信号分子,胞内结 构域与G蛋白耦联,调节相关酶活性,在胞内产生 第二信使。
• 旁分泌:信号分子通过扩散作用于邻近的细胞。 包括:①各类细胞因子;②气体信号分子。
三、受体(receptor)
• 能够识别和选择性结合某种配体(信号分 子),多为糖蛋白,分子上具有配体结合 区域和产生效应的区域。
• 特点:①特异性;②饱和性;③高度的亲 和力。
• 类别:细胞内受体和细胞表面受体。
第八章 细胞通信
CELL COMMUNICATION
FUNCTIONS OF CELL COMMUNICATION
Gene transcription Cell proliferation Cell survival Cell death Cell differentiation Cell function Cell motility Immune responses
• 作用:通过磷酸化调节蛋白质的活性。
Protein kinases
五、胞间通信的主要类型
• (一)细胞间隙连接 • 连接子中央为直径1.5nm的亲水性孔道,允
许小分子如Ca2+、cAMP通过。 • 作用:协同相邻细胞对外界信号的反应,
如可兴奋细胞的电耦联现象(电紧张突触)。
connexon
(二)膜表面分子接触通讯
Three conformation of the acetylcholine receptor
Ion-channel linked receptors in neurotransmission
二、G蛋白耦联型受体
• 7次跨膜蛋白,胞外结构域识别信号分子,胞内结 构域与G蛋白耦联,调节相关酶活性,在胞内产生 第二信使。
• 旁分泌:信号分子通过扩散作用于邻近的细胞。 包括:①各类细胞因子;②气体信号分子。
细胞生物学件8_内膜系统-内质网

• 真核细胞胞质核糖体80S,原核细胞的 70S,线粒体55S。
二、核糖体的化学组成
• 主 要 成 分 蛋 白 质 和 RNA , 蛋 白 质 占 40~50%, RNA占50~60%。
• 组成核糖体的RNA特称为核糖体RNA(r RNA ) , 占 细 胞 中 RNA 总 量 的 80% 以 上 。
一.内质网的形态结构
• 内质网膜约占细胞总膜面积的一半,是真 核细胞中最多的膜。
• 内质网是内膜构成的封闭的网状管道系统。 具有高度的多型性。
• 可分为粗面型内质网(RER)和光面型内 质网(SER)两类。
• RER呈扁平囊状,排列整齐,有核糖体附着。 肌 肉 细 胞 中 的 肌 质 网 是 一 种 特 化 的 RER , 可 贮存Ca2+,引起肌肉收缩。
glycosylation):与Ser和Thr的OH连 接,连接的糖为半乳糖或N乙酰半乳糖胺。 如胶原、血型糖蛋白等 • 2.N-连接的糖基化(N-linked glycosylation):与天冬酰胺残基的 NH2连接,连接的糖为N-乙酰葡糖胺。
• 内质网上进行的为N-连接的糖基化。图 7-9
• 糖的供体为核苷糖(nucleotide sugar), 如CMP-唾液酸、GDP-甘露糖、UDPN-乙酰葡糖胺等。糖分子首先被糖基转 移酶转移到膜上的磷酸长醇(dolichol phosphate)分子上,装配成寡糖链。 再被寡糖转移酶转到新合成肽链特定序 列(Asn-X-Ser或Asn-X-Thr)的天 冬酰胺残基上。
然后进行扩增。
第三节 核糖体(Ribosome)
• 核糖体在各类细胞中普遍存在,是一种 无膜包被的颗粒状结构,具有很强的嗜 碱性。
• 功能:蛋白合成
一、核糖体 的基本结 构与类型
二、核糖体的化学组成
• 主 要 成 分 蛋 白 质 和 RNA , 蛋 白 质 占 40~50%, RNA占50~60%。
• 组成核糖体的RNA特称为核糖体RNA(r RNA ) , 占 细 胞 中 RNA 总 量 的 80% 以 上 。
一.内质网的形态结构
• 内质网膜约占细胞总膜面积的一半,是真 核细胞中最多的膜。
• 内质网是内膜构成的封闭的网状管道系统。 具有高度的多型性。
• 可分为粗面型内质网(RER)和光面型内 质网(SER)两类。
• RER呈扁平囊状,排列整齐,有核糖体附着。 肌 肉 细 胞 中 的 肌 质 网 是 一 种 特 化 的 RER , 可 贮存Ca2+,引起肌肉收缩。
glycosylation):与Ser和Thr的OH连 接,连接的糖为半乳糖或N乙酰半乳糖胺。 如胶原、血型糖蛋白等 • 2.N-连接的糖基化(N-linked glycosylation):与天冬酰胺残基的 NH2连接,连接的糖为N-乙酰葡糖胺。
• 内质网上进行的为N-连接的糖基化。图 7-9
• 糖的供体为核苷糖(nucleotide sugar), 如CMP-唾液酸、GDP-甘露糖、UDPN-乙酰葡糖胺等。糖分子首先被糖基转 移酶转移到膜上的磷酸长醇(dolichol phosphate)分子上,装配成寡糖链。 再被寡糖转移酶转到新合成肽链特定序 列(Asn-X-Ser或Asn-X-Thr)的天 冬酰胺残基上。
然后进行扩增。
第三节 核糖体(Ribosome)
• 核糖体在各类细胞中普遍存在,是一种 无膜包被的颗粒状结构,具有很强的嗜 碱性。
• 功能:蛋白合成
一、核糖体 的基本结 构与类型
