逆转座
分子生物学2-7章作业及答案

第二章一、名词解释1、DNA的一级结构:四种脱氧核苷酸按照一定的排列顺序以3’,5’磷酸二酯键相连形成的直线或环状多聚体,即四种脱氧核苷酸的连接及排列顺序。
2、DNA的二级结构:DNA两条多核苷酸链反向平行盘绕而成的双螺旋结构.3、DNA的三级结构:DNA双螺旋进一步扭曲盘绕所形成的特定空间结构。
4、DNA超螺旋:DNA双螺旋进一步扭曲盘绕所形成的特定空间结构,是DNA结构的主要形式,可分为正超螺旋与负超螺旋两大类。
按DNA双螺旋的相反方向缠绕而成的超螺旋成为负超螺旋,反之,则称为正超螺旋。
所有天然的超螺旋DNA均为负超螺旋。
5、DNA拓扑异构体:核苷酸数目相同,但连接数不同的核酸,称拓扑异构体6、DNA的变性与复性:变性(双链→单链)在某些理化因素作用下,氢键断裂,DNA双链解开成两条单链的过程。
复性(单链→双链)变性DNA在适当条件下,分开的两条单链分子按照碱基互补配对原则重新恢复天然的双螺旋构想的现象。
7、DNA的熔链温度(Tm值):DNA加热变性时,紫外吸收达到最大值的一半时的温度,即DNA分子内50%的双链结构被解开成单链。
Tm值计算公式:Tm=69.3+0.41(G+C)%;<18bp的寡核苷酸的Tm计算:Tm=4(G+C)+2(A+T)。
8、DNA退火:热变性的DNA经缓慢冷却后即可复性,称为退火9、基因:编码一种功能蛋白或RNA分子所必需的全部DNA序列。
10、基因组:生物的单倍体细胞中的所有DNA,包括核DNA和线粒体、叶绿体等细胞器DNA11、C值:生物单倍体基因组中的全部DNA量称为C值12、C值矛盾:C值的大小与生物的复杂度和进化的地位并不一致,称为C值矛盾或C值悖论13、基因家族:一组功能相似、且核苷酸序列具有同源性的基因。
可能由某一共同祖先基因经重复和突变产生。
14、假基因:假基因是原始的、有活性的基因经突变而形成的、稳定的无活性的拷贝。
表示方法:Ψα1表示与α1相似的假基因15、转座:遗传可移动因子介导的物质的重排现象。
【国家自然科学基金】_转座因子_基金支持热词逐年推荐_【万方软件创新助手】_20140803
2014年 序号 1 2 3 4 5 6 7 8 9 10
2014年 科研热词 推荐指数 诱导多能干细胞 1 细菌进化 1 细菌基因组结构平衡理论 1 甲基化 1 猪 1 油脂含量测定代谢通路调控 1 沙门菌致病岛7 1 毒力因子 1 微生物油脂生物柴油 1 启动子 1
2013年 序号 1 2 3 4 5 6 7 8 9 10 11 12
科研热词 预测 长末端重复(ltr) 重组杆状病毒 突变体 玉米 正反馈 早期生长应答因子1 反转录转座子 功能 分子标记 人钠碘同向转运体 mutator转座子
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1
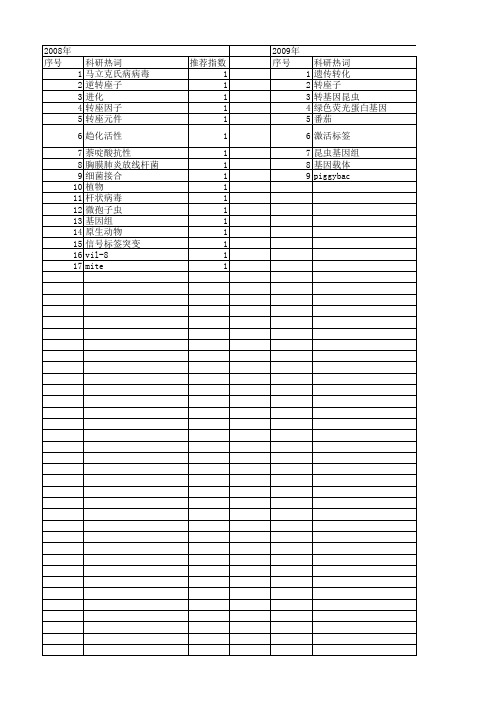
2009年 序号 1 2 3 4 5 6 7 8 9
科研热词 遗传转化 转座子 转基因昆虫 绿色荧光蛋白基因 番茄 激活标签 昆虫基因组 基因载体 piggybac
推荐指数 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
科研热词 重组抑制 重复序列 逆转座子 进化分析 转座子 葡萄球菌,金黄色 苹果 细菌毒素类 甲氧西林抗药性 毒力因子类 染色体 指纹图谱 异染色质化 克隆 东亚飞蝗 mariner irap-pcr dna甲基化 arac转录因子
推荐指数 1 1 1 1
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
科研热词 马立克氏病病毒 逆转座子 进化 转座因子 转座元件 趋化活性 萘啶酸抗性 胸膜肺炎放线杆菌 细菌接合 植物 杆状病毒 微孢子虫 基因组 原生动物 信号标签突变 vil-8 mite
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
