分子遗传学5章叶绿体和线粒体的基因组及其表达
叶绿体基因的遗传方式

叶绿体基因的遗传方式
1.自主性遗传:叶绿体内含有自己的DNA,被称为叶绿体DNA 或cpDNA,它能够进行自我复制和表达。
这意味着叶绿体中的一些基因可以独立于核基因组进行复制和传递给下一代。
2.母系遗传为主:在大多数植物中,受精卵中的细胞质几乎全部来自于卵细胞,因此,叶绿体及其携带的基因基本上通过雌性生殖细胞单向地传给后代,遵循的是细胞质遗传规律,而非孟德尔定律所描述的核基因遗传规律。
3.细胞分裂时的不均等分配:在有丝分裂和减数分裂过程中,尽管叶绿体会随着细胞质的分裂而被传递到子细胞中,但这种分配往往不是完全均等的,导致不同子细胞中叶绿体的数量和类型可能有所差异。
4.核基因调控:虽然叶绿体有自己的基因组,但是其蛋白质合成并不完全自主,很多与叶绿体功能相关的基因实际上是存在于细胞核内的,这些核基因编码的产物对叶绿体的功能和发育起着关键作用。
也就是说,虽然叶绿体基因直接参与某些过程,但它们的表达和功能还受到细胞核内基因的控制。
植物线粒体基因组结构及其演化分析

植物线粒体基因组结构及其演化分析1、植物线粒体基因组结构概述线粒体是细胞内负责进行能量代谢和氧化呼吸的重要器官。
作为一个独立的细胞器,线粒体被覆盖着一个双层膜,并具有自己的基因组和细胞内核的DNA。
植物线粒体基因组包含在线粒体内部的循环DNA中,通常包含有37-102个基因,其中约60-70%编码的是线粒体酶和蛋白质,剩余的编码的是tRNA和rRNA。
当我们对植物线粒体基因组进行结构分析时,可以发现该基因组由两段的DNA组成:环状线粒体DNA和线性DNA。
环状线粒体DNA是由单个双链环状DNA组成的,通常长度为100-250 kb;线性DNA是由多个线性DNA片段组成的,通常以长度为3-10 kb的终止子序列(telomeric sequence)为末端。
2、植物线粒体基因组演化分析植物线粒体基因组的演化可以分为三个阶段:线性DNA化阶段、基因缩减和基因拼凑阶段。
在线性DNA化阶段,植物线粒体基因组从环状基因组转变为线性基因组,这个过程存在两种不同的机制:复制缺失和复制折断。
这两种机制都可导致线性片段的产生。
在基因缩减阶段,基因组中的大量基因丢失。
这些基因编码了线粒体的功能蛋白和其他类型的分子。
在研究中,发现集一些线粒体基因在种间高度可变,但一些重要的线粒体基因在植物具有高度保守性。
例如,ATP酶亚基和呼吸链的一部分是高度保守的。
这表明植物线粒体基因组的保守性与其重要性有关。
在基因拼凑阶段,一些基因片段扮演了减少线粒体基因组大小的角色。
这些基因片段产生于逆转录过程,并在植物线粒体基因组中扮演了拼凑器(joiner)的角色,使得其他线粒体基因片断能够在大量的序列缺失的情况下被组装起来。
3、总结植物线粒体基因组是进行线粒体生物学研究中极其重要的物质基础,而其结构及演化分析则是进行这一领域研究的前提和核心内容。
通过对植物线粒体基因组的结构和演化分析,我们可以更好地了解线粒体的基本生物学过程,帮助我们更好地研究如何利用植物线粒体基因组改良作物。
分子生物学课件:线粒体医学

线粒体基因组的组成和结构
线粒体基因组由13个线粒体蛋白编码基因、22个线粒体 tRNA基因和2个线粒体rRNA基因组成,它们共同构成了一个 闭合环状的线粒体DNA(mtDNA)。
mtDNA的长度约为16.5kb,是哺乳动物细胞中唯一一个非 染色体DNA,其结构包括一个长柄(D-loop区)和两个短柄 (N-和C-端),其中D-loop区是mtDNA的转录和复制起点 。