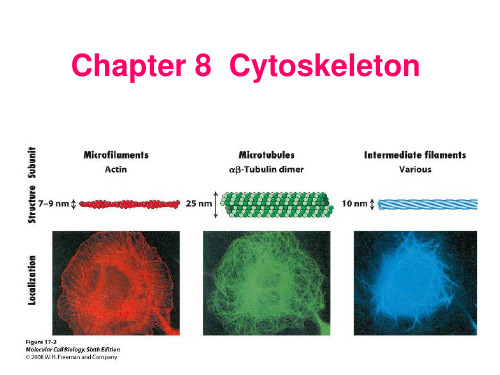
《细胞生物学》——细胞8章 细胞骨架1
2. 下列哪种蛋白可以帮助微丝快速组装()
A 球状肌动蛋白 B 纤维状肌动蛋白
G蛋 C白 D Arp2/3
提交
3. 微丝的组装不包括下列哪个阶段() A. 成核反应 B. 纤维的延长 C. 踏车行为 D. 解离阶段
单选题 10分
3. 微丝的组装不包括下列哪个阶段()
A 成核反应 B 纤维的延长 C 踏车行为 D 解离阶段
即正极与负极之别
(2)肌动蛋白单体组装成微丝的过程
① 缓慢成核期:肌动蛋白单体与起始复合物结合→形成寡聚体(至少2-3
个单体)。
包括2种肌动蛋白相关蛋白(Arp2/3)和5种
其它蛋白。
② 快速延长期:肌动蛋白单体具有ATP酶活性,可利用水解ATP释放的能
量来快速组装单体。当微丝的组装速度快于肌动蛋白水解ATP的速度时,在
提交
本章主要内容
微丝与细胞运动 微管及其功能 中间丝
第一节 微丝与细胞运动
微丝(microfilament, MF) 肌动蛋白丝(actin filament) 纤维状肌动蛋白
(fibrous, F-actin)
由肌动蛋白单体组装而成的 直径为7 nm的纤维状结构 存在于所有真核细胞中 微丝的组装/去组装 微丝结合蛋白
(MF binding protein)
一、微丝的组成及其组装 二、微丝网络结构的调节与细胞运动
一、微丝的组成及其组装
(一)微丝的结构与成分 (二)微丝的组装及其动力学特性 (三)影响微丝组装的特异性药物
(一)微丝的结构与成分
• 微丝的主要结构成分:肌动蛋白(actin)。
• 肌动蛋白在细胞内有2种存在形式:①球状肌动蛋白(G-actin): 肌动蛋白单体;②纤维状肌动蛋白(F-actin):由多个单体组装 而成。
A 球状肌动蛋白 B 纤维状肌动蛋白
G蛋 C白 D Arp2/3
提交
3. 微丝的组装不包括下列哪个阶段() A. 成核反应 B. 纤维的延长 C. 踏车行为 D. 解离阶段
单选题 10分
3. 微丝的组装不包括下列哪个阶段()
A 成核反应 B 纤维的延长 C 踏车行为 D 解离阶段
即正极与负极之别
(2)肌动蛋白单体组装成微丝的过程
① 缓慢成核期:肌动蛋白单体与起始复合物结合→形成寡聚体(至少2-3
个单体)。
包括2种肌动蛋白相关蛋白(Arp2/3)和5种
其它蛋白。
② 快速延长期:肌动蛋白单体具有ATP酶活性,可利用水解ATP释放的能
量来快速组装单体。当微丝的组装速度快于肌动蛋白水解ATP的速度时,在
提交
本章主要内容
微丝与细胞运动 微管及其功能 中间丝
第一节 微丝与细胞运动
微丝(microfilament, MF) 肌动蛋白丝(actin filament) 纤维状肌动蛋白
(fibrous, F-actin)
由肌动蛋白单体组装而成的 直径为7 nm的纤维状结构 存在于所有真核细胞中 微丝的组装/去组装 微丝结合蛋白
(MF binding protein)
一、微丝的组成及其组装 二、微丝网络结构的调节与细胞运动
一、微丝的组成及其组装
(一)微丝的结构与成分 (二)微丝的组装及其动力学特性 (三)影响微丝组装的特异性药物
(一)微丝的结构与成分
• 微丝的主要结构成分:肌动蛋白(actin)。
• 肌动蛋白在细胞内有2种存在形式:①球状肌动蛋白(G-actin): 肌动蛋白单体;②纤维状肌动蛋白(F-actin):由多个单体组装 而成。
细胞生物学第八章细胞信号转导

信号蛋白:
① ② ③ ④ ⑤ ⑥ ⑦ 转承蛋白:负责将信息传给信号链的下一组分。 信使蛋白:携带信息从一部分传递到另一部分。 接头蛋白:起连接信号蛋白的作用。 放大和转导蛋白:通常由酶或离子通道蛋白组成,介导产生级联反 应。 传感蛋白:负责信号不同形式的转换。 分歧蛋白:将信号从一条途径传播到另外途径。 整合蛋白:从 2 条或多条信号途径接受信号,并在向下传递之前进 行整合。
2、受体
受体:受体是一种能够识别和选择性结合某种配体(信号分子)的大 分子,绝大多数都是蛋白质且多为但蛋白,少数受体是糖脂,有的受 体是糖蛋白和糖脂组成的复合物。 (1)根据靶细胞上受体存在的部位,可将手提取分为 细胞内受体:位于细胞质基质或核基质中,主要是别和结合小 的脂溶性信号分子。
c、间隙连接通透性的调节:
意义:间隙连接对小分子的通透能力具有底物选择性。因此通过掌握调节间 隙连接通透性的途径有助于对信号分子的传递调控。 特性: 1、电荷选择性: 间隙连接的通透能力与底物所带电荷有关。
2 、组织特异性: 由不同连接蛋白所构成的连接子,在导电率、通透性 和可调控方面是不同的。由不同连接蛋白组成的异聚体连接子一般具有通透 功能,但在有些情况下却没有通透功能。如:Cx43与Cx40连接蛋白形成间隙 连接时,连接子没有通透功能。
二、信号转导系统及其特性
(一)信号转导系统的基本组成与信号蛋白 信号转导系统: 1、不同形式的胞外的信号刺激首先被细胞表面特异性受体所识别。 2、胞外信息通过适当的分子开关机制实现信号的跨膜转导,产生 细胞内第二信使或活化的信号蛋白。 3、信号放大:信号传递至胞内效应器蛋白,引发细胞内信号放大 的级联反应,使信号逐级放大。 4、启动反馈机制从而终止或降低细胞反应。
细胞生物学教学完整ppt课件

.
