糖的生物合成
糖的生物合成与代谢途径

糖的生物合成与代谢途径糖是生命中不可或缺的重要物质,它是生物体的主要能量来源之一,也是构成生物体的重要组成部分。
糖的生物合成与代谢是一系列复杂而精细的过程,它们通过一定的途径在细胞内进行。
在本文中,我们将探讨糖的生物合成与代谢的主要途径和相关机制。
第一节糖的生物合成糖的生物合成是细胞利用光能或化学能将无机物合成糖类化合物的过程。
主要的合成途径有光合作用和糖异生两种形式。
光合作用是指细胞通过叶绿体内的光化学反应,将二氧化碳和水转化为葡萄糖等有机物的过程。
在光照条件下,叶绿体中的叶绿素可以吸收太阳能,光合色素体可将太阳能转化为化学能,进而促使光合作用的进行。
光合作用分为光反应和暗反应两个阶段。
光反应发生在光合体系中,通过光合色素体捕捉光能,产生氧化还原电位,将光能转化为高能物质膜内的质子激励。
暗反应指的是光合作用中的还原和碳固定反应,主要在叶绿体基质内进行。
通过一系列酶的作用,将光反应所得的ATP和NADPH利用碳源还原为葡萄糖或其他有机物。
糖异生是指细胞在无光照条件下,通过有机物合成糖的过程。
糖异生主要发生在细胞质基质内,包括糖异生途径的两个重要过程:糖酵解和有机酸循环。
糖酵解是指将葡萄糖分解为丁醛酸,再将丁醛酸氧化为甲酸,最终合成糖的过程。
有机酸循环是指细胞质基质内的一系列反应,将葡萄糖分解为丙酮酸、柠檬酸等有机酸,最终通过一系列酶的作用合成糖。
第二节糖的代谢途径糖的代谢指的是细胞对糖化合物进行分解和利用的过程。
糖的代谢途径包括糖酵解、糖异生和糖氧化三个主要途径。
糖酵解是指细胞内部一系列酶的作用,将葡萄糖分解为丙酮酸或乙酸,产生ATP和还原能力分子NADH的过程。
糖酵解包括糖原糖酵解和异物糖酵解两种形式。
糖原糖酵解是指细胞内糖原被酵解,通过一系列的反应将糖原分解为葡萄糖,再进一步分解为丙酮酸,转化为乙酸最终释放能量。
异物糖酵解是指细胞利用外源性的碳水化合物,如蔗糖、木糖等进行糖酵解的过程。
糖异生是指细胞利用非糖类有机物合成糖的过程。
生物化学糖的生物合成1

(一)直链淀粉的生物合成-方式2
2.D-酶
D-酶是糖苷转移酶,作用于α-1,4糖苷键,用来合成引物。
D酶
+
麦芽三糖 给体
麦芽三糖 受体
++
麦芽五糖
葡萄糖
(一)直链淀粉的生物合成-方式3
3、淀粉合成酶 是淀粉合成的主要途径。
ADPG+引物 淀粉合成酶 淀粉+ADP
(二)支链淀粉的合成
1、淀粉合成酶: 只能催化形成α-1.4糖苷键, 合成直链淀粉。 2、Q酶(分支酶):既能催化α-1.4糖苷键的 断裂,又能催化α-1、6糖苷键的形成
注:支链淀粉降解时用的是脱支酶(R酶)
在Q酶作用下的支链淀粉的合成
2021
➢ 2. 两方面不同:
(1)糖异生必须克服糖酵解的三步不可逆反应。 (2)细胞定位:糖酵解在细胞液中进行,糖异生
则分别在线粒体和细胞液中进行。
糖酵解和糖异生的比较
三、糖异生作用的主要途径和关键反应
关键反应-迂回措施1
丙酮酸
CO2
ATP+H2O
ADP+Pi
丙酮酸羧化酶 (线粒体中) PEP羧激酶 (细胞质中)
F-1,6BP 活化 ATP 抑制
PEP
丙酮酸激酶
丙酮酸
PEP羧激酶 ADP 抑制
草酰乙酸
丙酮酸羧化酶
乙酰CoA 活化 ADP 抑制
7.2 蔗糖和多糖的生物合成
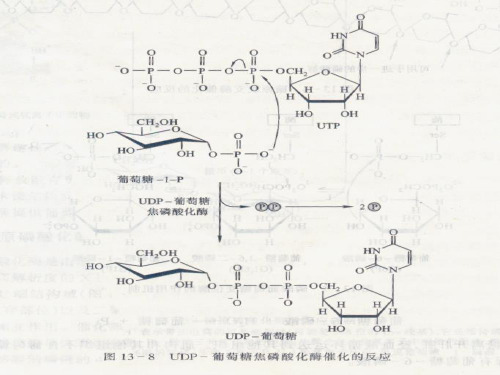
