无脊椎动物的比较完整版
无脊椎动物体腔的发育比较
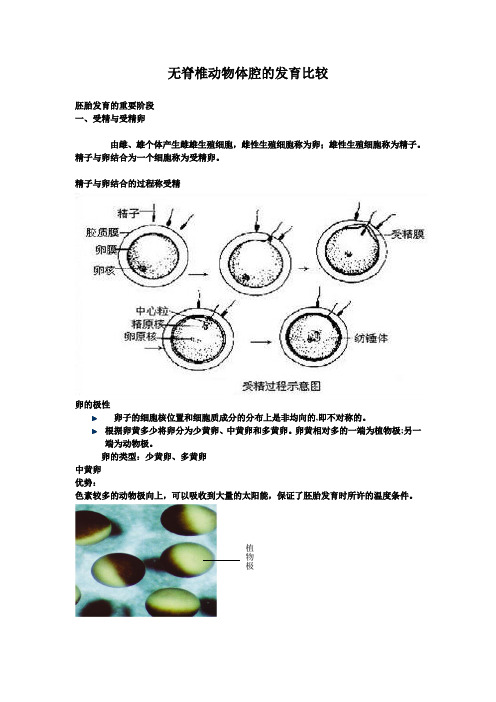
无脊椎动物体腔的发育比较胚胎发育的重要阶段 一、受精与受精卵由雌、雄个体产生雌雄生殖细胞,雌性生殖细胞称为卵;雄性生殖细胞称为精子。
精子与卵结合为一个细胞称为受精卵。
精子与卵结合的过程称受精卵的极性卵子的细胞核位置和细胞质成分的分布上是非均向的,即不对称的。
根据卵黄多少将卵分为少黄卵、中黄卵和多黄卵。
卵黄相对多的一端为植物极;另一端为动物极。
卵的类型:少黄卵、多黄卵 中黄卵 优势:色素较多的动物极向上,可以吸收到大量的太阳能,保证了胚胎发育时所许的温度条件。
二、卵裂受精卵的分裂==卵裂一种特殊的分裂方式,即在每次分裂之后,未到新细胞长大就继续不断下去;因此分裂成的细胞就越来越小,这些细胞叫做分裂球。
卵裂的方式:由于不同类动物卵细胞内卵黄多少和卵黄在卵内分布情况的不同,卵裂的方式也不同。
1、完全卵裂:在分裂时,受精卵分裂为完全分离的单个细胞.①如果分裂的子细胞形状大小相同的卵裂叫完全均等卵裂(等裂)。
多见于卵黄较少,分布均匀的海胆,文昌鱼等的卵.②如果分裂的子细胞大小不等,则称为完全不等卵裂(不等裂)。
如海绵动物,蛙类等卵黄分布不均匀的卵。
2、不完全卵裂:受精卵分裂不彻底,仅一部分发生分裂,即子细胞不完全分离。
①若卵黄多,细胞质和细胞核集中于卵一端,分裂仅限在不含卵黄一端的叫盘裂,如乌贼,鸟,鱼的卵。
②如果大量卵黄集中在卵的中央,分裂只限于卵表面的叫表裂。
如昆虫的卵。
三:囊胚的形成卵裂后分裂形成中空的球状胚,称为囊胚。
囊胚中间的空腔叫囊胚腔。
囊胚壁的细胞层叫囊胚层。
四、原肠胚的形成出现了内、外两胚层和原肠腔1.内陷:由囊胚的植物极细胞向内陷入,最后形成二层细胞,在外面的细胞层叫外胚层,向内陷入的细胞层叫内胚层。
内胚层所包围的腔,形成原肠腔。
原肠腔向外的通口称为原口或胚孔。
如海星2.外包:植物极细胞由于卵黄多分裂慢,动物极细胞卵黄少分裂快,结果动物极细胞逐渐向下包围了植物极细胞,成为外胚层。
内部的植物极形成细胞成为内胚层。
无脊椎动物的主要类群以及脊椎动物的特征比较

环节动物
蚯蚓、水蛭、沙蚕
身体由体节组成
软体动物
河蚌、蜗牛、乌贼、鲍鱼
身体柔软,有各种足。
棘皮动物
海星、海参、海胆
节肢动物
昆虫纲
蝗虫、蜜蜂
1、身体由体节组成,并且分部;2、体表有外骨骼;3、足和解角分节。
甲壳纲
沼虾、河蟹
蛛形纲
蜘蛛
多足纲
蜈蚣
【
类别
皮肤
循环系统
呼吸系统
生殖发育
体温
其它
鱼类
覆盖鳞片
心脏一心房一心室
【
类群
实例
特征
原生动物
草履虫、变形虫、疟原虫
单细胞动物,是动物界最原始的类群。
腔肠动物
水螅、海葵、珊瑚虫、海哲
1、生活在水中;2、体壁由内胚层、外胚层和中胶层构成,有网状神经;3、体内有消化腔;4、有口无肛门。
扁形动物
猪肉绦虫、血吸虫
这两类动物多寄生生活,运动器官和感觉器官退化,生殖器官发达。线形动物用鳃呼吸体外受精用鳍游泳
两栖类
皮肤裸露,能分泌黏液,有辅助呼吸的作用。
心脏二心房,一心室
幼体用鳃呼吸;成体一般用肺呼吸。
体外受精,变态发育。
不恒定