分子生物学笔记

分子生物学笔记中心法则(Central dogma)DNA的组成DNA的融解温度Tm,高GC含量使得DNA的Tm升高,以及GC的体积较小,使得测得密度较大DNA变性的条件:有机化合物,高pH,低盐浓度探针和DNA杂交基因组是一个生物体的所有遗传信息的集合。
染色体的组成:DNA、蛋白质、RNA组蛋白Histones:五种H1、H2A、H2B、H3、H4核小体核心由8个组蛋白组成H2A、H2B、H3、H4各两个(组蛋白八聚体)146bpDNA核小体核心+H1+linkerDNA组成了染色体组蛋白的修饰乙酰化:转录激活,结构变松散DNA复制半保留复制DNA聚合酶只能从5‘到3’合成DNA(前导链)2. 3‘到5’的DNA聚合酶移动是半不连续复制(后随链,也是从5’-3‘合成)冈崎片段(DNA+RNA引物),后随链绕DNA聚合酶一圈,使得两者的复制方向相同细菌的后随链片段约1000nt,真核细胞中约200nt3. 引物和模板依赖DNA聚合酶不能从头合成DNA,必须前面由10-12nt的RNA引物提供3’羟基引物酶在合成DNA前加上一小段RNA引物复制叉两条母链解开时形成复制叉(replication fork)拓扑异构酶(DNA旋转酶,gyrases):去除DNA的超螺旋结构DNA解旋酶(DNA helicase):DnaB作用以及DnaA、DnaC等其他蛋白质SSBP:单链结合蛋白,稳定解旋后的单链引物酶:合成RNA引物,需要引发体DNA聚合酶Ⅲ(原核):同时合成两条链,链伸长DNA聚合酶Ⅲ:从5‘-3’合成DNA片段,然后删去RNA引物(具有核酸外切酶5‘-3’活性),发生缺口平移(缺口出现在引物和冈崎片段之间)DNA连接酶:去除引物后,连接冈崎片段和之前合成的片段滑动夹:保持DNA聚合酶不从DNA上掉下来端粒酶(telomerase):DNA复制酶只能5‘-3’合成DNA片段,因此DNA两端5’的RNA引物去除后不能让DNA聚合酶Ⅲ生成替换RNA引物的DNA片段(末端隐缩)。
进化发展中的基因转座与基因组演化

进化发展中的基因转座与基因组演化基因转座是指基因组中某一段DNA序列在基因组内发生切割和重新整合的过程。
这种现象在进化的历程中起到了重要作用,并对基因组的演化产生了深远影响。
本文将探讨基因转座在进化发展中的意义以及其对基因组演化的贡献。
1. 基因转座的意义基因转座是生物进化中基因多样性的重要源泉之一。
通过基因转座,新的基因序列得以引入或替换,从而增强了物种的适应性。
基因转座使得基因组更加灵活多样,使得物种能够更好地应对环境变化。
这种基因组重组的方式与基因突变相比,更能够促进物种的进化。
2. 基因转座的机制基因转座主要通过转座子实现,转座子是一类能够在基因组中移动的DNA片段。
转座子常见的类型包括类似转座子、复制转座子和逆转录转座子等。
这些转座子自身具有编码转座酶,可以识别特定的DNA序列并将其切割并重新整合到基因组的其他位置。
基因转座的过程可能会导致某些基因的失活或突变,但与此同时也为物种的进化提供了机会。
3. 基因转座的驱动力基因转座的发生主要受到两个驱动力的影响:一是转座子活跃性的调控,二是环境压力的选择。
转座子的活跃性受到多种因素的影响,包括基因座上的启动子或抑制子、转座子自身编码的酶的稳定性等。
而环境压力的选择则是指转座子在某些环境条件下获得更高的适应性,从而被选择进化。
4. 基因转座对基因组演化的贡献基因转座对基因组演化有着重要的贡献。
首先,基因转座增加了基因座的多样性,使得物种具有更高的适应能力。
其次,基因转座促进了基因重组,使得物种能够获得新的基因组组合,从而进一步增强了适应性。
最后,基因转座还可以改变基因表达模式,促进物种进化。
5. 基因转座的应用与展望基因转座的研究既具有基础性意义,也有着重要的应用前景。
对基因转座的深入了解有助于我们更好地理解物种的进化过程以及基因组的演化机制。
此外,基因转座技术也可以应用于基因工程和遗传改良领域,例如转基因技术和基因治疗等。