线粒体拥有自身的遗传物质和遗传体系,但其基因组大小有限,是一种半自主细 胞器。
线粒体医学的发展历程
线粒体医学的发展历程可以追 溯到20世纪60年代,当时科学 家们开始研究线粒体在疾病中 的作用。
20世纪80年代,线粒体遗传学 开始崭露头角,对线粒体疾病 的研究逐渐深入。
近年来,随着线粒体生物学和 医学研究的快速发展,线粒体 医学逐渐成为研究的热点。
01
深入探究线粒体功能 与疾病的关系
通过高通量测序、生物信息学分析等 方法,深入研究线粒体基因突变与疾 病发生发展的关系,揭示线粒体在疾 病中的作用和机制。
02
推进跨学科合作与交 流
加强分子生物学、细胞生物学、病理 学、神经科学等学科之间的合作与交 流,共同推动线粒体医学的发展。
03
加强临床应用研究
通过开展临床试验,探究针对线粒体 的靶向性治疗策略在疾病治疗中的应 用效果,推动线粒体医学与临床医学 的深度融合。
06
相关案例分享
线粒体基因组研究与帕金森病诊断的案例分享
总结词: 线粒体基因组研究有助于解析帕 金森病的病因,并提供诊断依据。
• 线粒体基因组研究还可用于诊断疑似 病例,并指导治疗方案。
通过开发特定的药物,可 以针对线粒体进行治疗, 改善线粒体功能,缓解病 情。
遗传学(第3版)第5章核外遗传分析

sm-r mt + sm-s mt - 全部后代(99%)表现为sm-r sm-s mt + sm-r mt - 全部后代(99%)表现为sm-s
以上结果表明正反交结果不同,说明衣藻对链霉素的抗性或 敏感是由非孟德尔遗传因子所控制(图5-3)
研究表明:该性状由衣藻中叶绿体基
因组DNA上的基因所控制。交配型mt+ 和mt–细胞质中都含有叶绿体基因组 DNA,当发生交配时,两个亲本对合 子贡献的细胞质是相等的,而后代总 是保留mt+亲本与叶绿体基因组上控 制的性状有关的表型。一种分子解释
某些产物积累在卵母细胞的细胞质中,使子代表型不由自身 的基因型所决定而出现与母体表型相同的遗传现象,则称为 母体影响。 母体影响有两种: 一种是短暂的,只影响子代个体的幼龄期;
5.1.3衣藻抗生素抗性的遗传
衣藻通常行无性生殖,有时通过两种形态相同但交配型
不同的配子进行融合,行有性生殖。同型配子间不能融合,
仅相反交配型的配子能够融合产生二倍体的合子。衣藻的交 配型是由细胞核内一对等位基因mt +和mt -所决定的。配子
融合给合子提供了相等的细胞内含物,合子萌发时,通常立
即发生减数分裂,四个减数分裂的产物作为非顺序四分子在 一个子囊中,其中有两个mt +细胞和两个mt -细胞,即核基
②自体受精(Autogamy): 同一个体的两个小核经减数分裂,留下一个小核, 该小核分裂一次又相互合并,随后再分裂发育成小核和大核。 (Fig--45 ) 3).草履虫放毒型和敏感型的接合 若 某些草履虫 草履虫素 杀死其它的草履虫但对自己无害。 能产生草履虫素的个体叫做放毒型 而受害的个体就称为敏感型 根据遗传学实验,草履虫的放毒型必需有两种因子同时存在: ① 细胞质因子,叫做卡巴粒(Kappa particle , κ)
叶绿体线粒体基因和核基因特点

叶绿体基因组的结构现在已知道,叶绿体DNA(ctDNA)是双链环状分子,大小约为120~160kb。
大多数植物的ctDNA分子中有两段反向重复的核苷酸序列,是大多数植物ctDNA 所共有的特征,大小约在6~76kb之间,不同植物ctDNA大小的差异主要是由于反向重复序列大小的不同而引起的。
但在有些植物如蚕豆、豌豆并未发现有两段对称的反向重复序列,而在眼虫藻等却发现有3段正向重复序列。