在非细胞系统中蛋白质的翻译过程与SRP、 DP和微粒体的关系(表8-1)
.
分泌性蛋白的合成与跨越内质网膜的共翻 译转运图解(图8-3)
共翻译转运(cotranslational translocation):分泌 蛋白向rER腔内的转运是同蛋白质翻译过程偶联进行的,这 种蛋白在信号肽引导下边翻译边跨膜转运的过程称为共翻译 转运。
多次跨膜蛋白:含有多个SA和多个STA的肽链将成为多次跨 膜蛋白。
跨内质网膜肽段的取向:一般而言,带正电荷氨基酸残基多
的一端,或带正电荷氨基酸残基多的一侧,朝向细胞质基质
一侧(外侧)。
.
线粒体、叶绿体和过氧化物酶体的蛋白质的信 号序列特称为导肽(leader peptide),其基本 的特征是蛋白质在细胞质基质中的游离核糖体 上合成以后再转移到这些细胞器中,因此称这 种翻译后再转运的方式为翻译后转运(posttranslational translocation)。这种转运方式 在蛋白质跨膜过程中不仅需要消耗ATP使多肽 去折叠,而且还需要跨膜后由分子伴侣帮助蛋 白质再正确折叠形成有功能的蛋白。
继信号假说提出与确证后,人们又发现一系列
蛋白质分选信号序列,统称信号序列(signal
sequence),而且有些信号序列还可形成三维
结构的信号斑(signal patch),指导蛋白的靶
向转运和定位。
.
指导蛋白质从细胞基质转运到细胞器的靶 向序列的主要特征(表8-2)
.
二、蛋白质分选转运的基本途径与类型
第八章 蛋白质分选与膜泡运输
第一节 细胞内蛋白质的分选 第二节 细胞内膜泡运输
.
第一节 细胞内蛋白质的分选
真核细胞中绝大多数蛋白质都是由核基因编码,起始合 成均发生在游离核糖体上,然后或在细胞质基质中游离核糖 体上完成翻译过程,或在粗面内质网膜结合核糖体上完成合 成。之后蛋白质各就各位并组装成结构与功能的复合体,参 与实现细胞的各种生命活动。这一过程称蛋白质分选 (protein sorting)或蛋白质寻靶(protein targeting)。 蛋白质分选主要依靠蛋白质自身信号序列,从蛋白质起始合 成部位转运到其功能发挥部位的过程。
在非细胞系统中蛋白质的翻译过程与SRP、 DP和微粒体的关系(表8-1)
.
分泌性蛋白的合成与跨越内质网膜的共翻 译转运图解(图8-3)
共翻译转运(cotranslational translocation):分泌 蛋白向rER腔内的转运是同蛋白质翻译过程偶联进行的,这 种蛋白在信号肽引导下边翻译边跨膜转运的过程称为共翻译 转运。
多次跨膜蛋白:含有多个SA和多个STA的肽链将成为多次跨 膜蛋白。
跨内质网膜肽段的取向:一般而言,带正电荷氨基酸残基多
的一端,或带正电荷氨基酸残基多的一侧,朝向细胞质基质
一侧(外侧)。
.
线粒体、叶绿体和过氧化物酶体的蛋白质的信 号序列特称为导肽(leader peptide),其基本 的特征是蛋白质在细胞质基质中的游离核糖体 上合成以后再转移到这些细胞器中,因此称这 种翻译后再转运的方式为翻译后转运(posttranslational translocation)。这种转运方式 在蛋白质跨膜过程中不仅需要消耗ATP使多肽 去折叠,而且还需要跨膜后由分子伴侣帮助蛋 白质再正确折叠形成有功能的蛋白。
继信号假说提出与确证后,人们又发现一系列
蛋白质分选信号序列,统称信号序列(signal
sequence),而且有些信号序列还可形成三维
结构的信号斑(signal patch),指导蛋白的靶
向转运和定位。
.
指导蛋白质从细胞基质转运到细胞器的靶 向序列的主要特征(表8-2)
.
二、蛋白质分选转运的基本途径与类型
第八章 蛋白质分选与膜泡运输
第一节 细胞内蛋白质的分选 第二节 细胞内膜泡运输
.