结论基因转座是进化发展中的重要过程,通过重新整合基因组中的DNA序列,促进了基因多样性和基因组的演化。
第六章DNA的重组与转座4

◎细菌的特异位点重组
鼠伤寒沙门氏菌H1鞭毛蛋白和H2鞭毛蛋白抗原 表达的互变,称为鞭毛相转变,这种转变是由H片段 倒位决定的,此倒位是由于特异重组位点hix发生重 组后引起的。
第六章 DNA的重组与转座
第一节 DNA的重组
一 概念 DNA 分子内或分子间发生遗 传信息的重新组合称为DNA的重组(遗传重组
或基因重排)。
重组和突变是产生遗传变异的两大主要 因素,但二者的分子机制截然不同:重组是 已经存在的遗传信息发生重排,而突变是改 变基因组中的碱基。但二者在分子机制上又 是相互影响的。
开靶DNA,再将转座子连接到靶的凸出单链上, 最后填补空缺完成转座。
按在转座过程中是否发生复制,可以分为:
复制型转座 非复制型转座
1979年Shapiro提出了转座模型 这个过程开始是形成单链转移复合体
[有时也称为交换复合体],供体和靶的单链 连接,这样转座子的两个末端分别与靶及供 体单链相连接。连接转移复合体产生了一个 交换形成的结构在双链转座子处结合在一起, 这个交换结构(即共同中间体)决定了转座 方式的命运。交换结构的每个交错末端含有 一个单链区。这个区域是个假的复制叉。它 提供了DNA合成的模板。若复制继续,那么 它的延伸将经过转座子,将它的链分开,在 其末端终止复制,然后由连接酶封闭缺口。 这个结构变成了共合体。在转座子与复制子 之间的连接处有DR。复制可能由宿主编码 的酶来完成。
这个模型解释了: 复制性转座在原来的位置上保留原有的Tn; 在新位置上转座子的两端出现正向重复靶序列; 转座过程中出现共合体。
复制型转座
转座子结构特征

转座子结构特征
转座子(Transposon)是一个基因组中可移动的DNA序列。
转座
子通过“剪切并复制”的机制活动,即从一个位置切断并携带自己的DNA序列,然后在目的地插入该序列。
转座子结构特征从其功能、组成和分类三个方面进行了描述。
功能方面,转座子通过自身编码的转座酶来切断、复制和插入目
标DNA。
这种特殊功能使转座子成为基因组进化和变异的重要驱动力。
组成方面,转座子包含两个反向重复序列和一个中央区域。
反向
重复序列是指转座子两端相似、但互补的DNA序列,它有助于转座子
在进化过程中扩复制。
中央区域是转座子的功能中心,它包括了转座
酶编码信息、调节序列以及其他功能相关的序列。
分类方面,转座子可以分为类别I、类别II和类别III三类。
类
别I转座子通过逆转录过程活动,可被分为线性和环形的两类;类别
II转座子主要通过剪切转座活动,是大多数真核生物中最多的转座子
类型;类别III转座子较少见,是通过入侵基因组DNA和产生单链嵌
合体来活动的。
总之,转座子的特点决定了其在基因组变异和进化中的重要作用。
对于目前基因组学和生态学的研究,深入了解转座子结构和特征将有
利于更好地理解生命进化的奥秘。
基因互补作用实例
基因互补作用实例
基因互补作用是指两个基因之间的相互作用,使其共同控制某些特定的生物学特征或生理过程。
以下是一些基因互补作用的实例:
1. 人类血型的遗传:人类血型有ABO和Rh两个系统,它们都涉及到基因互补作用。
ABO血型由三种基因A、B和O控制,其中A和B 是互补基因,O则是两种互补基因的缺失形式。
Rh血型也是由两种基因控制,一个是RhD阳性基因,另一个是RhD阴性基因。
只有当两个基因都是RhD阴性时,才会表现出Rh阴性血型。
2. 同源染色体的联会:在减数分裂过程中,同源染色体会发生联会,即互相交换一些基因段。
这种交换可以增加基因的多样性,从而增加后代的适应性。
如果两个基因位点距离很近,联会的概率就会很小,因为很难在两个基因之间发生交叉。
相反,如果两个基因位点距离很远,联会的概率就会很高,因为交叉点的可能性更大。
3. 逆转录转座子的移动:逆转录转座子是一种DNA序列,可以自主移动到基因组中的其他位置。
这种移动可以影响基因的表达,甚至导致基因功能的改变。