Shinozaki等于1986年分别发表了烟草和地钱的ctDNA全序列,Hiratsuka也于1989年报道了水稻ctDNA的全序列,这些均显示ctDNA含有编码叶绿体rRNA和tRNA的基因以及100个左右的蛋白质结构基因。
同时发现大多数植物的ctDNA含有2个rDNA 拷贝,眼虫藻中有3个紧邻的rDNA拷贝,豌豆和蚕豆却仅含有一个rDNA拷贝,所以一般认为rDNA存在于ctDNA反向重复序列中。
埃内亚斯(Eneas Filho)等发现豌豆叶绿体中核糖体的60种不同蛋白组分中,有1/3是由ctDNA编码。
现在已公认在叶绿体的所有蛋白质中既有叶绿体基因组编码的(约占总种类的30%,例如光合作用反应中心的蛋白等)也有核基因组编码的(约占总种类的70%,例如类囊体膜的一些蛋白等),还有一些功能性寡聚蛋白质,其组成亚基分别是由两个基因组编码的,例如叶绿体中的可溶性蛋白Rubisco,其大亚基是由自身基因组编码,而小亚基则由核基因组编码。
但也有例外,Reith等就曾在灰藻和红藻中发现Rubisco的大小亚基皆由叶绿体自身基因组编码,且大小亚基的基因共同组成一个操纵子。
Turmel等于1978年报道在衣藻ctDNA的23SrDNA中发现有一个内含子(intron)的存在,这是第一次在ctDNA中发现的与真核生物核基因类似的断裂基因。
后来,Koch等在玉米ctDNA的trnI和trnA基因中也发现有内含子。
高等植物叶绿体DNA的内含子主要在tRNA基因中发现,很少在蛋白质结构基因中发现,但在绿藻中却发现大量的蛋白质结构基因也存在有内含子。
分子遗传学5章叶绿体和线粒体的基因组及其表达

叶绿体基因组(cpDNA)及其表达
(一)叶绿体基因组结构
(二)叶绿体基因组特点
(1)环状双螺旋分子,不与蛋白质结合。 (2)基因组复制和遗传复杂且难以确定。 (3)基因组大小在120kb-160kb。含120-140个基因。 (4)某些基因含有内含子。 (5)有的基因组合可形成操纵子。 (6)所带基因主要控制tRNA、rRNA及约80种蛋白质合成。
表20-2 在线粒体中密码子的改变
生物 共同 哺乳动物 哺乳动物 果蝇 果蝇 纤毛虫 另一类纤毛虫 酵母 酵母 酵母 酵母
密码子 UG AUA CCA CUG UAG
MtDNA编码 Trp 终止 Met(起始) Met Ser Glu Cys Met(延伸) Thr Ser Ser
支原体
UGA
Trp
核密码 终止 Arg Ile Ile Arg 终止 终止 Ile Leu Leu 终止 终止
(三)线粒体基因的转录和翻译调控
转录与原核生物的转录相似;转录产物也往往没有完整的5’帽子和3’ poly(A)结构;转录后调控中RNA编辑和剪接占有重要地位;
线粒体核糖体沉降系数为60-78S,也与原核生物的核糖体类似。线 粒体基因存在和核基因的协调互作作用,
(四)线粒体基因相关的胞质雄性不育(CMS)
线粒体 细胞核
母系遗传 核遗传
细胞质雄性不育 育性恢复
玉米CMS
三系法 杂交制 种技术
线粒体的起源
• 内共生假说 • 1970,Margulis,真核
细胞祖先是种吞噬细胞; 线粒体祖先是种革兰氏 阴性菌。前者吞后者 • 细胞分化假说 • 原始的原核细胞质膜内 陷包被DNA,然后再分 化形成独立的细胞器。
线粒体基因组

线粒体基因组概述:线粒体基因组是细胞内的一种特殊类型的基因组,位于线粒体内。
它具有自主复制和自主转录的能力,与细胞核基因组相互作用,并在细胞内的许多重要生物过程中发挥关键作用。
本文将介绍线粒体基因组的组成、结构、功能以及与人类疾病的关联。
组成和结构:线粒体基因组是一个环形的DNA分子,其大小约为16.6 kb到18.7 kb之间。