第一节 细胞内蛋白质的分选
真核细胞中绝大多数蛋白质都是由核基因编码,起始合 成均发生在游离核糖体上,然后或在细胞质基质中游离核糖 体上完成翻译过程,或在粗面内质网膜结合核糖体上完成合 成。之后蛋白质各就各位并组装成结构与功能的复合体,参 与实现细胞的各种生命活动。这一过程称蛋白质分选 (protein sorting)或蛋白质寻靶(protein targeting)。 蛋白质分选主要依靠蛋白质自身信号序列,从蛋白质起始合 成部位转运到其功能发挥部位的过程。
细胞生物学 第八章细胞骨架

二、微丝网络结构的调节与细胞运动
(二)细胞皮层 • 细胞皮层:紧贴细胞质膜下
富含微丝的区域,由微丝结 合蛋白交联成凝胶态的三维 网络结构。 • 其功能有: s 微丝与细胞质膜上的蛋白质 连接,从而限制膜蛋白的流 动性; s 为细胞质膜提供强度和韧性, 从而维持细胞的形状; s 细胞的多种运动如胞质环流、 吞噬等都与微丝的组装/去 组装转化有关。
由质膜及质膜下的轴心微丝束 构成。 • 微丝束的微丝呈同向平行排列, 即正极指向微绒毛顶端,负极 指向细胞内。 • 微绒毛的作用在于增加细胞的 吸收表面积,而其内部的轴心 微丝束起着支撑作用。
二、微丝动态结构的调节与细胞运动
(六)胞质分裂环 • 在有丝分裂末期,两个即
将分离的子细胞之间产生 一个对细胞质起收缩作用 的胞质分裂环(收缩 环)——由大量平行排列、 但极性相反的微丝组成。 • 收缩环的动力来源于肌球 蛋白在极性相反的微丝之 间的滑动。 • 随着收缩环的收缩,两个 子细胞被分开。
• 微管(microtubule,MT): 指真核细胞中由微管蛋白组成 的、中空的管状骨架纤维,其 外径为24(25)nm,内径为 15nm。
• 细胞中的微管多数是暂时性的 结构:如间期细胞内的微管, 分裂期细胞的纺锤体微管等;
• 也有一些微管形成相对稳定的 “永久性”结构:如纤毛或鞭 毛中的轴丝微管、神经元突起 内的微管束。
§2 微管及其功能
一、微管的结构组分与极性
• 微管组装的基本结构单位是由 α-微管蛋白和β-微管蛋白结合 而成的异二聚体——α/β-微管 蛋白二聚体,是细胞内游离态 微管蛋白的主要存在形式。
• α-微管蛋白和β-微管蛋白上各有 一个GTP结合位点:
s α-微管蛋白:不可交换位点
细胞生物学第8章-细胞骨架
动力蛋白驱动蛋白驱动蛋白和动力蛋白的头部都具有atp水解酶的活性酶解atp的能量可供这两个蛋白质的头部做一次构象变换完成一套与微管的结合解离再结合的动作从而使蛋白质沿着微管移动
细胞骨架
本章内容提要
细胞质骨架 微管 微丝 中间丝
膜骨架 核骨架
细胞骨架(Cytoskeleton)指存在于真核细胞中的 蛋白纤维网架体系。
轴丝微管的排列方式主要有3中模式:
①9+2型:轴丝的外围是9组二联体微管,中 间是2根由中央鞘所包围的中央微管。
②9+0型:外围与9+2型相同,有9组二联体 微管,但缺乏中央微管。
③9+4型:罕见,轴丝中央含有4根单体微管。
9+0型一般为不动纤毛,9+2型大多为动纤毛。
纤毛和鞭毛运动的机制--“滑动微管模型”。 由于二联管之间的动力蛋白臂的动力蛋白介导了 两条微管的互相滑动,才使纤毛发生运动。
细胞骨架是高度动态的细胞结构体系,不仅是重要 的机械支撑、决定了细胞器和大分子空间定位,而 且参与了几乎所有形式的细胞运动。
细胞骨架直接负责细胞的大幅度运动,例如,细胞 沿物体表面爬行、肌肉细胞的收缩以及胚胎发育时 细胞形状的改变。
没有细胞骨架,伤口无法愈合,肌肉毫无用处, 精子也不能触及卵子。
Microtubules
Microfilamemts
Intermediate filaments
一、微管
微管几乎存在于所有的真核细胞内,是由微 管蛋白(tubulin)装配而成的中空的管状 结构。
它可以迅速地在细胞内某个位置去组装,并 在另一个位置再组装。
1.结构:
微管由微管蛋白亚基组装而成,每个亚基 是由α微管蛋白和β微管蛋白,通过非共价键 结合成的异二聚体,是微管装配的基本单位。
细胞骨架
本章内容提要
细胞质骨架 微管 微丝 中间丝
膜骨架 核骨架
细胞骨架(Cytoskeleton)指存在于真核细胞中的 蛋白纤维网架体系。
轴丝微管的排列方式主要有3中模式:
①9+2型:轴丝的外围是9组二联体微管,中 间是2根由中央鞘所包围的中央微管。
②9+0型:外围与9+2型相同,有9组二联体 微管,但缺乏中央微管。
③9+4型:罕见,轴丝中央含有4根单体微管。
9+0型一般为不动纤毛,9+2型大多为动纤毛。
纤毛和鞭毛运动的机制--“滑动微管模型”。 由于二联管之间的动力蛋白臂的动力蛋白介导了 两条微管的互相滑动,才使纤毛发生运动。
细胞骨架是高度动态的细胞结构体系,不仅是重要 的机械支撑、决定了细胞器和大分子空间定位,而 且参与了几乎所有形式的细胞运动。