一、糖核苷酸的作用及形成 1.定义: 单糖与核苷酸通过磷酸酯键结合的化合物称为糖核苷酸。 2.作用:糖核苷酸是葡萄糖的活化形式与供体。 3.种类:目前发现的糖核苷酸主要有 UDPG,ADPG,TDPG,GDPG,CDPG等。在糖类代谢中,以 UDPG,ADPG为最重要。 4.形成:
糖的生物合成

②PEP羧激酶催化草酰乙酸形成PEP
PEP沿酵解途径逆向反应转变成1,6-二磷酸果 糖。
7.2.1.2 1,6-二磷酸果糖转化成6-磷酸果糖
二磷酸果糖酯酶是变构酶,受AMP、2,6-二磷 酸果糖变构抑制,但受ATP、柠檬酸变构激 活。
Rubisco是植物光合作用的关键酶, 为复合物,有8个大亚基(Mr53000), 每个含一个活性位点;8个小亚基 (Mr14000),功能还不完全清楚。
植物中酶位于叶绿体的基质,占叶绿 体总蛋白的50%,是生物圈中最丰富的 酶。
②羧化产物的还原
在激酶催化下3-PGA磷酸化生成1,3-二磷酸甘 油酸再被脱氢酶催化还原为3-磷酸甘油醛。
7.2.1.3 6-磷酸葡萄糖转化成葡萄糖
哺乳动物的糖异生作用在肝脏中进行;高等植 物主要发生在油料种子萌发时脂肪酸氧化产 物和甘油向糖的转变。
7.2.2 糖酵解和糖异生的互补调节
在细胞中糖异生作用和糖酵解作用相互协调、 受到很多代谢物的调控:
①高水平的ATP、NADH变构抑制磷酸果糖激 酶和丙酮酸激酶,而变构地激活二磷酸果糖 酯酶。
由于卡尔文在光合作用碳转化途径上作出了重 大贡献,于1961年获得诺贝尔奖。卡尔文循 环的最初产物为3-磷酸甘油酸,因此,此途 径也称C3-途径。
NADPH
的光
必反
需应
底 物
是 二 氧
产 生 的
ATP
化
碳 还
和
原
Calvin cycle可分为三个阶段。
①CO2的固定 在 核 酮 糖 -1,5- 二 磷 酸 羧 化 酶 (ribulose
高等植物叶绿体中含有叶绿素和类胡萝卜素两 类色素。
12糖类的生物合成

多分支,提高水 溶性,利于储存;
多处磷酸化酶作 用点,提高效率, 迅速提供能量
2、变位 G-1-P 磷酸葡萄糖变位酶
G-6-P
3、G的生成(只存在于肝、肾中,肌组织中没有此步骤)
G-6-
G
P
葡萄糖-6-磷酸酶
糖原的合成与分解代谢
UDP
Gn+1
Pi
Gn
糖原合酶 分支酶
UDPG
糖原磷酸化酶
Gn
• 酵解途径与糖异生途径多数反应是共有 的,除了酵解途径中3个不可逆反应。 糖异生方向
1.丙酮酸 磷酸烯醇式丙酮酸 2.1,6-双磷酸果糖 6-磷酸果糖 3.6-磷酸葡萄糖 葡萄糖
丙 酮
丙酮酸羧化酶
草 酰 磷酸烯醇式丙酮酸羧化激酶
酸
乙
CO2 ATP ADP+Pi酸 GTP GDP CO2
磷酸 烯醇 式丙 酮酸
3.调节酸碱平衡
长期饥饿时,肾糖异生增加,排氢保钠。 有利于维持酸碱平衡。
谷氨酰胺
谷氨酸
H2O
NH3
+
原尿H+
NH4+
-酮戊二酸 NH3
糖异生
糖异生的调节
1.激素的调节作用
6-磷酸果糖和1,6-双磷酸果糖之间,糖酵解和糖异 生之间
肝脏:胰高血糖素 腺苷酸环化酶
(无活性) 腺苷酸环化酶(活性)
ATP
2 胰高血糖素,主要的升血糖激素。
3 糖皮质激素,可引起血糖升高,肝糖原增加。
4 肾上腺素,强有力的升血糖激素。
• 胰岛素:降低血糖的唯一激素 1、促进G转运入细胞,加强利用 2、增强磷酸二酯酶的活性,降低cAMP水平,
激活糖原合酶,抑制磷酸化酶
第十七章:糖原的分解和生物合成(1)

葡糖-6-磷酸 葡糖 磷酸
CO2+H2 O 核糖
戊糖磷 酸途径 戊糖磷酸 磷酸丙糖 丙酮酸
酵解
乳酸、 乳酸、乙醇 发酵
糖异生 生糖氨基酸
乙酰辅酶A 乙酰辅酶
三羧酸循环 乙醛酸循环
ATP CO2+H2 O
