光照强度对浮游植物丰度及吸收铁的影响
湖泊水深1m的光照强度

湖泊水深1m的光照强度
湖泊水深1m的光照强度是一个复杂而有趣的议题。
光照强度,通常表示为每平方米的光能量,是湖泊生态系统中至关重要的参数之一。
它不仅影响着湖泊中生物的光合作用和生长,还与水体的透明度和水质密切相关。
在湖泊水深1m处,光照强度受到多种因素的影响。
首先,太阳的角度和高度对光照强度起着决定性作用。
在正午时分,太阳直射水面,光照强度达到最大值。
然而,随着太阳角度的变化,光照强度也会逐渐减弱。
其次,水体的透明度对光照强度产生重要影响。
清澈透明的水体允许更多的阳光穿透,从而增加水深1m处的光照强度。
相反,浑浊的水体会吸收和散射光线,导致光照强度降低。
此外,湖泊中的浮游生物、溶解物质和底质也会对光照强度产生影响。
浮游生物如藻类和浮游植物会吸收光能进行光合作用,从而减少到达水深1m处的光照强度。
溶解物质如有机物和矿物质颗粒也会散射和吸收光线,进一步影响光照强度。
底质的颜色和反射性也会对光照强度产生一定的影响。
湖泊水深1m的光照强度对于湖泊生态系统具有重要意义。
它直接影响着水生植物的光合作用效率和生长速度,进而影响整个生态系统的物质循环和能量流动。
同时,光照强度还对湖泊中的浮游生物群落结构和分布产生重要影响,从而改变水体的透明度和水质。
综上所述,湖泊水深1m的光照强度是一个综合多种因素的复杂指标。
它受到太阳角度、水体透明度、浮游生物、溶解物质和底质等多种因素的影响,并对湖泊生态系统产生重要影响。
因此,对于湖泊管理和生态保护而言,了解和监测湖泊水深1m的光照强度至关重要。
光照强度的变化对植物的影响规律和应用

当超过一定光强,光合速率增加就会 转慢,当达到某一光强时,光合速率 就不再增加,而呈现光饱和现象。开 始达到光合速率最大值时的光强称为 光饱和点此点以后的阶段称饱和阶段
前一阶段中主要是光强制约着光合速 率,而饱和阶段中CO2是主要限制因 素。
比例阶段
三)大多数植物能在明亮的阳光下发育的 很好也能忍受一定程度的荫蔽,他们叫 耐阴植物。
这不携补洋所消用的大植在在光
些能带偿中谓耗量下于物海海的
藻很到点的的相刚部呼的洋洋穿
的性
会升点或藻点之植物。作的分限
死到以者类。处物的在用透布制
亡表下被沉如,的光透量光,着
。层而洋降果就呼合光才带只植
时又流到海是吸作带能内有物
,
,
光强的影响机理
光照强度过强时,甚至会破坏原生质,引 起叶绿素分解,或使细胞失水过多而导致 气孔关闭,造成光合作用的减弱或者停止。 光照度过弱时,植物光合作用制造的有机 物质比呼吸作用消耗的还少,这是植物就 会停止生长,甚至死亡。
光照强度对植物的产量,质量的影响
饱和阶段
培育作物新品种时,要求作物品种所需的光补 偿点越低,光饱和点越高,越能够充分利用光 能,获得高产。
强光伤害——光抑制
光能不足可成为光合作用的限制因素,光能过剩也会对光合 作用产生不利的影响。当光合机构接受的光能超过它所能利 用的量时,光会引起光合速率的降低,这个现象就叫光合作 用的光抑制
光照强度与植物生长的关系
➢ 原理 植物必须生产足够的糖类以弥补呼吸
消耗,接生长 受一定量的光照是植物获得净生发育 产量的 必要条件。
➢ 影响 ⑴使植物体内的各种器官和组织保持发
育上的正常比例 ⑵使植物体重量的增加,使植物生
海洋浮游植物的光合作用机制

海洋浮游植物的光合作用机制一、引言海洋是地球上最大的生态系统之一,其中光合作用是维持海洋生物生存的重要过程。