逆转录转座子可以插入到基因内部,使其产生部分或全部缺失,或者插入到启动子或其他调控序列中,从而影响基因的表达。
4. 基因表达的协同调控:许多基因在同一时间内被激活或抑制,以协同地控制某些生理过程。
这种协同调控通常是由多个转录因子和调控区域共同作用而实现的。
这些调控区域可以位于基因的上游或下游,可以与基因直接相接或通过某些中间因子进行交互。
这些实例都展示了基因互补作用的不同方面,强调了基因在生物体内的复杂互动和协同作用。
对于遗传学和生命科学研究来说,理解这些互补作用是至关重要的。
第3章 可移动的遗传因子
(2)反转录转座子 ) 转座因子的DNA 先被转录成 先被转录成RNA,再借助反转 转座因子的 , 录酶反转录成 录酶反转录成DNA,插入到新的染色体位点,从 反转录成 ,插入到新的染色体位点, 而引起基因的突变或重排。 而引起基因的突变或重排。正因为反转录转座子 是通过复制方式实现转座,每转座一次,其拷贝 是通过复制方式实现转座,每转座一次, 数就增加一份,因而极大地增加了基因组的大小。 数就增加一份,因而极大地增加了基因组的大小。
遗传学效应: 遗传学效应: 引起插入突变 产生新的基因 产生染色体畸变 引起生物进化
一、转座子的分类和结构特征
(1)DNA转座子 ) 转座子 通过DNA复制或直接切除产生可移动片段,该片 复制或直接切除产生可移动片段, 通过 复制或直接切除产生可移动片段 断插入基因组后导致基因突变或重排, 断插入基因组后导致基因突变或重排,一般不改 变基因组大小。 变基因组大小。 根据转座的自主性, 根据转座的自主性,这类转座子又分为自主转座 子和非自主转座子, 子和非自主转座子,前者本身能够编码转座酶而 进行转座, 进行转座,后者则要在自主转座子存在时才能够 实现转座(玉米的 体系)。 实现转座(玉米的Ac/Ds体系)。 体系
3.非自主性反转录转子 非自主性反转录转座子
(non-autonomous retrotrasposons) 通过DNA-RNA-DNA方式转座,本身不能编码蛋白质, 方式转座,本身不能编码蛋白质, 通过 方式转座 必须借助胞内酶才能实现转座。 必须借助胞内酶才能实现转座。 100-300bp,拷贝数达数十万份。 ,拷贝数达数十万份。 人类基因组中的非自主性反转录转座子主要为Alu 家族 人类基因组中的非自主性反转录转座子主要为 和加工后的假基因。 和加工后的假基因。
梨反转录转座子逆转录酶序列预测及其进化和转录分析
园艺学报,():– 2014411121962207 http: // www. ahs. ac. cn Acta Horticulturae Sinica E-mail: yuanyixuebao@收稿日期:2014–07–08;修回日期:2014–10–10 基金项目:国家自然科学基金项目(31201592)梨反转录转座子逆转录酶序列预测及其进化和转录分析蒋 爽,蔡丹英,滕元文*(浙江大学园艺系,农业部园艺植物生长发育与品质调控重点开放实验室,杭州 310058)摘 要:基于生物信息学方法对‘酥梨’基因组中不同类型的逆转录酶进行预测,共获得345条copia 类和99条gypsy 类逆转录酶。
通过系统聚类,copia 类逆转录酶可分为Ivana 、Ale 、TAR 、Angela 、Maximus 和Bianca 等6类;gypsy 类逆转录酶可分为Athila 、Tat 、CRM 、Reina 和Tekay 等5类。
序列比对结果显示梨中逆转录酶具有较高的异质性,copia 类逆转录酶序列分歧度为0.44,gypsy 类为0.38。
挑选出8类逆转录酶设计引物,并对梨属其它植物进行PCR 扩增,结果显示这8类逆转录酶广泛存在于梨属植物中。
在砂梨品种‘圆黄’的叶片、种子和果实中均发现该8类逆转录酶存在一定的转录水平,这是首次发现在梨属植物正常生长组织中逆转录酶发生转录。