爬行类
体表覆盖着鳞片或甲
心脏二心房,一心室,心室内有不完全隔膜。
用肺呼吸
体内受精,陆上产卵,表面有坚韧的卵壳。
不恒定
鸟类
被覆羽毛
心脏四腔
用肺呼吸,并且有气囊辅助呼吸,双重呼吸。
卵生
恒定
有喙无齿,前肢变成翼,骨中空,内充气体。
哺乳类
体表被毛
心脏四腔
用肺呼吸
胎生,
哺乳
无脊椎动物部分的总结(by吴筱璇)

原生动物门鞭毛纲虫体具鞭毛,为运动胞器;同时具有感觉;捕食或附着功能。
具有三种营养方式。
繁殖分为无性(纵向的二分裂)和有性。
自由生活或寄生。
多数种类为单细胞,少数为多细胞。
肉足纲有些种类体表没有坚韧的表膜,形态不固定;有些种类外面有壳外包;运动主要依靠伪足(Pseudopodium);伪足的形成是细胞质中的内质和外质的相互转化而成的;伪足还具有摄食和排泄等功能;无性生殖为二分裂;生活史复杂种类的有性生殖为受精。
孢子纲全为寄生性。
细胞内寄生阶段一般无运动细胞器,如有伪足则是摄食作用。
生殖方式包括无性和有性两类。
无性生殖有裂体增殖(schizogony)产生裂殖子,以及孢子增殖(sporogony)产生具感染性的子孢子(sporozoite);有性生殖是通过雌雄配子结合进行的配子生殖(gametogony)。
以上两种生殖方式或可在一个宿主或分别在两个宿主体内完成.有或无宿主更换。
纤毛纲体表覆有纤毛,为运动、感觉和捕食的胞器;生殖方式为无性(横向二分裂)和有性(接合生殖conjugation);营养方式为异养;细胞和有两种类型:大核(营养)和小核(繁殖)小结个体细小, 多为单个细胞构成。
以各种胞器完成各种生活机能。
自由生活原生动物的运动胞器为鞭毛, 纤毛或伪足。
营养方式包括植物性、动物性和腐生性;呼吸靠体表进行;排泄依靠体表或伸缩泡完成;体形结构多样化;生殖方式分无性(二分裂, 出芽和复分裂)和有性(受精和接合);分布非常广泛。
海绵动物门小结原始的多细胞动物。
体形多为不对称。
细胞有分化现象,但没有器官、系统和组织。
体壁由两层细胞构成,外侧为扁平的表皮细胞;内侧为领细胞。
中间为中胶层。
胚胎发育有逆转现象。
腔肠动物门水螅纲一般为小形的水螅型和水母形动物水螅型结构较简单,只有简单的消化循环腔水母型有缘膜,触手基部有平衡囊(感觉器官)水螅群体以无性出芽的方式产生单体的水母型,水母型又以有性生殖的方式产生水螅型群体。
无脊椎动物总结10

节肢动物-不完全变态(渐变态和半变态)
- 完全变态
分节(metamerism)
分节现象从环节动物开始,标志着进入高 等无脊椎动物。
动物的分节是从 环节动物的同律分节, 发展到节肢动物的异律分节而逐步进化的。
运动:无附肢→附肢
鞭毛、纤毛、伪足(运动细胞器) 疣足、刚毛+肌肉(原始的附肢) →节肢 中胚层出现后,肌肉参与运动
个体发育(development)
直接发育 间接发育: 单幼虫:腔肠动物--浮浪幼虫; 环节动物--担轮幼虫; 多幼虫(寄生中间宿主;自由生活)
毛蚴、胞蚴、雷蚴、尾蚴、囊蚴 甲壳动物——无节幼体、溞状幼体……. 棘皮动物——耳状幼虫、羽腕幼虫、短腕幼虫 软体动物——担轮幼虫、面盘幼虫、钩介幼虫。
神经系统
神经网( ? 动物)→梯型神经系统 ( ? 动物)→索式神经系统( ? 动物、 ? 动物)