细胞骨架直接负责细胞的大幅度运动,例如,细胞 沿物体表面爬行、肌肉细胞的收缩以及胚胎发育时 细胞形状的改变。
没有细胞骨架,伤口无法愈合,肌肉毫无用处, 精子也不能触及卵子。
Microtubules
Microfilamemts
Intermediate filaments
一、微管
微管几乎存在于所有的真核细胞内,是由微 管蛋白(tubulin)装配而成的中空的管状 结构。
它可以迅速地在细胞内某个位置去组装,并 在另一个位置再组装。
1.结构:
微管由微管蛋白亚基组装而成,每个亚基 是由α微管蛋白和β微管蛋白,通过非共价键 结合成的异二聚体,是微管装配的基本单位。
细胞生物学-第8章-蛋白质分选与膜泡运输
二、COPII包被膜泡的装配与运输
• 负责从内质网高尔基体的物质运输; • COPII包被蛋白由5种蛋白亚基组成:Sar1、 Sec12、
Sec23/Sec24、 Sec13/Sec31、Sec16 • 包被蛋白的装配是受控的; • COPII包被膜泡具有对转运物质的选择性并使之浓缩。
COPII蛋白能识别并结合跨膜内质网蛋白胞质面一端的信 号序列(Asp-X-Glu) 内质网腔面的受体能与ER腔中的可溶性蛋白(如分泌蛋白 )结合。
6.组成型分泌
7.调节型分泌
8.分选到溶酶体
9.胞吞途径
蛋白质的分泌与胞吞途径概观
膜泡运输
• 在供体膜上如何形成膜泡? • 如何运输膜? • 膜泡如何锚定靶膜并与之融合?
在供体膜上如何形成膜泡?
电子显微镜下观察,发现膜泡表面有一层蛋白质
包被蛋白的功能:
1、形成膜泡:使供体膜弯曲 2、选择膜泡成分:被运输的货物 3、靶膜融合:包含与靶膜融合相关蛋白
• 蛋白质分选的运输方式: 1.门控运输;2.跨膜运输;3.膜泡运输;4.细胞 质基质中的蛋白质运输。
• 蛋白质分选的信号机制: 信号肽;信号识别颗粒;信号识别颗粒的受体 ;ER膜上的易位子等。
• 3种类型的有被小泡: 1. 网格蛋白/接头蛋白小泡 ;2. COPI小泡;3. COPII小泡
anchor sequence, STA)
• 内在信号锚定序列(internal signal anchor sequence, SA)
膜整合蛋白 一次跨膜
膜整合蛋白 两次跨膜
多肽跨膜次数的确定
内质网膜整合蛋白的拓扑学类型
膜整合蛋白 方向性
蛋白分选机制
1、蛋白质自身信号序列 2、靶细胞器上包含特定的信号识别装置
细胞生物学第八章
号分子影响其它细胞。
3.细胞间形成间隙连接,使细胞质相互沟通—动物
细胞间隙连接、植物细胞胞间连丝通过交换小分
子实现代谢偶联或电偶联的通讯方式。
细胞分泌化学信号的作用方式
(1)内分泌(endocrine)
内分泌腺 激素 血液循环 靶器官(靶细胞)
(2)旁分泌(paracrine) 信号细胞 局部化学介质 细胞外液 临近靶细胞
白磷酸化,通过蛋白磷酸酶使靶蛋白去磷酸化,从而调节 蛋白质的活性。
细胞内信号传导过程中两类分子开关蛋白
蛋白激酶 蛋白磷酸酯酶
二、信号转导系统及其特性
(一)信号转到系统的基本组成与信号蛋白 通过细胞表面受体介导的信号途径由下列4个步骤组成: 1. 信号刺激首先被细胞表面特异性受体所识别; 特异性是识别反应的主要特征,这源于信号分子与互补受 体上的结合位点相适应。 2. 胞外信号(第一信使)通过适当的分子开关机制实现信号 的跨膜转导,产生胞内第二信使或活化的信号蛋白; 绝大多数被激活的细胞表面受体是通过小分子第二信使和 细胞内信号蛋白网络传播信号的。
胞内信号分子
靶蛋白
新陈代谢酶 基因调控蛋白 细胞支架蛋白
从细胞表面到细胞核的信号途径是由细胞内多种不同的信 号蛋白组成的信号传递链,这条信号蛋白链负责实现上述4个 号传递的主要步骤,除细胞表面受体之外还包括如下各类蛋 白质: ① 转承蛋白:负责简单地将信息传给信号链的下一个组分; ② 信使蛋白:携带信号从一部分传递到另一部分; ③ 接头蛋白:连接信号蛋白; ④ 放大和转导蛋白:通常由酶或离子通道蛋白组成介导产生 信号级联反应; ⑤ 传感蛋白:负责信号不同形式的转换; ⑥ 分歧蛋白:将信号从一条途径传播到另外途径; ⑦ 整合蛋白:从2条信号途径接收信号,并在向下传递之前进 行整合; ⑧ 潜在基因调控蛋白:这类蛋白在细胞表面被活化受体激活, 然后迁移到细胞核刺激基因转录。
3.细胞间形成间隙连接,使细胞质相互沟通—动物
细胞间隙连接、植物细胞胞间连丝通过交换小分
子实现代谢偶联或电偶联的通讯方式。
细胞分泌化学信号的作用方式
(1)内分泌(endocrine)
内分泌腺 激素 血液循环 靶器官(靶细胞)
(2)旁分泌(paracrine) 信号细胞 局部化学介质 细胞外液 临近靶细胞
白磷酸化,通过蛋白磷酸酶使靶蛋白去磷酸化,从而调节 蛋白质的活性。
细胞内信号传导过程中两类分子开关蛋白
蛋白激酶 蛋白磷酸酯酶
二、信号转导系统及其特性