海洋浮游植物是海洋生态系统中的重要组成部分,它们依靠光合作用将太阳能转化为有机物,为整个生态系统的持续运转提供能量和营养物质。
二、光合作用的基本原理光合作用是指光合有机物和光合氧的生成过程。
在光照条件下,浮游植物利用叶绿素等色素吸收光能,将光能转化为化学能,并利用水和二氧化碳进行反应,产生有机物质和氧气。
光合作用可以用化学方程式表示如下:光照能量+ 6CO2 + 6H2O → C6H12O6 + 6O2从上述方程式可以看出,光合作用需要光能、二氧化碳和水的参与。
在海洋环境中,由于二氧化碳和水的丰富,光是限制海洋浮游植物光合作用的主要因素。
三、光合作用的机制1. 光能的吸收与传递海洋浮游植物中的光能吸收主要由叶绿素等光合色素完成。
这些色素具有吸收不同波长的光能的能力,可以利用可见光的大部分区域进行光合作用。
其中,叶绿素a是光合作用最主要的色素之一,它主要吸收红、橙和蓝紫色光线。
一旦光能被吸收,它将在光合色素的载体蛋白中传递,并逐渐转化为化学能。
2. 光能转化的化学过程光能的转化是通过光合作用中的光化学反应完成的。
光化学反应包括光能与光合色素的相互作用、释放电子和形成能量等一系列的过程。
当光能传递到叶绿素分子中时,光合色素分子中的电子被激发,并且从低能级跃升到高能级。
这个过程中,光合色素分子变得富余电子,称为激发态。
随后,激发态的光合色素会释放出电子,并将其传递给其他化学物质,如细胞色素等。
这个过程称为光化学反应的能量传递。
同时,由于光合色素分子释放电子,它本身也需要补充电子。
这个电子供给过程是通过水分子分解而来的。
在光合作用的初期,光合色素将电子从水分子中取出,形成氧气和氢离子。
氢离子被用于合成光合有机物,而氧气则释放到周围水体中。
3. 二氧化碳的固定浮游植物通过气体交换从海水中吸收二氧化碳,并将其固定为光合有机物。
海洋食物网的关键环节浮游植物与浮游动物的相互关系

海洋食物网的关键环节浮游植物与浮游动物的相互关系海洋食物网的关键环节:浮游植物与浮游动物的相互关系海洋食物网是指海洋生物之间通过摄食和被摄食的关系形成的复杂网络系统。
其中,浮游植物和浮游动物是海洋食物网的重要组成部分,它们之间的相互关系对整个海洋生态系统的稳定与发展起着至关重要的作用。
一、浮游植物的重要性浮游植物是海洋中最主要的初级生产者,通过光合作用将阳光能转化为有机物质。
浮游植物主要包括微型浮游藻类和细菌等。
它们在水体中形成大量的浮游植物群落,为其他海洋生物提供丰富的有机物质,为海洋生态系统的物质循环提供能量来源。
浮游植物的生长和繁殖受到光照、水温、营养盐和微量元素等因素的影响。
在海洋中,浮游植物的分布呈现明显的垂直和水平分布特征。
在上层水域,由于光照充足,浮游植物生长旺盛;而在深层水域,受限于光照,浮游植物数量较少。
此外,浮游植物的季节性变化也对整个海洋食物网的结构和演化产生重要影响。
二、浮游动物的重要性浮游动物是海洋食物网中的二级消费者,它们以浮游植物为食,同时也成为其他海洋生物的食物来源。
浮游动物包括浮游性甲壳类动物、浮游性腔肠动物等。
它们通过摄取浮游植物和其他有机物质来获取能量和营养,同时将浮游植物的能量传递给更高级别的生物。
浮游动物的丰富度和群落结构受到浮游植物的生长和分布状况的影响。
一方面,浮游植物的数量和品种丰富程度直接影响浮游动物的数量和群落结构;另一方面,浮游动物会对浮游植物进行摄食,从而维持浮游植物的种群数量和结构平衡。
三、浮游植物与浮游动物的相互关系浮游植物与浮游动物之间存在着密切的相互作用关系。