(节肢动物的神经生殖方式:
无性生殖:迅速增加个体数量 细胞分裂:二分裂,裂体生殖 机体:出芽、芽球、再生 有性生殖:接合、配子、幼体、孤雌
生殖腺:
生殖腺不固定( 动物以前)→出现了固定的 生殖腺和生殖导管( 动物开始) 雌雄同体→雌雄异体
消化系统:提高消化吸收效率 细胞内消化→细胞外消化
→有口无肛门不完全消化系统(腔肠、扁形动物) →有口有肛门完全消化系统,食物与粪便分开, (假体腔动物),消化管分为前肠、中肠和后肠
1消化道出现肌肉、消化腺(环节动物)
→分化分工、结构复杂化,咽、食道、胃(贲门胃 幽门胃)、肠 →出现各种口器(节肢动物),附属结构:齿舌、 晶杆、胃盾(软体动物 ),磨胃(甲壳动物) 2腺细胞→腺体分泌消化酶,咽腺、食道腺、肝,肝 胰脏
呼吸系统
无脊椎动物的纵向比较与复习

高等
比较原肾管protonephridium 与后肾管或肾管metanephridium
器官
动 物
结构与机制
作 用
原 肾 管
1.盲管不是两端开口 2.火焰细胞(flame cells) 涡虫,轮虫 细胞管状,管中有纤毛, 等 深入排泄管中.其纤毛 摆动可将管中液体火从 排泄孔流出
排出体内多余 水分,同时把某 些离子重吸收. 其实,废物还是 主要由体表排 出.
消化系统
原生动物:胞口、胞咽、胞肛 海绵动物:水沟系统 腔肠动物:消化循环腔(原始消化腔) 扁形动物:口、咽、肠(不完全消化系统) 线虫动物:出现肛门(完全消化系统) 环节动物:消化管分为前、中、后肠,可进行物理性 消化 软体动物:口、口腔、胃、肠、肛门 节肢动物:口、咽、食道 、胃(贲门胃、幽门胃)、 肠、直肠、肛门
棘皮动物的体腔
次生体腔发达,具有 特殊的水管系统、血 系统和围血系统 水管系统具有摄食、 运动、感觉、呼吸和 排泄功能 体腔中充满体腔液, 起到运输营养物质的 作用
呼吸系统
原生动物门、扁形动物门、线虫动物门(体表) 环节动物门(体表):多毛类(疣足)、尾鳃蚓(鳃) 软体动物门(鳃、外套膜):陆生如蜗牛种类(肺囊) 节肢动物门:甲壳纲(鳃)、鲎(书鳃)、蜉蝣幼虫 (气管鳃)、蛛形纲(气管和书肺)、昆虫纲(气管) 棘皮动物门:海星(管足和皮鳃)、海参(呼吸树)排泄
无脊椎动物排泄系统的演化
动物分类 原生动物 门 海绵动物门 腔肠动物门 扁形动物门 线虫动物门 环节动物门 软体动物 门 节肢动物门 后肾管 肾脏 绿腺 马氏管 主要起排 协作用 中胚层与外 胚层 中胚层与外 胚层 与后肾同源 内胚层或外 胚层 原肾管 排泄方式/器官 伸缩泡 细胞表面 体表 功能 主要调节 水分,同 时起排泄 作用 外胚层 来源 进化程度 低等
七年级无脊椎动物知识点

七年级无脊椎动物知识点无脊椎动物是指没有脊柱的动物,通常比较小,但数量庞大,是生物链中很重要的一环。
在七年级生物学课中,学生需要学习很多与无脊椎动物有关的知识。
本文将从多个方面为读者介绍七年级无脊椎动物知识点。
一、无脊椎动物的分类根据其特征,科学家将无脊椎动物分为很多类别,比如蜗牛、水母、虫子、甲壳动物和软体动物等。
这些动物都有着不同的形态和功能,它们之间的分类是根据它们的外观、内部结构和生活习性进行的。