重点
本章回顾及小结: 本章回顾及小结:
糖原的分解过程(掌握三种酶)、 糖原的合成过程(三个步骤三种 酶)及其调节机制。
3、磷酸葡萄糖变位酶 、
1-磷酸葡萄糖需要转变为6-磷酸才能进入代谢 磷酸葡萄糖需要转变为6 主流。催化磷酸基团转移的酶称为磷酸葡萄糖 主流。催化磷酸基团转移的酶称为磷酸葡萄糖 变位酶。1,6-二磷酸葡萄糖转变为6 变位酶。1,6-二磷酸葡萄糖转变为6-磷酸葡萄 磷酸葡萄糖变位酶又恢复原来的形式。 糖,磷酸葡萄糖变位酶又恢复原来的形式。此 催化机理与3 磷酸甘油酸变为2 催化机理与3-磷酸甘油酸变为2-磷酸甘油酸的 机理很相似。 机理很相似。 磷酸葡萄糖变位酶发挥催化活性需要少量的 1,6-二磷酸葡萄糖的存在,1,61,6-二磷酸葡萄糖的存在,1,6-二磷酸葡萄糖 由1-磷酸葡萄糖在磷酸葡萄糖激酶的催化下形 如果1,6 1,6成。如果1,6-二磷酸葡萄糖从磷酸葡萄糖变位 酶分子上脱落,酶的活性就会钝化。 酶分子上脱落,酶的活性就会钝化。
3. 作为机体组织细胞的组成成分
是糖蛋白、蛋白聚糖、糖脂等的组成成分。 是糖蛋白、蛋白聚糖、糖脂等的组成成分。 4. 分子识别作用 血型物质A和 、粘附分子、整合素、 血型物质 和B、粘附分子、整合素、细菌 的各种凝集素等
二、糖类合成
A.植物的光合作用 A.植物的光合作用 在植物叶绿体中,在光能驱动下 在植物叶绿体中,在光能驱动下CO2与H2O合成葡萄 合成葡萄 放出氧气的过程。 糖,放出氧气的过程。 B.动物的糖异生 B.动物的糖异生 异生——非糖物质合成糖原。 非糖物质合成糖原。 异生 非糖物质合成糖原 部位: 部位:肝脏 a.过程 过程
09糖的生物合成

3-P-甘油醛
A
A G-6-P磷酸酯酶
B F-1.6-P磷酸酯酶
C1 丙酮酸羧化酶
B
C2 PEP羧激酶
磷酸二羟丙酮
天冬氨酸
PEP
C2
草酰乙酸
丙酮酸
-酮戊二酸 谷氨酸 穿梭系统
-酮戊二酸 谷氨酸
天冬氨酸
草酰乙酸
丙氨酸
C1 丙酮酸
3-P-甘油 乳酸
甘油
乙酰CoA
(胞液) (线粒体)
TCA循环 13
3-P-甘油醛
9.1 光合作用
光合作用是绿色植物通过叶绿体,利用光 能,把二氧化碳和水合成储存能量的有机物(葡萄 糖)并且释放出氧气的过程。
1
• 叶绿体中的色素
叶绿素
吸收红光 和蓝紫光
叶绿素a 叶绿素b
类胡萝卜素
吸收蓝紫光
胡萝卜素 叶黄素
2
3
•总反应式:
6CO2+12H2O*叶绿体 C6H12O6+6H2O+6O2* 光 •包括两个阶段: 1.光反应 2.暗反应 4
引物:结合有一个寡糖链的多肽 酶:糖原合成酶,分支酶 糖基供体:UDPG
1、糖原合成酶(glycogen synthase)
—— 催化-1,4-糖苷键合成
2、糖原分支酶 ( glycogen branching enzyme)
—— 催化-1,6-糖苷键合成
25
糖原合成酶
糖原合成酶催化的糖原合成反应不能从头开始合成第一 个糖分子,需要至少含4个葡萄糖残基的α-1,4-多聚葡萄糖作 为引物(primer),在其非还原性末端与UDPG反应,UDPG上 的葡萄糖基C1与糖原分子非还原末端C4形成α-1,4-糖苷链, 使糖原增加一个葡萄糖单位,UDPG是活泼葡萄糖基的供体, 其生成过程中消耗UTP,故糖原合成是耗能过程,糖原合成 酶只能促成α-1,4-糖苷键,因此该酶催化反应生成为α-1,4-糖 苷键相连构成的直链多糖分子。 酶催化合成糖原反应如下:
糖的生物合成-
注:支链淀粉降解时用的是脱支酶(R酶)
在Q酶作用下的支链淀粉的合成