(一)信号转到系统的基本组成与信号蛋白 通过细胞表面受体介导的信号途径由下列4个步骤组成: 1. 信号刺激首先被细胞表面特异性受体所识别; 特异性是识别反应的主要特征,这源于信号分子与互补受 体上的结合位点相适应。 2. 胞外信号(第一信使)通过适当的分子开关机制实现信号 的跨膜转导,产生胞内第二信使或活化的信号蛋白; 绝大多数被激活的细胞表面受体是通过小分子第二信使和 细胞内信号蛋白网络传播信号的。
胞内信号分子
靶蛋白
新陈代谢酶 基因调控蛋白 细胞支架蛋白
从细胞表面到细胞核的信号途径是由细胞内多种不同的信 号蛋白组成的信号传递链,这条信号蛋白链负责实现上述4个 号传递的主要步骤,除细胞表面受体之外还包括如下各类蛋 白质: ① 转承蛋白:负责简单地将信息传给信号链的下一个组分; ② 信使蛋白:携带信号从一部分传递到另一部分; ③ 接头蛋白:连接信号蛋白; ④ 放大和转导蛋白:通常由酶或离子通道蛋白组成介导产生 信号级联反应; ⑤ 传感蛋白:负责信号不同形式的转换; ⑥ 分歧蛋白:将信号从一条途径传播到另外途径; ⑦ 整合蛋白:从2条信号途径接收信号,并在向下传递之前进 行整合; ⑧ 潜在基因调控蛋白:这类蛋白在细胞表面被活化受体激活, 然后迁移到细胞核刺激基因转录。
细胞生物学-8细胞核与染色体-01
细 胞 核 与 染 色 体
学习要求
1、掌握细胞核(包括核膜与核孔复合体、染 色质与染色体、核仁与核基质) 2、理解染色质与染色体间的互变和染色质结 构与基因转录间的关系 3、了点为细胞核各部分的结构与功能, 及其相关的生命过程 2.本章难点为核孔复合体的结构与功能、染 色质和染色体的结构、巨大染色体、染色 质结构与基因转录、核基质与核体的知识
细胞核的形态结构
细胞核(nucleus)与染色体(chromosome)
●细胞核是真核细胞内最大、最重要的细胞器,
是细胞遗传与代谢的调控信息中心
●细胞核的结构组成: 核被膜(nuclear envelope)与核孔复合体(NPC) 染色质(chromatin) 染色体 (chromosome )
组蛋白的功能
◆核小体组蛋白(nucleosomal histone),
包括H2A、H2B、H3和H4, 作用是与DNA
组装成核小体
◆H1不参加核小体的组建, 在构成核小体时
起连接作用,并赋予染色质以极性。
组蛋白的化学修饰
虽然组蛋白分子的氨基酸序列是高度
保守的,也会进行一些化学修饰,如
酰基化、甲基化、磷酸化,以及ADP核
核孔复合物的功能
核孔运输特点
◆被动运输 ◆主动运输 ●信号引导
●双向性
通过核孔复合体物质运输的功能示意图(引自B.Talcott等,1999)
核孔的运输作用
核孔的被动运输
• 1965年,Ccarl Feldherr 将各种不同大小 的金颗粒注射到变形虫的细胞质中,然后 检查这些颗粒进入细胞核的情况,发现∶ 小的金颗粒很快进入了细胞核,但体积越 大进入的速度就越慢,大于10nm的金颗粒 就进不了细胞核,由此推测∶核孔可作为 水性的运输通道,允许小分子的物质自由 扩散出入细胞核
学习要求
1、掌握细胞核(包括核膜与核孔复合体、染 色质与染色体、核仁与核基质) 2、理解染色质与染色体间的互变和染色质结 构与基因转录间的关系 3、了点为细胞核各部分的结构与功能, 及其相关的生命过程 2.本章难点为核孔复合体的结构与功能、染 色质和染色体的结构、巨大染色体、染色 质结构与基因转录、核基质与核体的知识
细胞核的形态结构
细胞核(nucleus)与染色体(chromosome)
●细胞核是真核细胞内最大、最重要的细胞器,
是细胞遗传与代谢的调控信息中心
●细胞核的结构组成: 核被膜(nuclear envelope)与核孔复合体(NPC) 染色质(chromatin) 染色体 (chromosome )
组蛋白的功能
◆核小体组蛋白(nucleosomal histone),
包括H2A、H2B、H3和H4, 作用是与DNA
组装成核小体
◆H1不参加核小体的组建, 在构成核小体时
起连接作用,并赋予染色质以极性。
组蛋白的化学修饰
虽然组蛋白分子的氨基酸序列是高度
保守的,也会进行一些化学修饰,如
酰基化、甲基化、磷酸化,以及ADP核
核孔复合物的功能
核孔运输特点
◆被动运输 ◆主动运输 ●信号引导
●双向性
通过核孔复合体物质运输的功能示意图(引自B.Talcott等,1999)
核孔的运输作用
核孔的被动运输
• 1965年,Ccarl Feldherr 将各种不同大小 的金颗粒注射到变形虫的细胞质中,然后 检查这些颗粒进入细胞核的情况,发现∶ 小的金颗粒很快进入了细胞核,但体积越 大进入的速度就越慢,大于10nm的金颗粒 就进不了细胞核,由此推测∶核孔可作为 水性的运输通道,允许小分子的物质自由 扩散出入细胞核
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