二、无脊椎动物的特征无脊椎动物在结构上比较简单,它们没有明显的脊柱,因此身体通常比较柔软,可以自由弯曲和伸展。
它们的运动过程也比较有趣,例如像蚯蚓一样的动物会通过留下一些粘液来移动,而像是水母的动物会通过腕足的律动来游泳。
另外,无脊椎动物的简单结构不代表它们在生态系统中的作用不重要,有些无脊椎动物会在环境中扮演着很重要的角色。
三、无脊椎动物的生态角色无脊椎动物在自然界中有着重要的生态角色。
它们可以分解腐殖质,清除有害生物,帮助植物授粉,还有的能被其他动物作为食物。
因此,无脊椎动物在海岸生态系统的食物链中扮演着重要的角色,很多动物会依赖于它们来生存。
四、常见的无脊动物无脊椎动物有很多种类。
我们常见到一些昆虫,比如蝴蝶和蚂蚁等。
在水中我们看到的虫子和蠕虫不仅数量庞大,也非常多样化。
此外,我们在海洋中也可以看到很多无脊椎动物,例如海星、水母和海螺等。
五、如何保护无脊椎动物无论是在我们现在的生活中还是未来的生活,保护自然界和生态环境都是非常重要的事情。
在实际行动中,我们应该尽可能地减少对自然生态环境的干扰、捕捞、采集无脊椎动物等行为。
此外,还应该着重加强对于有些濒危的无脊椎动物的保护,将它们从濒危物种中解救出来。
以上是七年级生物学中的无脊椎动物知识点,从无脊椎动物的分类、特征、生态角色、常见种类和保护等角度分别进行了介绍。
阅读本文后,相信大家会对无脊椎动物有更多的了解和认识。
【期末复习总结】无脊椎动物总结
P 全部海水 发达 -胃层 有 皮层和胃层
生活史
只水螅型
二、主要纲的比较
涡虫纲 代表动物 表皮 消化管/营养 口吸盘 节片 排泄孔 生殖 生活方式 杜氏涡虫 纤毛上皮 杆状体 分支 吞食 无 无 背面,成对 两性;再生 多数自由 吸虫纲 华枝睾吸虫 合胞体 无纤毛和杆状体 分支少 吞食、渗透 有 无 1 个,后端 两性;幼虫生殖 内、外(单殖)寄 生 绦虫纲 猪带绦虫 合胞体,具微绒毛 无纤毛和杆状体 无 渗透 无 有 1 个,后端 两性,各节片均具 生殖器官 脊椎动物消化道 寄生
刚毛(生于体壁) 无
主要纲特征比较
瓣鳃纲 数海、淡水,底栖(固 着、附着、埋栖、匍 匐)、少动。 左右对称 无明显的头部 无齿舌 腹足纲 海、淡水、陆地,底栖、 活动能力较强,多匍匐生 活。 不对称或外形次生对称 头部明显,具眼、触角等 感觉器官 具齿舌 海水 头足纲
左右对称 头部明显,眼发达 具齿舌
四足类
羊膜类
恒温动物
分裂方式
第9节 总结
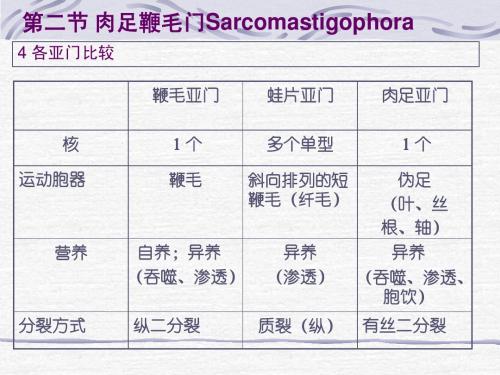
核 肉足鞭 毛门 盘蜷门 单型 单型 运动 胞器
鞭毛/ 伪足
顶复 合器 无 无
粘网 通路 无 有
孢子 无 无
生殖方式 二分/质裂 有性 游孢子 裂体 配子 孢子 复分裂、 孢子 二分/复分 /孢子 横裂/出芽 接合
营养 方式 自养 异养 异养
伪足
顶复门
粘体门 胶虫门 微孢子 门 纤毛门
足斧状,适于钻掘
双壳发达,常坚厚 心耳、鳃、肾等对称
足块状,适宜爬行
单壳,常螺旋状 心耳、鳃、肾等只单侧保 留
足特化为腕和漏斗
单壳(内或外),多退化 心耳、鳃、肾等对称
开管循环
发育过程中内脏不扭转