它主要包含有37个基因,其中13个编码线粒体内膜的成分,22个编码线粒体的氧化磷酸化系统相关的成分,以及两个编码RNA转移酶。
线粒体基因组的特殊之处在于它是双膜结构的线粒体内的一部分,并独立于细胞核。
每个线粒体内都有多个复制的线粒体基因组存在,并且每个线粒体基因组都由自己的DNA复制和维护系统。
功能:线粒体基因组编码的蛋白质与线粒体的氧化磷酸化系统密切相关。
该系统是细胞内能量产生的主要途径,通过将氧化的有机物质转化为细胞可用的能量(ATP)。
线粒体细胞呼吸链中的五大复合物是由线粒体基因组编码的蛋白质组成的。
线粒体基因组还编码了一些与线粒体形态和功能调控相关的蛋白质。
例如,它编码了调控线粒体融合和分裂的蛋白质,以及调控线粒体对细胞应激的响应的蛋白质。
通过这些功能,线粒体基因组在细胞生理学和代谢调节中发挥着重要的作用。
与人类疾病的关联:线粒体基因组的突变或缺陷与许多人类疾病的发生和发展密切相关。
这些突变或缺陷可能导致线粒体功能受损,进而影响细胞的能量供应和其他重要生物过程。
许多遗传性线粒体疾病与线粒体基因组突变有关。
这些疾病包括线粒体脑肌病、线粒体呼吸链缺陷症、线粒体病等。
许多常见疾病,如癌症、神经退行性疾病和心血管疾病,也与线粒体基因组突变存在关联。
近年来,随着线粒体基因组测序技术的发展,更多的线粒体基因组突变与疾病的关联被发现。
这为诊断和治疗线粒体相关疾病提供了新的机会。
结论:线粒体基因组是一个独立于细胞核的环形DNA分子,具有自主复制和转录的能力。
它编码了与线粒体功能和生物过程密切相关的蛋白质,并在细胞能量供应和其他重要生物过程中发挥关键作用。
《线粒体与叶绿体》课件
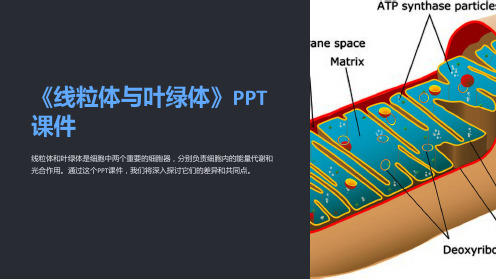
线粒体和叶绿体是细胞中两个重要的细胞器,分别负责细胞内的能量代谢和 光合作用。通过这个PPT课件,我们将深入探讨它们的差异和共同点。
概述
结构和功能
线粒体和叶绿体都是细胞内含 有膜结构的细胞器,分别在细 胞内进行不同的生化反应和代 谢过程。
ATP合成过程
线粒体将化学能转化为ATP,而 叶绿体通过光合作用产生ATP以 供细胞使用。
叶绿体DNA
独立存在的叶绿体DNA,与其它类型的DNA 一样在遗传和表达方面有各种特点和限制。
线粒体和叶绿体的区别
结构差异
线粒体和叶绿体在外形、膜结构 和内部的器官组织结构等方面有 很大区别。
功能差异
线粒体负责的是细胞内的能量代 谢,而叶绿体则为细胞提供光合 产物和氧气等物质。
遗传差异
虽然两者都有自己独立的DNA, 但是与此同时也有着各自特殊的 遗传方式和规律。
线粒体和叶绿体DNA
与核DNA不同,线粒体和叶绿 体都有自己独立的DNA,这个 特点对它们的功能和进化都有 着重要的意义。
线粒体
结构和功能
线粒体由外膜、内膜、基质和内嵌 膜(或内质网)等结构组成,完成 细胞内ATP的合成和代谢物的分解 等多个重要生化反应。
呼吸链与ATP酶
线粒体的呼吸链是其产生ATP所必 需的生化反应,在线粒体内部的 ATP酶则将化学能转化为能供细胞 使用的能量分子ATP。
线粒体和叶绿体的共同点
1 都有自己独立的DNA
2 都含有膜结构
3 都与能量代谢密切相关
线粒体和叶绿体都有自己的 DNA,这个特点对其进化和 生殖方式都有影响。
线粒体和叶绿体都含有膜系 统,会通过这个特殊的结构 进行不同的代谢反应和生化 变化。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
如何知道某一遗传性状为细胞质遗传?