“糖的生物合成”习题
一、名词解释: 糖异生、Cori循环、底物循环、丙酮酸羧化支路 二、问答题:
1、糖异生如何绕过几个‘能障’,需要什么酶的参与? 2、丙酮酸羧化酶催化丙酮酸转变为草酰乙酸。但是,只有在乙酰辅 酶A存在时,它才表现出较高活性。乙酰辅酶A的这种活化作用, 其生理意义何在?
• 所以不管草酰乙酸的去向如何,最终效应都是使体内的乙
酰辅酶A趋于平衡。
A
淀粉合成酶
+
ADP 直链淀粉(Gn+1)
A
(一)直链淀粉的生物合成-方式3 3、蔗糖转化为淀粉 2 3 4 Pi
1
5 6 7
8
1、蔗糖合成酶;2、UDPG焦磷酸化酶;3、ADPG焦磷酸化酶
4、淀粉合成酶; 5、果糖激酶;6、异构酶;7、变位酶;8、淀粉磷酸化酶
(二)支链淀粉的合成
1、淀粉合成酶: 只能催化形成α -1.4糖苷键 ,合成直链淀粉。 2 、 Q 酶(分支酶) : 既能催化 α -1.4 糖苷键 的断裂,又能催化α -1、6糖苷键的形成
二、蔗糖的生物合成-----有三条途径:
1、蔗糖磷酸化酶途径(微生物)
1-P葡萄糖+果糖 蔗糖磷酸化酶 蔗糖+Pi
2、蔗糖合成酶(植物) UDPG+果糖 蔗糖合成酶 UDP+蔗糖
也可利用ADPG,GDPG,TDPG,CDPG作为葡萄糖基供体。 3、磷酸蔗糖合成酶途径(植物光合组织) UDPG+6-P果糖
磷酸蔗糖合成酶
磷酸蔗糖+UDP
磷酸蔗糖 蔗糖+Pi 一般认为,此途径是植物合成蔗糖的主要途径。
糖类生物合成途径及其应用研究

糖类生物合成途径及其应用研究糖类是人类和其他生物体内不可或缺的重要营养物质,也是许多药物的基础。
糖类的合成和利用涉及多种生物化学反应,其中最重要的是糖类的生物合成途径。
本文将介绍糖类的生物合成途径及其应用研究。
一、糖类生物合成途径1. 糖原生物合成途径糖原是一种储存多余能量的多糖,也是人体内最重要的能量储备物质。
糖原的生物合成途径包括两种途径:糖原合成途径和糖原分解途径。
糖原合成途径主要涉及到葡萄糖,通过多个酶催化反应将葡萄糖转化为α-1,4- -D-葡萄糖苷键之间的分枝多糖分子,最终形成糖原。
糖原分解途径,则是糖原的分解过程,将其转化为葡萄糖分子释放能量。
2. 葡萄糖合成途径葡萄糖是生命活动所必需的主要能量源,其生物合成途径也是多种反应的复杂组合。
葡萄糖的生物合成途径同样需要多种酶的参与,在体内主要通过六碳糖的环化来合成葡萄糖分子。
此外,生命体需要维持体内葡萄糖水平的稳定,因此在葡萄糖的生物合成途径中,还需要进行调节糖联的产生和分解等。
3. 糖类的修饰途径糖类的修饰起到了重要的作用,可以改变糖类的结构、功能、稳定性、相互作用等等。
常见的糖类修饰途径包括糖基化、乙酰化、硫化、酯化等。
其中,糖基化是最为常见和复杂的一种修饰方式,通过酶的催化反应将糖分子与蛋白质、核酸等生物大分子连接,形成糖蛋白、糖核酸等新的复合生物大分子,所修饰的糖类不仅可做生物活性调节剂,同时也被广泛应用于医药、农业等领域。
二、糖类合成途径在医药、化妆品等领域的应用研究1. 新型药物开发糖类合成途径在新型药物开发领域有着广泛的应用。
糖蛋白、糖核酸等复合生物大分子是人体内最基本的分子之一,其糖基化修饰的差异常常会影响到人体生理状况。
因此,针对人体糖基化修饰失调的疾病,如糖尿病、肿瘤等,研究人员可以开发新型药物,调节糖基化修饰的平衡,减轻疾病症状。
2. 化妆品制造糖类作为功能性成分,除了在医药领域广泛应用外,在化妆品领域也有着广泛的应用。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