(完整版)无脊椎动物重点知识

第五单元第一节腔肠动物和扁形动物腔肠动物1、水螅生活在水流缓慢,水草繁茂的清洁淡水中,通常会附着在水草上,身体几乎透明,顶端有多条细长的触手,呈辐射对称。
(辐射对称的意义)2、水螅的体壁由内胚层和外胚层两层细胞构成,两层细胞之间填充着他们分泌的胶状物质,中间的空腔叫消化腔,消化腔与口相通,食物在消化腔中消化,食物残渣从口排出。
外胚层有刺细胞,它是攻击和防御的利器,刺细胞在触手处尤其多。
3、水螅主要通过出芽方式进行生殖。
在营养条件适宜时,利用芽体进行出芽生殖,属于无性生殖;也可以进行有性生殖。
4、常见的腔肠动物还有海蜇、海葵、珊瑚虫、水母等,腔肠动物大多数种类生活在海洋中。
海蜇加工后可以食用。
珊瑚是珊瑚虫分泌的石灰质物质,珊瑚堆积构成珊瑚礁,他们为海洋生物提供了重要的栖息场所和庇护地。
然而由于过度采挖、环境污染以及全球变暖等原因,珊瑚礁破坏严重,珊瑚虫大量死亡。
5、腔肠动物的特征:①身体呈辐射对称②体表有刺细胞③有口无肛门。
扁形动物6、涡虫背面呈褐色。
背腹扁平,体形象一片柳叶。
身体呈两侧(左右)对称。
涡虫身体前端呈三角形,头部背面有两个可以感光的黑色眼点。
消化器官由口、咽、肠等器官组成,没有肛门。
身体的两侧或腹面通常有专门的运动器官。
7、大多数扁形动物消化器官简单,有的甚至没有专门的消化器官。
他们的生殖器官却特别发达。
身体呈两侧对称,前端感觉器官集中,能最先感知外界刺激。
8 、华枝睾吸虫、血吸虫、绦虫、涡虫都属于扁形动物。
9、多数扁形动物是营寄生生活的。
涡虫是营自由生活的。
吃未煮熟的鱼虾容易感染华枝睾吸虫,进入含有钉螺的水域容易感染血吸虫。
10、扁形动物的主要特征:①身体两侧对称,②背腹扁平,③有口无肛门。
11、比较腔肠动物和扁形动物第二节线形动物和环节动物线形动物1、蛔虫寄生在人的小肠里,靠吸食小肠中半消化的食糜生活。
它的身体呈圆柱形。
前端有口、后端有肛门、体表具有角质层它可以起保护作用。
消化管的结构简单,肠仅由一层细胞组成,可消化食糜。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
无脊椎动物的形态结构与生理一、体制指动物躯体结构的排列形式和规律。
一般分为有规律可寻(对称)无规律可寻(不对称)•原生动物不对称(尾草履虫、变形虫)球辅对称(太阳虫、团藻虫)辐射对称(钟虫)球辐对称:通过身体中心点可分成许多相同的两半。
•海绵动物不对称或辐射对称•腔肠动物辐射对称或两辐对称辐射对称:指通过身体的中央轴有许多个切面可以将身体分为左右相等的两部分(对称面)。
主要适应附着、漂浮、及不太运动的生活方式。
两辐对称;通过动物体轴仅可分成两个对称面。
(如海葵)•扁形动物两侧对称;通过体轴只有一个对称面。
两侧对称的重要意义;(1)使动物身体明显地分为前后、背腹和左右,由不定向运动变为定向运动。
(2)使动物由水中固着或漂浮生活向水底爬行生活及陆地爬行奠定了基础。
•扁形动物以后的各类群全部是两侧对称。
仅有两个特例;1. 软体动物腹足纲;由于胚胎发育发生了扭转,因此成体不对称。
2. 棘皮动物早期发育的羽腕幼虫及短腕幼虫(两侧对称),成体由于适应不太运动的生活方式产生了次生性的辐射对称。