关键词:梨;Ty1-copia ;Ty3-gypsy ;逆转录酶;预测中图分类号:S 661.2 文献标志码:A 文章编号:0513-353X (2014)11-2196-12Prediction ,Evolution and Expression Analysis of Reverse Transcriptase of LTR Retrotransposons in PearJIANG Shuang ,CAI Dan-ying ,and TENG Yuan-wen *(Department of Horticulture ,The State Agricultural Ministry Laboratory of Horticultural Plant Growth ,Development & Quality Improvement ,Zhejiang University ,Hangzhou 310058,China )Abstract :Different types of reverse transcriptase (RT )sequences in the whole genome of Pyrus pyrifolia white pear group ‘Suli ’were predicted by bioinformatics methods. A total of 345 RT sequences were obtained from copia group and 99 RT sequences were from gypsy group. The cluster analysis indicated that there were six lineages (Ivana ,Ale ,TAR ,Angela ,Maximus and Bianca )in copia group and five lineages in gypsy group (Athila ,Tat ,CRM ,Reina and Tekay ). Sequence alignment showed a high heterogeneity in both copia group and gypsy group ,and the divergence of RT in both groups was 0.44 and 0.38,respectively. Eight types of RT were selected to design primers ,each pair of primers showed clear amplified bands by PCR using genomic DNA of other Pyrus species. Eight types of RT were expressed with different levels in leaves ,seeds and fruits of ‘Wonhwang ’pear ,which was the first report on the expression of RT in the organs of pear trees under normal growing condition.Key words :Pyrus ;Ty1-copia ;Ty3-gypsy ;reverse transcriptase ;prediction* 通信作者 Author for correspondence (E-mail :ywteng@ )11期蒋爽等:梨反转录转座子逆转录酶序列预测及其进化和转录分析 2197反转录转座子是真核生物基因组中普遍存在的一类可移动的遗传因子,它们以RNA为媒介,在基因组中不断自我复制(Beauregard et al.,2008)。
基因工程 植物转座子在基因克隆中的应用
邯郸农业高等专科学校学报2000年第17卷第3期第20页J OURNAL OF HANDAN AGRICULTURAL COLLEGE2000117(3):20植物转座子在基因克隆中的应用蔡玉红 邢少辰(邯郸农业高等专科学校,永年057150) Ξ 摘 要:本文系统介绍了目前植物中转座子的种类、结构特征和在基因转化、基因克隆等方面的应用新进展,同时也详细介绍了类copia逆转座子在水稻上的应用研究。
