六种石斑鱼核型特征比较和染色体进化研究
石斑鱼生物学及人工繁育研究进展

中国水产科学 2018年7月, 25(4): 737-752 Journal of Fishery Sciences of China研究论文收稿日期: 2018-04-01; 修订日期: 2018-04-16.基金项目: 科技部科技基础性工作专项(2013FY110700); 海南大学南海海洋资源利用国家重点实验室开放课题(2016001). 作者简介: 丁少雄(1973–), 教授, 从事鱼类资源保护与遗传育种研究. E-mail: sxding@ DOI: 10.3724/SP.J.1118.2018.18110石斑鱼生物学及人工繁育研究进展丁少雄1, 2, 刘巧红1, 3, 吴昊昊2, 曲朦31. 福建省海洋生物资源开发利用协同创新中心, 福建 厦门 361102;2. 厦门大学 海洋生物多样性与全球变化研究中心, 厦门大学 海洋与地球学院, 福建 厦门 361102;3. 厦门大学 海洋生物制备技术国家地方联合工程实验室, 厦门大学 海洋与地球学院, 福建 厦门 361102 摘要: 石斑鱼(Epinephelidae)是名贵的海产鱼类, 主要栖息于热带亚热带近岸岩滩和珊瑚礁水域, 是岩礁生态系生物群落的重要类群。
本文对国内外石斑鱼类生物学、生态学、渔业资源及人工繁育养殖的研究成果进行梳理, 综合报道石斑鱼类的形态比较特征, 栖息、摄食和繁殖习性, 系统演化与分类, 种类地理分布, 人工繁育等研究进展以及养殖发展概况; 报道全球石斑鱼自然资源明显衰退的现状评估以及对石斑鱼资源保护的共识和相关的渔业管理措施。
此外, 根据作者多年开展的鱼类遗传生物学研究, 指出强化我国海域野生石斑鱼资源保护工作, 深入开展杂交石斑鱼的基础生物学研究, 加强石斑鱼杂交选育行业管理, 是我国石斑鱼渔业产业可持续发展的必要保障。
关键词: 石斑鱼类; 生物学; 资源; 人工繁育; 研究进展中图分类号: S917 文献标志码: A 文章编号: 1005-8737-(2018)04-0737-16石斑鱼(Epinephelidae)是一类具有重要商业价值的鱼类, 广布于三大洋的热带与亚热带海域。
石斑鱼的性别分化和性腺发育机制研究

石斑鱼的性别分化和性腺发育机制研究石斑鱼是一种重要的经济性鱼类,在亚洲地区有广泛的养殖和消费市场。
然而,对于石斑鱼的性别分化和性腺发育机制仍存在较多的研究空白。
下面我将就这一主题展开探讨。
一、石斑鱼的性别分化在石斑鱼的性别分化中,存在着许多复杂的调控因素,如遗传因素、环境因素等。
其中,环境因素在形成石斑鱼性别决定中起着至关重要的作用。
比如说,养殖环境的水温、光照、饲料、密度等都会对石斑鱼的性别产生影响。
1.1 温度对石斑鱼性别的影响温度是影响石斑鱼性别决定的主要环境因素之一。
对于大多数石斑鱼而言,较高的水温通常会诱导雄性的发生,而较低的水温会使成鱼分化为雌性。
例如,常见的黑斑石斑鱼,温度在28-36℃之间是典型的雄性环境,而在22-28℃之间则是雌性环境。
这是由于在高温下,睾丸细胞可以继续分裂,而在低温下,卵巢细胞比较容易分裂。
因此,可以通过控制水温来实现养殖石斑鱼时的性别选择。