一方面,浮游植物为浮游动物提供了重要的食物资源,能够满足其能量和营养需求。
浮游动物通过摄食浮游植物,将植物的有机物质转化为自身的能量和营养物质。
另一方面,浮游动物通过摄食浮游植物控制了浮游植物的生长和数量。
当浮游植物过度繁殖时,浮游动物的摄食作用可以减少浮游植物数量,维持水体中的生态平衡。
流水槽不同区域溶氧昼夜变化测定试验
2018.7池塘工业化养殖是生态养殖的典型模式之一,具有增产增效、环境友好、质量安全、管理便捷等优点,已作为江苏省海洋与渔业局重大科技推广项目。
当前,盐城市建湖县已建成全国面积最大的池塘循环水生态养殖基地,走在全国同类养殖模式前列。
笔者曾经在基地亲身蹲点数月,参与一线的养殖过程,开展了相关试验。
本次对流水槽不同区域溶氧昼夜数据的测定,旨在追踪流水槽养殖过程中溶解氧动态变化的全程信息,为今后的系统研究积累原始资料。
一、材料与方法1.试验时间试验开展于2016年8月19日,时间选择在夏季,因为夏季溶解氧的变化尤其是垂直变化更为明显,且高温季节的溶解氧含量通常作为渔业生产最重要的限制性因子之一。
2.试验地点试验地点位于盐城建湖县的池塘循环水养殖基地,彼时已建成52条流水槽,每条流水槽长22米×宽5米×深2米,规模为全国之最。
本次试验所选择的流水槽养殖鳊鱼苗约12万尾,苗种平均规格为250尾/千克。
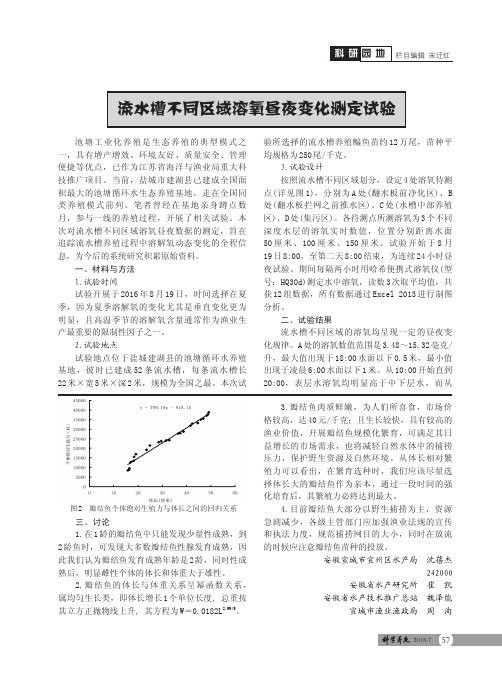
3.试验设计按照流水槽不同区域划分,设定4处溶氧待测点(详见图1),分别为A 处(翻水板前净化区)、B 处(翻水板拦网之前推水区)、C 处(水槽中部养殖区)、D 处(集污区)。
各待测点所测溶氧为3个不同深度水层的溶氧实时数值,位置分别距离水面50厘米、100厘米、150厘米。
试验开始于8月19日8:00,至第二天8:00结束,为连续24小时昼夜试验。
期间每隔两小时用哈希便携式溶氧仪(型号:HQ30d)测定水中溶氧,读数3次取平均值,共获12组数据,所有数据通过Excel 2013进行制图分析。
二、试验结果流水槽不同区域的溶氧均呈现一定的昼夜变化规律。
A 处的溶氧数值范围是3.48~15.32毫克/升,最大值出现于18:00水面以下0.5米,最小值出现于凌晨6:00水面以下1米。
从10:00开始直到20:00,表层水溶氧均明显高于中下层水,而从三、讨论1.在1龄的瓣结鱼中只能发现少量性成熟,到2龄鱼时,可发现大多数瓣结鱼性腺发育成熟,因此我们认为瓣结鱼发育成熟年龄是2龄,同时性成熟后,明显雌性个体的体长和体重大于雄性。
海洋生态系统中的海洋光照变化响应研究

海洋生态系统中的海洋光照变化响应研究海洋,占据了地球表面约 71%的面积,是一个神秘而又充满生机的世界。
在这个广袤的生态系统中,海洋光照的变化对其中的生物和环境产生着深远的影响。
海洋光照作为一个重要的环境因子,不仅影响着海洋生物的生长、繁殖和分布,还对海洋生态系统的结构和功能起着关键的调控作用。
海洋中的光照强度和光谱分布会随着深度、季节、地理位置以及天气等因素而发生变化。
在浅海区域,阳光可以相对充足地穿透水面,为海洋植物和浮游生物提供了丰富的光能。
而在深海,光线逐渐减弱,直至完全黑暗。
这种光照强度的垂直梯度变化,导致了海洋生物在垂直分布上呈现出明显的分层现象。
例如,浮游植物通常集中在海洋表层,因为这里光照充足,有利于它们进行光合作用。
而一些深海生物则进化出了特殊的适应机制,以在微弱光照甚至无光的环境中生存。
海洋光照的变化还会影响海洋生物的生理过程。
对于光合作用的生物来说,光照是其能量的主要来源。