C3途径的三个阶段:
A、羧化阶段 3RuBP + 3CO2
B、还原阶段 6PGA + 6ATP + 6NDAPH C、再生阶段 5GAP + 3ATP
6PGA + 6H+
6GAP+6ADP+6NADP+ +6Pi 3RuBP + 3ADP + 2Pi
C3循环
R5P磷酸 异构酶 Ru5P激酶 核酮糖-5-P 转酮酶 差向异构酶 SBP磷酸酶 还原阶段 SBP醛 缩酶 还原阶段 FBP磷酸酶 二磷酸醛缩酶 3-磷酸甘油醛 转酮酶 羧化阶段
总反应式
3CO2+3H2O+3RuBP+9ATP+6NADPH G3P+6NADP++9ADP +3RuBP+9Pi
5.C3途径的调节
①自身催化:中间产物的浓度增加会调节卡尔文循环的 速率。 ②光调节:指光调节酶的活性。如:RuBPCase、NADPDPGA-DHase、1,6-P-F酶、1,7-P-S酶、Ru5P激酶。 ③光合产物运转的调节
磷酸转运体
在细胞质中合成蔗糖后释放出Pi,Pi进入叶绿体促进
磷酸丙糖运出叶绿体,光合速率加快。
(二)C4途径
C C 44 酸 酸
C4 酸 C3途径
草酰乙酸 (OAA)
PEP
C4途径
C3 酸
C3 酸
蔗糖 淀粉
1.C4途径的生化历程
①CO2的固定
COOH CH2 C-0 COOH PEP ②C4-二羧酸的转化和转移 PEPCase P +14CO2+H2O CH2 C=0
9.1.2 光能的吸收、转变和同化力产生
9.1.2.1 叶绿体的基本结构
A)被膜:有外膜和内膜两层,内膜具选择
透性。
B)基粒:由类囊体垛叠而成的。光能的吸收、
传递、转换场所。
C)间质:为叶绿体膜以内的基础物质。主要
是可溶性蛋白质(酶),为CO2固定与转化场 所。
叶绿体超微
结构电镜图
9.1.2.2 光合色素和光化学反应
B)从物质代谢角度看,光合作用过程是植物利用光能将
无机物(CO2和水),通过一系列复杂的化学变化,合 成碳水化合物等有机物的过程。
C)从能量代谢角度看,光合作用过程是植物将光能转变为化
学能 的过程。依此可将光合过程分为3大步骤:
1)原初反应:光能的吸收、传递和转换为电能; 2)电子传递和光合磷酸化:电能转变为活跃的化学能 3 )碳同化:活跃的化学能再转变为稳定的化学能。
③
CHO HCOH
ATP
HCOH
②
CH2O
PGA
P
CH2O
DPGA
P
CH2O
G3P
P
3.RuBP的再生
4.C3途径的计量关系
羧化阶段 3CO2 + 3RuBP+3H2O 3ADP+3Pi 3ATP 6PGA 6NADPH 5G3P+G3P+6NADP ++6ADP+6Pi 还原阶段 6PGA
6ATP
再生阶段
DHAP异构酶
1.CO2的固定 CH2O C=0 P CH2O P + COOH HCOH
Mg2+
HCOH+*CO2+H2O
①
HCOH
*COOH
HCOH
CH2O P
CH2O
PGA
P
PGA
RuBP
2.CO2的还原 COOH ADP C=0 NADPH+H 0 P HCOH NADP+ Pi
PSⅠ
PSⅡ
hv
hv
电子与质子传递链(Z scheme)
9.1.2.4 光合磷酸化
机理:化学渗透学说 类型:环式、非环式、假环式 同化力:ATP + NADPH
光合膜上电子和质子的传递及ATP生成
9.1.3 光合作用的碳素同化途径
(一) C3途径 (Calvin cycle):
CO2受体: 羧化酶: 初产物: RuBP Rubisco PGA (三碳化合物)
Acceptor (原初电子受体)