1.2 光照对石斑鱼性别的影响石斑鱼对于光照的敏感性也是其性别分化和性腺发育机制的重要因素之一。
日照时间和夜晚的亮度这两种因素都会对石斑鱼的性别分化产生影响。
在黑斑石斑鱼中,夜间亮度的增加会促进卵巢的成熟和卵子产生,而在白斑石斑鱼中则是促进睾丸的发育和精子的生产。
1.3 其他环境因素对石斑鱼性别的影响除温度和光照之外,水质、饲料等因素也会对石斑鱼的性别分化产生影响。
在石斑鱼的养殖中,通过控制这些因素,可以调整石斑鱼的性别分布,实现优质种苗的生产。
二、石斑鱼的性腺发育机制在石斑鱼的性腺发育机制研究中,主要研究的是石斑鱼的性腺组织结构、激素调节机制和生殖周期等。
2.1 石斑鱼的性腺组织结构石斑鱼的卵巢和睾丸都是由成排的生殖小叶组成的。
其中,卵巢小叶含有大量的卵母细胞,而睾丸小叶则含有大量成熟的精子。
在石斑鱼的性腺组织结构中,还存在着基质细胞和间质细胞等非特定性细胞。
2.2 石斑鱼的激素调节机制通过控制激素水平,可以调节石斑鱼性腺的生长和发育。
云纹石斑鱼、鞍带石斑鱼及杂交子代表型和遗传性状及亲缘关系分析

上 海 海 洋 大 学 硕士学位论文题 目:云纹石斑鱼、鞍带石斑鱼及杂交子代表型和遗传性状及亲缘关系分析英文题目: Phenotypic and genetic traits and geneticrelationship analysis in Epinephelusmoara (♀), E . lanceolatus (♂) and Yunlonggrouper专 业:动物遗传育种与繁殖 研究方向:石斑鱼杂交育种 姓 名:唐江 指导教师:田永胜 研究员二O 一八年 五月二十一日学校代码:研究生学号: M15010714910264上海海洋大学学位论文原创性声明本人郑重声明:我恪守学术道德,崇尚严谨学风。
所呈交的学位论文,是本人在导师的指导下,独立进行研究工作所取得的成果。
除文中已经明确注明和引用的内容外,本论文不包含任何其他个人或集体已经发表或撰写过的作品及成果的内容。
论文为本人亲自撰写,我对所写的内容负责,并完全意识到本声明的法律结果由本人承担。
学位论文作者签名:日期:年月日上海海洋大学学位论文版权使用授权书学位论文作者完全了解学校有关保留、使用学位论文的规定,同意学校保留并向国家有关部门或机构送交论文的复印件和电子版,允许论文被查阅或借阅。
本人授权上海海洋大学可以将本学位论文的全部或部分内容编入有关数据库进行检索,可以采用影印、缩印或扫描等复制手段保存和汇编本学位论文。
保密□,在年解密后适用本版权书。
本学位论文属于不保密□学位论文作者签名:指导教师签名:日期:年月日日期:年月日上海海洋大学硕士学位论文答辩委员会成员名单云纹石斑鱼、鞍带石斑鱼及杂交子代表型和遗传性状及亲缘关系分析Phenotypic and genetic traits and genetic relationship analysis in Epinephelus moara(♀), E. lanceolatus (♂) and Yunlong grouper学位论文完成时间:指导老师签字:答辩委员会成员签字:云纹石斑鱼、鞍带石斑鱼及杂交子代表型和遗传性状及亲缘关系分析摘要石斑鱼是世界重要海水名贵鱼类,养殖范围从我国的南方沿海城市逐渐发展到北方山东、天津等地,其具有重要的经济价值,市场需求大,受到人们的广泛的关注,20世纪末,研究学者们相继开展了石斑鱼人工繁殖与杂交育种技术,突破了石斑鱼在人工繁育上的瓶颈,使得石斑鱼产业取得了巨大的发展。