可编辑ppt
3
叶绿体基因组(cpDNA)及其表达
(一)叶绿体基因组结构
可编辑ppt
4
(二)叶绿体基因组特点
(1)环状双螺旋分子,不与蛋白质结合。 (2)基因组复制和遗传复杂且难以确定。 (3)基因组大小在120kb-160kb。含120-140个基因。 (4)某些基因含有内含子。 (5)有的基因组合可形成操纵子。 (6)所带基因主要控制tRNA、rRNA及约80种蛋白质合成。
(4)植物线粒体基因存在混栖基因现象。
(5)线粒体基因的密码与核基因密码具有不通用性。 (6)所带基因主要控制tRNA、rRNA及某些蛋白质合成。
可编辑ppt
9
表20-2 在线粒体中密码子的改变
生物 共同 哺乳动物 哺乳动物 果蝇 果蝇 纤毛虫 另一类纤毛虫 酵母 酵母 酵母 酵母 支原体
密码子 UGA AGA/AGG AUA AUA AGA UAA/UAG UGA/UAG AUA CCA CUG UAG UGA
MtDNA编码 Trp 终止 Met(起始) Met Ser Glu Cys Met(延伸) Thr Ser Ser Trp
核密码 终止 Arg Ile Ile Arg 终止 终止 Ile Leu Leu 终止 终止
可编辑ppt
10
(三)线粒体基因的转录和翻译调控
转录与原核生物的转录相似;转录产物也往往没有完整的5’帽子和3’ poly(A)结构;转录后调控中RNA编辑和剪接占有重要地位;
可编辑ppt
6
紫茉莉花斑枝条的叶绿体细胞质遗传
表20-8 紫茉莉花斑植物杂交的结果
母本枝条表型 父本枝条表型 后代表型
白色
白色
白色
绿色
白色
花斑
白色
绿色
白色
绿色
绿色
绿色
花斑
绿色
花斑
白色
花斑,绿色,白色
绿色
花斑,绿色,白色
可编花 辑斑 ppt
花斑,绿色,白色 7
线粒体基因组(mtDNA)及其表达
(一)线粒体基因组结构
线粒体核糖体沉降系数为60-78S,也与原核生物的核糖体类似。线 粒体基因存在和核基因的协调互作作用,
可编辑ppt
11
(四)线粒体基因相关的胞质雄性不育(CMS)
线粒体 细胞核
母系遗传 核遗传
细胞质雄性不育 育性恢复
玉米CMS
三系法 杂交制 种技术
可编辑ppt
12
线粒体的起源
• 内共生假说 • 1970,Margulis,真核
可编辑ppt
8
(二)线粒体基因组特点
(1)一般认为是环状双螺旋分子,不与蛋白质结合。但物理结构复 杂。
(2)动物中和植物中线粒体基因组大小差别很大。动物15-18kb, 植物200-2500kb。且同一植物之间也存在较大差异,
(3)有些物种的线粒体中基因中含内含子(植物、酵母),而有的 物种则没有(人)。
细胞祖先是种吞噬细胞; 线粒体祖先是种革兰氏 阴性菌。前者吞后者 • 细胞分化假说 • 原始的原核细胞质膜内 陷包被DNA,然后再分 化形成独立的细胞器。
可编辑ppt
13
Maternal inheritance & Maternal effect
可编辑ppt
14
第五章 叶绿体和线粒体的基因 组及其表达
可编辑ppt
1
Chloroplast
Cell (plant)
Mitochondria
可编辑ppt
Байду номын сангаас
2
细胞核遗传与细胞质遗传
细胞核遗传:由细胞核中的DNA决定的遗传现象。 细胞质遗传:由细胞质中的DNA决定的遗传现象。
细胞质遗传特点:
(1)母系遗传 (2)杂交后代都不出现一定的分离比例(非孟得尔遗传)
可编辑ppt
5
(三)叶绿体基因的转录和翻译
(1)转录的mRNA无5’端帽子结构,3’通常无poly(A)。
(2)多顺反子转录,启动子基序与原核生物相似。
(3)转录后调控包括内含子的剪切(反式剪切现象),RNA编辑 等。 (4)所有叶绿体mRNA均由叶绿体中的核糖体负责翻译。
(6)叶绿体基因翻译产物往往和核基因翻译产物协同作用,完成其 生理功能。(Rubisco 全酶的组装)