关键词:植物;转座子;进展 转座子(transpos on)又称转座因子或者跳跃因子,这类因子实际上也是DNA片段,它可以在生物的染色体组中移动,从染色体的一个位点“跳”到另一个位点,还可以从一条染色体转移到另外一条染色体上,从而引起基因功能的改变。
转座子是1951年美国玉米遗传育种学家Mcclintock最早发现的,她是针对玉米籽粒中色斑不稳定现象而提出来的。
当时这是一个新概念,它突破了以往人们认为基因在染色体上的位置是固定不变的认识,所以一开始并不被大家接受,直到1967年在大肠杆菌(E.coli)的半乳糖操纵子研究中发现了这类插入序列,才得以被普遍认同。
现在的研究说明,在生物界中转座子是普遍存在的,并认为在生物的遗传进化方面有重要作用。
1 转座子的种类根据DNA的结构和转座的机理,可以将转座子分成二个大家族(superamil)。
第一类是转座子(transpos on),这类因子是基于DNA—DNA的转座过程,是最早发现的一类转座因子。
目前应用最成功的当属玉米中的Ac/Ds系统,除此之外,还有玉米中的spm因子,金鱼草中的T am因子等;第二类是逆转座子(retrotranspos on),这类因子的转座过程是基于DNA—RNA—DNA的转录和逆转录过程,因为和研究的比较清楚的逆转录病毒过程十分相似,故归为一类,属于这一类的因子有果蝇中的copia因子,酵母中的T y因子,烟草中的类T y1因子。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Changes in copy number
• TE累积的最直观的结果是基因组的增加,单是L1,Alu就给 基因组贡献了750Mb,基因组的增加是连续的过程,人类 在过去的6Myr累积了2000个L1,7000个Alu,1000个SVA copies,大约8Mb长添加到基因组中 • 同样重要的是,non-NTR的不断扩增形成了显著个体间多 样性。在人基因组序列中发现了数百新的活动元件插入, 这些人类特异性逆转座子插入在人类个体间的同源位点上 是多态性的,他们可以形成有信息含量的遗传标记,可用 来调查人类进化历史,群体结构和人口统计学。
逆转录转座子 在人类基因组进化中 的影响
• 摘要:
转座子能在基因组内移动使得他们影响基因组进 化。在过去8千万年灵长类进化中,不含长末端重 复序列(Non-LTR)的逆转座子(包括LINE1,Alu, SVA元件)扩增,到现在大约占了人类基因组的 1/3。 在这篇review里,我们着重探讨这几类元件,和 他们可能影响基因组的几种方式: 产生插入突变,造成基因组不稳定,改变基因表 达,引起基因革新(产生新基因)。
Evolutionary dynamics
进化动态学 • Non-LTR逆转座子在人类基因组进化中的 影响源自 • 1 copies数目多 • 2 数百年来连续的活性
影响逆转座子的细胞因素(box2)
1. 发生在转录水平L1逆转座子的调控
在进化中形成新的调控区,现在的L1的5‘端UTR包含了对转录起始 或激活有重要作用的转录因子结合位点。 在启动子区的DNA甲基化抑制L1的表达。 受转录后调控,例如,有RNA干扰形成的RNA沉默 L1的富集A编码区含有19个多聚腺核苷酸信号,这些信号能通过早熟 聚腺苷酸化截断全长的L1转录本,造成L1活性衰减。
• 逆转座子最直接的改变方法改变基因组的功能,潜在影响 进化,是通过插入到蛋白质编码区或调控区。由于这样插 入会产生直接被察觉的表型变化。例如:L1,SVA,Alu重新 插入引起的人类基因疾病一直累积着,已经有65例可遗传 性的疾病发现。
人类基因组中大 约有45%序列来 自转座子,其 中non-LTR逆转 座子 :L1, Alu, SVA,等
TE在基因组中的高密度提出问题:
TE如何影响人类基因组进化?