光照强度的增加或减少,会直接影响光合作用的速率,从而影响生物的生长和繁殖。
当光照强度不足时,浮游植物的光合作用受到抑制,可能会导致其生长缓慢、数量减少,进而影响整个食物链的基础。
另一方面,光照强度过高也可能对生物造成伤害,例如导致光合作用系统的损伤和氧化应激反应。
海洋中的生物还会根据光照的变化调整其行为模式。
许多海洋动物具有明显的昼夜节律,它们的觅食、迁徙和繁殖等行为都与光照周期密切相关。
例如,一些鱼类和虾类在白天会躲在暗处,夜晚则出来活动觅食。
此外,海洋生物的体色和形态也可能会因光照环境的变化而发生适应性改变。
一些海洋生物为了躲避天敌或更好地捕食,会在不同的光照条件下改变自身的颜色和形态,以达到伪装或警示的目的。
除了对生物个体的影响,海洋光照的变化还会对海洋生态系统的结构和功能产生宏观层面的影响。
在全球气候变化的背景下,海洋温度升高、海平面上升以及海冰融化等现象,都可能导致海洋光照条件的改变。
这些变化可能会影响海洋生态系统中生产者、消费者和分解者之间的平衡,进而影响物质循环和能量流动。
海洋生态学实验:实验三、光照强度与浮游植物光合作用速率的关系

三、仪器与设备:
实验三、光照强度与浮游植物光合作用速率的关系
一、实验目的:
1、了解光照强度与浮游植物光合作用(产氧量)的关系, 掌握用黑白瓶氧含量与光合作用熵计算总产量与净产量。
二、原理:
1、光强与浮游植物光合作用速率的关系:在低光照 条件下,光合作用速率与光强成正比,随光强增加,光合 作用速率逐渐达最大值,这时光强称为饱和光强(IK) ,光 强继续增加,光合作用因光照过度而受抑制。不同种类的 饱和光强值不同。因此,浮游植物在适宜光强范围内,光 照越强,光合作用速率越快,初级生产力越高。
室内光照
(填写光强)
培养箱光照1
(
)
培养箱光照1
(
)
培养箱光照2
(
)
起始氧 含量
(mgO2/L· h)
白瓶氧含 量
(mgO2/L· h)来自黑瓶氧含 量(mgO2/L· h)
总生产量 (mgC/L·h)
净生产量 (mgC/L·h)
七、思考题:
试分析黑白瓶测氧法的优缺点及主要影响因素。
2、光合作用速率的测定(黑白瓶测氧法): 浮游植物光合作用可用下式简单表示:
CO2+H2O→(CH2O)+O2 在一定时间内,浮游植物在光合作用过程中吸收的CO2、 释放的O2和生成的有机物之间存在一定比例,因此可由产 氧量间接估算光合作用产量。
光合作用熵(PQ):指光合作用释放的O2分子数与所还原 CO2分子数的比值。光合作用产物不同,PQ值不同,合成 碳水化合物的PQ值为1,脂类为1.4,以NH4-N为氮源合成 蛋白质时, PQ值为1.05,以NO3-N为氮源则为1.6。PQ平 均值为1.25。由此可得下式: P(mg·C/L·h)= 3/8× O2(mg/L·h)×1/PQ。
光照对我国常见藻类的影响机制及其应用

光照对我国常见藻类的影响机制及其应用光照对藻类的影响机制1.光照强度对藻类生长的影响表1 我国常见藻类最适光强/最适光强范围2.光照强度对藻类生理活动的影响除直接影响藻类生长外,光照强度对藻类光合活性、放氧速率、酶活性、胞内色素组成、脂肪和碳水化合物含量、新陈代谢产物等都会产生影响。
刘世明等的研究结果表明:在14,16,18 ℃的温度梯度和100,250,360 μmol/(m2·s)的光照强度梯度下,铜绿微囊藻在18 ℃、100 μmol/(m2·s)下生长速率和光合活性都显著高于其他组。