二、胚层与体腔1.胚层指多细胞动物胚胎发育时期由于细胞分化而形成的特殊区域。
多细胞动物早期的胚胎发育;受精→卵裂→囊胚→原肠胚→中胚层和体腔的形成→胚层分化•海绵动物没有明确的胚层分化,体壁由两层细胞构成。
由于胚胎发育的“逆转现象”,故不能称其为外胚层和内胚层(只称皮层和胃层)。
•腔肠动物两个胚层(外胚层、内胚层)中胶层不是细胞结构。
•扁形动物以后各类群由于出现了中胚层,故都称为三胚层动物。
2. 体腔指动物体消化道与体壁之间的腔隙。
•扁形动物及以前各类群没有体腔•原体腔(线形动物)动物出现原体腔原体腔指胚胎发育的囊胚腔演化形成的体壁与脏壁之间的腔隙。
原体腔(假体腔、初生体腔)特点:(1)只有体壁中胚层,没有肠壁中胚层和体腔膜。
(2)腔内充满体腔液。
(3)体腔对外没有孔道。
•环节动物具有真体腔(次生体腔)蛭类除外。
真体腔指中胚层的脏壁与体壁分离后,形成的动物内脏和体壁之间的腔隙。
真体腔的重要意义:(1)肠壁出现了肌肉,为消化道的进一步分化打下了物质基础。
(2)导致了循环系统的形成,改善了排泄、生殖、神经系统的功能。
(3)体腔液有参与循环、运动、维持体形的作用。
真体腔形成的方式端细胞法(裂体腔法) 原口动物在胚孔两侧的内、外胚层交界处植物极的一个细胞(端细胞)分裂后移入内、外胚层之间,经过不断分裂形成了中胚层带,随后在中胚层带中间开裂形成真体腔。
如环节动物等。
体腔囊法(肠体腔法) 后口动物的原肠背部两侧的内胚层向外形成一对囊状突起,并不断扩展并与原肠的内胚层脱离形成中胚层带,在内、外胚层之间形成中胚层和体腔。
如棘皮动物等。
•软体动物混合体腔(并存式混合体腔)指真体腔退化变小,初生体腔扩大并形成血窦。
如河蚌的真体腔只留下围心腔、生殖腔和排泄管腔。
•节肢动物混合体腔(打通式混合体腔)真体腔不发达,围心腔等破裂并与初生体腔打通。
故又称血腔。
•棘皮动物真体腔发达,又拓展成为水管系统和围血系统。
☆棘皮动物是后口动物,其真体腔的形成为肠体腔法。
三、分节与分部1. 分节(真分节)指由中胚层起源的结构将动物体分成许多形态、机能相似的体段,是无脊椎动物发展到高级阶段的重要标志。
同律分节为一种原始的分节现象,其特点是身体除头节和最后一节以外,其它体节在形态和机能上基本相似。
异律分节身体部分形态与功能相似的体节常相互愈合,同时各部分的机能发生分化。
动物分节的重要意义;(1)由于重要的器官在每个体节重复排列,使动物的新陈代谢水平及对外界环境的适应能力增强。
(2)使动物的运动能力加强。
•原生到原体腔动物体不分节★绦虫有节片,蛔虫有环纹;但均为外胚层形成的产物,非真分节。
•环节动物出现真分节(同律分节)•软体动物不分节•节肢动物真分节(异律分节)•棘皮动物幼体内部分节,成体不分节。
2. 分部在异律分节的基础上,外表的分节现象消失而形成了体区(部)。
•分部是节肢动物分类的依据甲壳纲、肢口纲、蛛形纲分为头胸部和腹部;原气管纲、多足纲分为头部和躯干部;昆虫纲分为头、胸和腹部;四、体壁与骨骼•腔肠动物外胚层中胶层内胚层水螅体壁主要有六种细胞;皮肌细胞、间细胞、刺细胞、感觉细胞、神经细胞和腺细胞构成。