由于创新方法论,灵长类生物基因组测序的发展, 使得关注方向发生改变:关注TE研究—>关注TE活性
对基因组进化的影响程度研究
关于TE是如何影响人类基因组进化的研究使得在 基因组进化中量化TE活性影响成为可能。 例如,虽然长时间以来人们认为TE的重组能够引 发基因缺失,直到近来人类和其他灵长类在全基 因组范围内比较后,人们能确定在进化范围内TE 重组介导的缺失的量级大小和意义。
Human non-LTR retrotransposons
• L1 elements • Alu elements • SVA elements
L1 elements
结构:L1 是进化上保守的一个超基因家族, 5′、3′端为非编码区 (untranslated region , UTR) , 中间含二个开放读框(ORF) . ORF1 和ORF2。 5′UTR 含有一个内部启动子, 3′UTR 含一个非典型的 加尾信号和polyA 尾序列. 二个开放读框之间为66 bp 的间隔序列. ORF1 序列编码一种RNA 结合蛋白P40 , ORF2 蛋白是一种双功能 蛋白质, 近N 端有内切酶结构域( EN) , 近C 端为逆转录酶结构域 (RT) ,具有内切酶活性和逆转录酶活性。
在这篇review中,我们关注non-LTR逆转座子的 丰足性和活性如何影响人类进化的 • 首先,我们讨论non-LTR逆转座子的结构和他们 移动的机制 • 其次,我们探索non-LTR逆转座子的进化动力学, 即是,什么导致他们在基因组内进化上如此成功。 提出这个问题能够帮助我们理解TE影响基因组进 化的方式和范围, TE的影响包括造成基因组局 部不稳定,大范围结构多样性。不仅影响新基因 形成,还可以改变基因表达。
TE:转座子(jumping gene) 1.1940年发现,能在基因组内移动的离散的DNA 片段 2. 人类基因组有一半序列来自TE,而编码区序列 仅占1.5% 3.TE对基因组的影响是显著的
TE分为两类(根据是否有RNA参与)
DNA转座子
能从基因组中切除,像DNA一样移动,插入到新的基因 组位点,现在在基因组中已没有活性活动,但三千七百万 年前 灵长类进化过程中是有活性的,占基因组的3%
• 亚家族不仅年龄差异,而且定义亚家族的特征序 列变异和改变是层次性累积的,就是说两个亚家 族不是从一个祖先亚家族独立继承而来,而是大 部分亚家族存在于不间断的线性连续进化模式中, 这种模式就是一个亚家族连续的来自另外一个。
证据显示,人类基因组中所有的L1亚家族在过去40Myr连续的来自于 一个单一的家系。A1,SVA也发现了相似的模式。
Alu elements:
• copy num使它成为基因组最成功的TE. • 由左右二部分同源序列组成 , 其间为富含腺苷酸 的连接区(A2richlinker) . Alu 5′端含内部RNA聚合 酶三启动子, 3′端有polyA 尾序列. • 不能编码蛋白质,利用L1逆转座机制转座
• Alu 元件没有RNA聚合酶三终止信号,ALU 转录本延伸到下游侧翼序列,直到发现终 止子。无编码能力,非自主性转座子,他 利用L1 元件逆转录分子机制,因此被称为 “寄生虫的寄生虫”
Stealth drivers and long-term evolution 潜在的动力和长期进化
• 逆转座子活性如何保存这么长时间 Alu Yb 家系的进化历史的重建显示大概起源于18~25Myr 前早期人科进化 在早期几百万年间, 在人类基因组中Alu Yb 家系扩展了 ~2000个copies,而在人科灵长类非人类的生物中仅有几 个元件,Alu Yb 元件尽管能够以物种特异的方式产生很多 copies,但能在基因组内以很低甚至没有转座活性存在 15~20Myr 以上事例说明,长期存在的,低活性的source 元件能作为 潜在动力,偶尔产生copy,其中一些是有很强活性的。然 而活性高的master 元件可能被察觉且经历负选择。低活 性的潜在动力,可能允许Alu Yb 家系长期存在,移动性的 衰减可能是多种逆转座子的普遍的进化策略。 因此,能保持低的但适度的逆转座活性能力是人类 nonLTR逆转座子能长期有活性的一个重要特征。