Pigment (作用中心色素)
Donor(原初电子供体)
光合作用的原初反应
为光合作用最初
的反应,它包括 对光能的吸收、 传递以及将光能 转换为电能的具 体过程。
H2O e D
光
光 光
P
反应中心
A NADP
9.1.2.3 电子传递与光合磷酸化
两个相互串连的光系统 PSⅠ:中心色素为P700,吸收长波红光(700nm) PSⅡ:中心色素为P680,吸收短波红光(680nm)
叶 绿 素 与 类 胡 萝 卜 素 的 结 构
2) 光合色素按其功能分为两类:
反应中心色素:具有光化学活性,可将光能转换为电能(陷阱), 少数特殊状态chla 天线色素:无光化学活性,只能吸收光能并传递 给中心色素, 大部分chla、全部chlb和类胡萝卜素
光合单位: 光合膜上能 进行完整光
14COOH
+Pi
OAA
③ PEP的再生
2. C4途径的类型
à À Í Ð NADPÆ » ¹ û Ë á à ¸ À à Ð Í NADÆ » ¹ û Ë á à ¸ À à Ð Í PEPô È » ¯ à ¸ À à Í Ð ¬ ¹ Î Ü Ê ø Ö Ð µ Ä C4á Ë » ¹ Æ û Ë á ì ¶ Ì ¬ ° ±Ë á ì ¶ Ì ¬ ° ±Ë á Ñ ô Í È ² ¿ Î » ¶ Â Ò Ì Ì å ß Á Ï £ Ì å ¸ ° Ï û Ö Ê ¶ È Ò â Ï ¸ ° û Ö Ð µ Ä C3á Ë û Í ± ª Ë á û Í ± ª Ë á û Í ± ª Ë á ¡ ¢ ± û ° ±Ë á ² Î Ö ï Ö À à ñ Ã Ó ×¡ ¢ ¸ Ê Õ á ¡ ¢ ¸ ß Á » ·Î ¹ ² ² Ý ¡ ¢ Â í ³ Ý Ü È ò ² Ñ Ý ¡ ¢ · Ç Ö Þ Ê ó Î ² Ë Ú
1.光合色素
1) 类别
光合色素 叶绿素a: 蓝绿色
1、叶绿素
叶绿素b: 黄绿色 胡萝卜素: 橙黄色 2 、类胡萝卜素 叶黄素: 黄色
3 、藻胆素
藻蓝素 藻红素
叶绿素:类胡萝卜素=3:1 所以叶片一般呈绿色 叶绿素a:叶绿素b=3:1 叶黄素:胡萝卜素=2:1
解释:秋后或衰老的叶片多呈黄色,秋后枫树叶子呈红色
P700* A0 A1 FX
-1. 2 -0. 8 P680*
E 0’(伏特)
-0. 4 0 0. 4 0. 8 1. 2 H2O Mn 1O +2 H + 2 2 Z P680
Pheo QA QB PQ cyt bH cyt bL FeS cyt f PC P700
FA/FB Fdx NADP +
反应的最小
结构单位
2、光化学反应—光能的转换
光化学反应实质就是由光引起的中心色素分子 与原初电子受体间的氧化还原反应。
PA
基态反应中心
P* . A
激发态反应中心
P+ . A电荷分离的反应中心
电荷分离后,反应中心情况为:
D.(P+. A- ). A1
D+.(P.A).A-1
原初反应的基本过程
D· P· A →D· P*· A →D· P+· A- →D+· P· AD· P· A D· P· A 为光系统或反应中心
9.1 光合作用
9.1.1 光合作用概述
一、概念 光能 氧化
CO2+ 2H2O*
还原
叶绿体
(CH2O) + H2O + O2*↑
ห้องสมุดไป่ตู้
注:由上式可知,光合作用释放的氧气来自于水。
A)光合作用过程相当复杂,光合作用靠光发动,但并非全
过程都需要光。根据需光与否,可将光合作用过程分为
光反应和暗反应。
光反应发 生在类囊 体膜上 暗反应发 生在叶绿 体的基质 中