• 不同细胞对cAMP信号途径的反应:
– 在肌肉细胞,1秒钟内可启动糖原降解为葡糖1‐ 磷酸,而抑制糖原合成。 – 在某些分泌细胞,需要几个小时, 激活的PKA 进入细胞核,将CRE (cAMP response element ) 结合蛋白磷酸化,调节相关基因的表达。CRE 是DNA上的调节区域。
• ③ 腺苷酸环化酶: 跨 膜 12 次 。 在 Mg2+ 或 Mn2+ 的 存 在 下 , 催 化 ATP 生 成 cAMP 。
Adenylate cyclase
• ④蛋白激酶A(Protein Kinase A,PKA): 由两个催化亚基和两个调节亚基组成。
•cAMP与调节亚基结合,使调节亚基和催化亚基解离,释放出催化亚基, 激活蛋白激酶A的活性。
G蛋白偶联型受体的钝化
• 细胞通过受体钝化适应持续刺激:
– ①受体失活(receptor inactivation); – ②受体隐蔽(receptor sequestration); – ③受体下行调节(receptor down‐regulation)。
(四)小G蛋白(Small G Protein)
第一节 基本概念
一、几个容易混淆的概念
• 细胞信号发放(cell signaling) • 细胞通讯(cell communication) • 细胞识别(cell recognition) • 信号转导(signal transduction)
二、细胞信号分子
• 1.化学感受器中的G蛋白 • 气味分子与受体结合,激活腺苷酸环化酶, 产生cAMP,开启cAMP门控阳离子通道,引 起钠离子内流,膜去极化,产生神经冲动, 形成嗅觉或味觉。
2004年Axel和Buck因发现气味受体和化学感 受器系统的组成而获诺贝尔生理与医学奖。
Richard Axel
Linda B. Buck
•
第二信使:起信号转换和放大的作用,如 cAMP、cGMP、IP3、DAG、Ca2+。
三、受体(receptor)
• 能够识别和选择性结合某种配体(信号分 子),多为糖蛋白,分子上具有配体结合 区域和产生效应的区域。 • 特点:①特异性;②饱和性;③高度的亲 和力。 • 类别:细胞内受体和细胞表面受体。
2.视觉感受器中的G蛋白
• 黑暗时视杆细胞中cGMP浓度较高,cGMP门 控钠通道开放,钠内流,膜去极化,突触 持续向次级神经元释放递质。 • 有光时视紫红质作用于G蛋白,激活磷酸二 酯酶,cGMP浓度下降,形成光感受。
cGMP in Photoreception
视紫红质为7次跨膜蛋白,由视蛋白和视黄醛组成。其信号途径为: 光信号→Rh激活→Gt活化→cGMP磷酸二酯酶激活→胞内cGMP减少→Na+离子 通道关闭→离子浓度下降→膜超极化→神经递质释放减少→视觉反应。
GTP-binding regulatory protein
• cAMP信号途径可表示为:
– 激素→受体→G蛋白→腺苷酸环化酶→cAMP→ 蛋白激酶A→基因调控蛋白→基因转录。
• 霍乱毒素能催化ADP核糖基共价结合到Gs的 α亚基上,使α亚基持续活化,导致患者细 胞内Na+和水持续外流,产生严重腹泻而脱 水。
Inositol phospholipid signaling
Mimicked by ionomycin
PIP2 Hydrolysis
IP3开启胞内IP3门控钙通道: Ca2+浓度升高,激活钙调蛋 白,CaM将靶蛋白(如: CaM-Kinase)活化。 DAG激活蛋白激酶C:PKC位 于细胞质,Ca2+浓度升高时 PKC转位到质膜内表面,被 DAG活化。
• ⑤环腺苷酸磷酸二酯酶 (PDE) :降解 cAMP 生成5’‐AMP,终止信号。
Degredation of cAMP
• Gs调节模型: • 激素与Rs结合,Rs构象改变,与Gs结合,Gs 的α亚基排斥GDP,结合GTP而活化,Gs解离 出α和βγ。
– α亚基活化腺苷酸环化酶,将ATP转化为cAMP。 – βγ亚基复合物可直接激活某些胞内靶分子。
(三)受体酪氨酸磷酯酶
• 使特异的胞内信号蛋白去磷酸化。 • 控制磷酸酪氨酸残基的寿命,使静止细胞 具有较低的磷酸酪氨酸残基水平。 • 与酪氨酸激酶协同工作,如参与细胞周期 调控。
• 三聚体GTP结合蛋白,即,G蛋白: • 组成:αβγ,α和γ属脂锚定蛋白。 • 作用:分子开关。 α 亚基结合 GDP 失活,结 合GTP活化。 α也是GTP酶,催化结合的ATP 水解,恢复无活性状态,其GTP酶活性可被 GAP增强。
(一)cAMP信号途径
• 通过调节胞内 cAMP 的浓度,将细胞外信 号转变为细胞内信号。 • 主要组分: • ①受体:Rs、Ri • ②G蛋白:Gs、Gi
• 特点: • ①单次跨膜蛋白; • ②接受配体后发生二 聚化和自磷酸化,起 动下游信号转导。