A,插入诱变 B产生并修复DNA双链缺 口 C微卫星形成 D基因转换
E插入介导删除 F易位重组 G转导
Local genomic instability 局部基因组不稳定
• 逆转座子可以从不同途径造成基因组不稳 定,这一节我们在局部基因组范围内考虑 逆转座子的影响
Insertion mutagenesis 插入诱变
L1的两种结果差别,可能是由于使用的方法的假设不同 这种差异负 选择。
SVA:1 insertion/900birth 基于:基因组比较 (数据不全,不太确定)
• 尽管新的可遗传的逆转录发生在生殖系,同时也发生在体 细胞,从癌症到脑发展过程中一直存在,逆转录引起的体 细胞多样性可能深入了解TE的生物学和对人类的影响。 • TE的扩展率不是一成不变的,例如:Alu元件在大约 40Myr一个扩展高峰插入,大约1 insertion/1birth,相似的, 过去70Myr进化,L1扩展率发生变化,最显著L1亚家族的 扩展是在12-40Myr前。 • 人与黑猩猩基因组比较提供根据,证明近代LINE1,Alu, SVA逆转录率是发生变化的,在6Myr前两个物种发生分歧, 物种特异性元件插入的不同数目来显示出。 • 在短时间内扩展率的波动显示了在宿主群体水平的影响
Impact on genome evolution 对基因组进化的影响
Amplification rates 扩展速率 L1,Alu,SVA连续的活性和在基因组中长年累月积累,使得他们对 灵长类进化在功能和结构上产生巨大的影响。评价这种影响,一种想 法是考虑在生殖系中逆转录发生频率 Alu:目前速率 1 insertion/20 births 基于: 比较核苷酸替代和从头插入的引起疾病的频率 人与黑猩猩基因组比较和多重人类基因组序列比较 L1: 1 insertion/20 births 基于:从头插入的引起疾病的频率 1 insertion/200 births 基于:基因组比较
SVA
elements
• 结构包括六聚物重复区,Alu类似区,数量变化串 联重复区,HERV-K10类似区和变化长度的寡聚A 的信号结尾区 • RNA聚合酶二转录,由L1逆转座推测其转座。
其他non-LTR逆转座
• 除了上述的,还有古老的没有活性的nonLTR逆转座家族,占基因组的6%,这些元 件提供了很多的分子化石记录,能够证明 TE和人类基因组的长时间关系
Subfamily structure and source elements 亚家族结构和源元件
• 关键概念: L1,Alu,SVA序列进化动态学相关的一个概 念是:基于所有亚家族成员特异性共享特 征核苷酸替换,插入,删除,这些元件能 被分为亚家族或是分支。
例如:在人类基因组中Alu亚家族发现超过200个, 但是年轻的SVA家族只有6个,
2. 对ALU的活性的影响。
初始序列影响活性,高密度的CPG二核苷酸5’甲基胞嘧啶残基去氨 基酸化促进突变的积累,突变的累积能改变重要的模式。多聚A尾 的长度和同质性,ALU要插入的基因组环境,寡聚A尾和RNA聚合酶 三终止子的距离。总的来说,Alu copy与家族共有序列达到10%的 差异时,保持活性的可能性就很小。
转座机理: TPRT机制 (target-primed reverse t ranscription靶引物反转录 )
TPRT机制
Mass 量最多,在基因组中最成 功的TE。 并不是所有的L1 copies有逆转 座能力的,作为TPRT机制的产 物并且随着时间衰退,大部分 L1 copies由于被截断,内部重 排,突变等而失去活性,人类 基因组中>500,000的L1元件 中只有不足100个copies是有功 能的 TSD:靶位点重复序列