李娜等的研究结果表明:光照强度为0.9~252 μmol/(m2·s)时,刚毛藻的净产氧量和毛产氧量与光照呈曲线相关,光照为103.5 μmol/(m2·s)时刚毛藻光合作用的产氧量最高。
卿人韦等发现:极大螺旋藻的过氧化物歧化酶SOD活性在不同高光照强度胁迫下,呈先增加后下降趋势,但活性值始终高于正常光照;过氧化氢酶CAT活性则呈先下降后上升的变化,活性值始终低于正常光照。
当受到光限制时,藻类能够通过增加光合单位(photosythetic units)的数量及光捕获色素蛋白复合体的大小来增加细胞的色素;当藻类受到强光抑制时,捕光色素可能会被分解,光保护色素的含量则会相对增加。
Foy 等对颤藻的研究发现:低光照会导致藻青蛋白和叶绿素a含量的增加,但藻青蛋白的上升比例要高于叶绿素a,藻青蛋白与叶绿素a含量的比例与藻类对光的捕获效率呈显著正相关。
低光强条件下,藻类类囊体膜的表面积及其上色素蛋白复合体的数量均会增加,脂肪含量较高,碳水化合物含量相对较低;高光条件下,光合色素含量减少,膜脂合成速率降低,碳水化合物合成能力增加,进而导致脂肪含量较低,碳水化合物含量升高。
石娟等研究了小新月菱形藻(MACC/B228)和等鞭金藻(MACC/H060)在不同光强下的脂肪含量和碳水化合物含量,结果表明:2种藻均在低光下脂肪含量多,碳水化合物含量少。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
东
北
Hale Waihona Puke 林业大学
学
报
V o 1 . 4 3 No . 1 J a n .2 0 1 5
J OUR NAL OF NOR T HE AS T F ORE S T RY UNI VE RS I T Y
光 照 强 度 对 浮 游 植 物 丰 度 及 吸 收 铁 的 影 响 )
强为 1 2 0 0 0l x时 , 吸收 F e量为 0 . 0 9 1 m g・ L 的培养瓶 中浮游植 物丰度增加 了 2 7 9 1 . 2 i n d・ L ~, 说 明了光照 强度对 藻 类生长的限制作用明显 高于培养液 中营养元素对藻类生长的限制作 用。 关 键 词 光 照 强度 : 浮游植物 ; 丰度 ; 吸收铁 ; 小 兴 凯 湖 分 类 号 Q1 7
L i n 0 o f ph y t o pl a nk t o n Fe u pt a k e.I n 8 0 00 l x.t he p h y t o p l a n k t o n a b un d a nc e wa s p o s i t i v e l y c o r r e l a t e d wi t h Fe up t ak e .I n
1 2 0 0 0l x. t h e p h y t o p l a n k t o n a b u n d a n c e i n c r e a s e d b y 27 91 . 2 i n d / L i n 0 . 0 9 l mg /L o f F e u p t a k e . T h e l i g h t i n t e n s i t y o n t h e r e s t ic r t i o n o f a l g a e g r o wt h i n n u t ie r n t s o l u t i o n c u l t u r e wa s mo r e o b v i o u s i n t h e a l g a e g r o th w r e s t i r c t i o n .