外胚层常分泌角质、石灰质骨骼。
外胚层皮肌细胞的肌原纤维方向与螅体的纵轴平行排列,因此其收缩时可使水螅体和触手变粗缩短。
内胚层的肌原纤维方向与螅体纵轴垂直排列,其收缩可引起水螅体和触手变细变长。
•扁形动物皮肌囊结构表皮层外胚层柱状上皮细胞排列组成基膜非细胞构造,具有弹性肌肉层中胚层形成,分外环、中斜、内纵肌实质中胚层合胞体的网状组织,有输送和储存营养物、代谢产物、再生、生殖等功能。
寄生生活种类体表发生特化;纤毛消失,上表皮特化为富含粘多糖的合胞体结构,具皮棘,皮层的细胞核埋在肌肉层之下,微绒毛,孔道。
皮肌囊由外胚层形成的表皮与中胚层形成的肌肉层相互紧贴而构成的体壁呈囊状结构包裹动物全身,称之为皮肤肌肉囊。
具有保护、运动等功能。
•原体腔动物皮肌囊结构角质层非细胞结构有保护和抵抗消化酶作用表皮层合胞体结构,其细胞界线不明显肌肉层肌原细胞构成原体腔由胚胎时期的囊胚腔演化形成肠壁无肌肉层•环节动物皮肌囊结构角质膜(非细胞结构)表皮层(柱状细胞、刚毛、腺细胞和感觉细胞)肌肉层(外环肌、内纵肌)壁体腔膜真体腔真体腔脏体腔膜(黄色细胞)肌肉层(纵肌、环肌)肠上皮•软体动物表皮有纤毛,并形成外套膜。
外套膜(Mantle)是软体动物背侧的体壁向腹面延并常包裹着动物整体或一部分,具有保护、呼吸和运动等功能。
•软体动物表皮有纤毛,并形成外套膜。
外套膜(Mantle)是软体动物背侧的体壁向腹面延并常包裹着动物整体或一部分,具有保护、呼吸和运动等功能。
贝壳(Shell)是由外套膜外上皮分泌的钙质保护性外壳。
贝壳一般包括三层:角质层(壳皮)角化蛋白成分、黑褐色、较薄。
棱柱层(壳层)碳酸钙、硫酸锶成分,白色,较厚。
珍珠层(壳底)成分同于棱柱层、极厚、有金属光泽。
•节肢动物体表被有厚而坚硬的体壁,又称几丁质外骨骼;由表皮(称为外骨骼)、上皮和基膜三部分组成。
上表皮蜡质,拒水性,防止水分渗入或蒸发。
外表皮较薄,含蛋白质、几丁质、钙盐坚硬。
内表皮较厚,含蛋白质、几丁质,柔软。
上皮外胚层的多角形细胞层,分泌外骨骼。
基膜由上皮向内分泌一层薄的基膜。
•蜕皮:节肢动物身体长到一定限度后,在内分泌激素控制下内表皮溶解、外表皮脱出并重新形成新表皮的过程。
(两次蜕皮之间为幼虫的龄期,龄期等于蜕皮加一)•棘皮动物由角质层、表皮、真皮、围脏膜(体腔膜)构成。
表皮上有纤毛,真皮内有骨骼。
五、消化系统•原生动物消化细胞器食物泡,细胞内消化。
•腔肠动物出现消化系统,原始的消化循环腔,无肛门。
高等种类具有分化(如胃、胃囊、辐管系统、隔膜等。
细胞内外消化兼行。
如;涡虫的消化道由口、咽和肠三部分组成。
但吸虫纲动物消化管退化,绦虫纲动物消化管消失。
•扁形动物不完全的消化管,细胞内外消化兼行。
•原体腔动物完全消化管(出现肛门),细胞外消化,但肠壁无肌肉。
如蛔虫的消化道组成为;口→咽→肠→直肠→肛门分为前肠、中肠和后肠;前肠(口、咽)、后肠(直肠和肛门)•环节动物完全消化管,细胞外消化,肠壁出现肌肉,消化道进一步分化。
如环毛蚓的消化道组成为;口→咽→食道→嗉囊→砂囊→胃→肠(有盲道和盲肠)→肛门。
出现消化腺;咽腺、钙质腺、胃肠腺、黄色细胞。
蛭类的咽头腺可分泌蛭素,具有发达的的嗉囊。