(一)受体酪氨酸激酶 • 1、酪氨酸激酶 • ①胞质酪氨酸激酶:如Src、JAK; • ②核内酪氨酸激酶 :如:Abl、Wee; • ③受体酪氨酸激酶:如EGF受体。相关信号 途径: RAS 途径、 PI3K 途径、磷脂酰肌醇途 径。
第二节 膜表面受体介导的信号转导
• ①离子通道型受体; • ②G蛋白耦联型受体; • ③酶耦联的受体。 • 第一类存在于可兴奋细胞,后两类存在于 大多数细胞。
Cell surface receptors
一、离子通道型受体
• 受体本身为离子通道,即配体门通道。主要 存在于神经、肌肉等可兴奋细胞,信号分子 为神经递质。分为:
•MEK Homo sapiens MAP kinase kinase
PI3K PATHWAY
PDK is phosphoinositol dependent kinase Akt is PKB, influence cell survival
(二)受体丝氨酸/苏氨酸激酶
• 配 体 是 转 化 生 长 因 子 ‐βs 家 族 成 员 , 包 括 TGF‐β1‐5。 • 依细胞类型不同,可抑制细胞增殖、刺激 胞外基质合成、刺激骨骼的形成、通过趋 化性吸引细胞、作为胚胎发育过程中的诱 导信号等。
Receptor tyrosine kinases
Receptor tyrosine kinases
• 2、信号分子间的识别结构域 ①SH2结构域:介导信号分子与含磷酸酪氨酸 的蛋白结合。 ②SH3结构域:介导信号分子与富含脯氨酸的 蛋白结合。 ③PH结构域:与PIP2、PIP3、IP3等结合。
• 成分:短肽、蛋白质、气体分子(NO、 CO)、氨基酸、核苷酸、脂类、胆固醇衍 生物。 • 特点:①特异;②高效;③可被灭活。 • 类别:脂溶性、水溶性信号分子。
Some signal molecules
•
第一信使:水溶性信号分子(如神经递质) 不能穿过靶细胞膜,只能经膜上的信号转 换机制实现信号传递,称第一信使。
二、G蛋白耦联型受体
• 7次跨膜蛋白,胞外结构域识别信号分子,胞内结 构域与G蛋白耦联,调节相关酶活性,在胞内产生 第二信使。 • 类型:①多种神经递质、肽类激素和趋化因子受 体,②味觉、视觉和嗅觉感受器。 • 相关信号途径:cAMP途径、磷脂酰肌醇途径。
G-protein linked receptor
3、RAS信号途径
• RPTK结合信号分子活化,激活RAS; • RAS的调节因子具有GEF、GAP; • GEF需要通过接头蛋白与RPTK连接。 • 活化的RAS启动蛋白激酶的磷酸化级联反应, 激活转录因子,调节基因表达;
RPTK-Ras GEF→ Ras→ Raf (MAPKKK)→ MAPKK→ MAPK→ 细胞 核→ 转录因子→ 基因表达。
• 信号的终止: • IP3信号:去磷酸化为IP2;磷酸化为IP4 。 • Ca2+ 信号:被钙泵和 Na+‐Ca2+ 交换器抽出细 胞,或被泵回内质网。 • DAG信号:被DAG激酶磷酸化为磷脂酸;或 被DAG酯酶水解成单酯酰甘油。
Controls on Cytosolic Calcium
(三)其它G蛋白偶联型受体
– 阳离子通道,如乙酰胆碱受体; – 阴离子通道,如γ-氨基丁酸受体。
Chemical synapse
Acetylcholine receptor
Three conformation of the acetylcholine receptor
Ion-channel linked receptors in neurotransmission
• 特点:分子量小,为单体。 • 功能:与Gα类似,起分子开关的作用。 • 类型:Ras、Rho、微管蛋白β亚基。 • 调节因子:GEF、GDI、GAP等。
三、酶耦联型受体 enzyme linked receptor
• 定义:本身具有激酶活性;或者可以连接非受体 酪氨酸激酶。 • 类别:①受体酪氨酸激酶、②受体丝氨酸/苏氨酸 激酶、③受体酪氨酸磷脂酶、④ 酪氨酸激酶连接 的受体、 ⑤受体鸟苷酸环化酶、 ⑥组氨酸激酶连 接的受体(与细菌的趋化性有关)。
• Gi调节模型 • ①通过α亚基与腺苷酸环化酶结合,直接抑制酶的活性; • ②通过βγ亚基复合物与游离Gs的α亚基结合,阻断Gs的α亚 基对腺苷酸环化酶的活化。 • 百日咳毒素抑制Gi的活性。
(二)磷脂酰肌醇途径
• 又称双信使途径。 • 信号分子与G蛋白耦联受体结合,激活质膜 上的磷脂酶 C ( PLC‐β ),使 4 , 5‐ 二磷酸磷 脂酰肌醇(PIP2)水解成IP3和DAG。
• 细胞对信号的反应不仅取决于其受体的特 异性,而且与细胞的固有特征有关。
– 相同信号可产生不同效应:如Ach可引起骨骼肌 收缩、心肌收缩频率降低,唾腺细胞分泌。 – 不同信号可产生相同效应:如肾上腺素、胰高 血糖素,促进肝糖原降解而升高血糖。