E f r e c t 0 f L i g I l t I n t e n s i t y o n t h e Ab s o r p t i o n o f I r o n a n d t h e P h y t o p l a n k t o n Ab u n d a n c e / / Y u H o n g x i a n , Z h a n g Y i n g y — i n g ,Ma C h e n g x u e , J u Y o n g f u ( N o r t h e a s t F o r e s t r y U n i v e r s i t y , Ha r b i n 1 5 0 0 0 0 , P . R . C h i n a ) / / J o u ma l o f N o r t h e a s t F o r e s t - r y U n i v e r s i t y , 2 0 1 5 , 4 3 ( 1 ) : 9 3 - 9 5 .
于 洪 贤 张 莹 莹 马 成 学 鞠 永 富
( 东北林业大学 , 哈尔滨 , 1 5 0 0 4 0 )
摘 要 在小兴凯湖采集水样 , 滤掉浮游动物 , 对浮游植物进行为期 1 2 d的 实验 室培 养。试验设 置 了4个光 照 梯 度 和 5个 F e浓 度 , 分 别 进 行 对 比试 验 。结 果 表 明 : 随着 F e浓 度 的 增 加 , 浮游植物 对 F e的 吸 收 量 也 相 应 的 增 加 。在低光 照强度下 , 浮游植物 对 F e的吸收规律 不明显。在光 强为 4 0 0 0l x时, 浮游植物吸收 F e 量 为 0的培养瓶 中 浮游 植 物 丰度 降低 了 6 3 . 6 i n d・ L ~。 在 光 强 为 8 0 0 0 l x条 件 下 , 浮游植物丰度与吸收 F e量 呈 正相 关 关 系。 在 光
s o r p t i o n a mo u nt o f Fe h a d 1 3 0 r e gu l a r l a w.I n 4 0 00 l x o f l i g ht i nt e ns i t y,t he p hy t o p l a nk t o n a b u nd a n ce r e d uc e d b y 6 3. 6 i n d /
We c o l l e c t e d w a r e r s a mp l e s .f i l t e r e d p l a n k t o n i c a n i aa t 1 .c u l t u r e d t h e p h y t o p l a n k t o n i n t h e S ma l l Xi n g k a i L a k e f o r 1 2 d i n t h e l a b o r a t o r y . We s e t f o u r l i g h t g r a d i e n t s a n d f i v e d i f f e r e n t F e c o n c e n t r a t i o n s b y c o mp a r a t i v e e x p e r i me n t s .W i t h t h e i n c r e a s e o f F e c o n c e n t r a t i o n.a b s o r p t i o n o f p h y t o p l a n k t o n o n F e i n c r e a s e d c o r r e s p o n d i n g l y .I n l o w l i g h t i n t e n s i t y .t h e a b —