•软体动物与环节动物相似。
但出现了真正的肝脏。
河蚌还具有特殊的晶杆胃及直肠穿过心室等特征。
除瓣鳃类外一般具有齿舌。
节肢动物基本同于环节动物。
昆虫出现了特殊的取食口器;如咀嚼式、刺吸式、虹吸式、舐吸式、嚼吸式。
蝗虫的消化道组成;口→咽→食道→嗉囊→砂囊→胃→回肠→结肠→直肠→肛门•棘皮动物完整的消化管,但肛门通常不用。
如海盘车的消化道组成为;口→食道→贲门胃→幽门胃→肠→肛门↓↓幽门盲囊肠盲囊六、呼吸系统•原生动物至原体腔动物由体表进行气体交换。
•环节动物一般用体表进行气体交换,有的出现特化的辅助呼吸结构。
•软体动物出现鳃和肺(假肺)※本鳃由外套膜内壁拓展形成的具有纤毛和丰富血管的呼吸结构。
如河蚌本鳃呼吸时的水流;入水管→外套腔→鳃小孔→鳃水管→鳃上腔→出水管※肺陆生软体动物外套膜内表面形成的呼吸结构。
※次生鳃(二次性鳃)腹足纲后鳃亚纲动物的本鳃退化后,由体表向外形成的膜状突起。
•节肢动物用鳃、肺、气管进行呼吸,是分类的重要依据。
鳃或书鳃:指水生节肢动物附肢基部的体壁向外突起形成的呼吸结构。
书肺:指陆生节肢动物由书鳃内陷后形成的呼吸结构。
气管:指陆生节肢动物体壁内陷形成的管道状呼吸结构。
甲壳纲:一般用鳃呼吸(虾、蟹),小型种类由体表呼吸(水蚤),陆生种类用伪气管(鼠妇)呼吸。
蛛形纲:书肺呼吸(蝎),书肺和气管呼吸(蜘蛛)。
昆虫纲:气管呼吸(蝗虫),有些水生昆虫的幼虫用气管鳃(蜻蜓、蜉蝣)呼吸。
•棘皮动物用体表皮鳃呼吸,管足也有辅助呼吸作用。
七、循环系统•原生动物无循环系统由原生质流动完成。
•腔肠动物、扁形动物无特异的器官,由原始的消化循环腔兼行。
•原体腔动物无特异的器官,原体腔兼行。
•环节动物闭管式循环(由于真体腔出现)但蛭纲真体腔退化,被葡萄状组织填充,行开管式循环。
•软体动物真体腔退化,行开管式血循环。
头足纲除外,行闭管式循环。
河蚌血循环途径:心室→动脉→血窦→静脉→心耳→心室。
•节肢动物真体腔退化,行开管式循环。
蝗虫的血液循环图示;•混合体腔(血腔)被2个纵隔分隔为背部的围心窦、围脏窦和围神经窦,隔上有孔隙,使三个腔彼此相通。
心脏位于背血窦中,由8个心室组成,每个心室两侧具有心孔,血液后行经腹血窦及围脏窦隔膜上的孔进入背血窦,由心孔返回心室。
•棘皮动物循环系统退化,由体腔承担血循环的功能。
※围血系统由真体腔演化形成的管腔结构,是中轴器、环血管、辅血管包绕原体腔所形成的血窦。
类似于其他动物的血窦作用,无血循环功能。
八、排泄系统•原生动物至腔肠动物无特异的排泄器官,由体表完成排泄。
草履虫的伸缩泡显示•扁形、原体腔动物具有原肾管,为水调节器,有学者认为可以将代谢废物排出体外。
原肾管由外胚层沿身体两侧内陷形成的网状多分支的管道系统,它由一对纵行的排泄管及其许多分支的小管及末端的焰细胞组成的盲管。
•环节动物后肾管排泄。
后肾管中胚层起源的体腔膜形成的具有两端开口盘曲的体腔导管,一端位于体腔的漏斗状开口称为肾口;另一端称肾孔开口于体外。
环毛蚓在每体节中有数百个小肾管;包括三类:即体壁小肾管、咽头小肾管和隔膜小肾管。
后肾与原肾的区别:(1)两端开口,原肾为盲管。
(2)起源与原肾不同。
•软体动物由后肾管演化的肾脏。