巨石斑鱼染色体核型分析
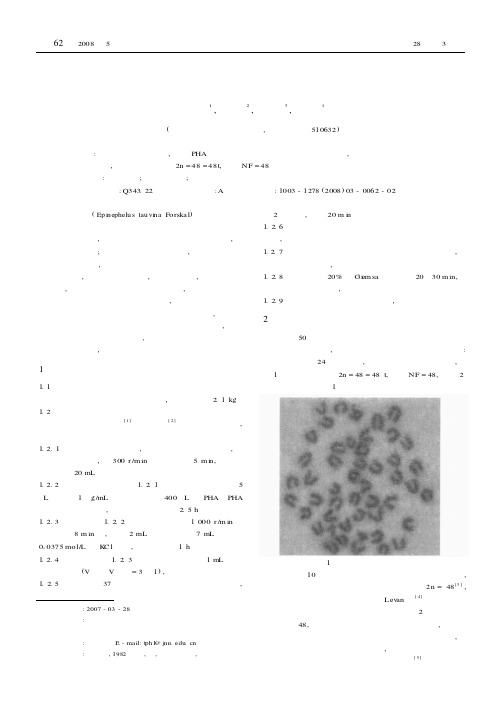
收稿日期:2007-03-28基金项目:中国水产科学院水产种质资源与养殖技术重点开放实验室资助通讯作者:蒲含林。
E -mail:t ph l @j .cn作者简介王小丽,年生,女,山西繁峙人,遗传学专业硕士研究生。
巨石斑鱼染色体核型分析王小丽1,郑元升2,戴 云3,蒲含林1(暨南大学生物工程研究所,广东广州 510632)摘要:以巨石斑鱼为材料,采用PHA 、秋水仙素体外培养肾细胞直接制片法,分析了巨石斑鱼染色体核型。
结果表明,巨石斑鱼核型为2n =48=48t,臂数NF =48。
关键词:巨石斑鱼;染色体核型;臂数中图分类号:Q343.22 文献标识码:A 文章编号:1003-1278(2008)03-0062-02 巨石斑鱼(Epi nephelus tauvi na Forska l )属硬骨鱼纲、鲈形目、石斑鱼亚科、石斑鱼属。
它主要分布于热带印度洋和太平洋近海,我国见于南海诸岛和台湾等地海域,属暖水性底层鱼类;栖息于底质粗杂的海区,以底栖甲壳类及其它鱼类为食,是石斑类中个体最大的种类。
巨石斑鱼俗称龙趸,因其肉富含胶质,味略似鸡肉,故也有海鸡鱼之称,其不仅味道鲜美而且营养丰富,是优质食用鱼。
由于巨石斑鱼是海洋中的珍贵品种,所以近年来对巨石斑鱼的研究主要集中于其海产养殖等相关方面,对细胞学方面的研究较少。
本文报道了巨石斑鱼的核型,对其进行细胞遗传学的基础研究,为巨石斑鱼养殖的选种育种提供基础资料,对促进其养殖业的发展具有重要意义。
1 材料与方法1.1 材料实验鱼购自黄沙清平水产市场,每条体重约2.1kg 。
1.2 方法实验方法参照舒琥[1]和王梅林[2]的方法进行修改,具体操作如下。
1.2.1 取材 活体取鱼肾,用预冷的生理盐水洗涤,之后将肾撕碎过滤,在300r /m in 转速下离心5m i n,然后吸取上清液20mL 。
1.2.2 秋水仙素作用 在1.2.1得到的上清液中加入5μL 浓度为1μg/mL 的秋水仙素和400μL 的PHA 。
海水养殖石斑鱼的线粒体基因组测序与分析

海水养殖石斑鱼的线粒体基因组测序与分析石斑鱼(Epinephelus coioides)作为一种重要的经济鱼类,在海水养殖业中具有广泛的应用前景。
了解石斑鱼的遗传特征对于改良品种、提高养殖效益以及保护自然资源都具有重要意义。
线粒体基因组是研究鱼类遗传变异和进化关系的重要工具。
本文将重点介绍海水养殖石斑鱼的线粒体基因组测序与分析,为石斑鱼遗传研究提供参考。
线粒体基因组是细胞中的一个独立进化的遗传系统,具有高度保守性和多样性。
它由几个蛋白编码基因、转运RNA基因和核酸RNA基因组成。
线粒体基因组的测序与分析可以提供关于个体间的亲缘关系、物种起源和进化关系的重要信息。
在进行石斑鱼线粒体基因组测序前,需要提取石斑鱼的线粒体DNA样品。
可以选择鳃、肌肉、鳍、眼球等组织进行提取。
提取的DNA样品可以通过PCR扩增获得所需长度的线粒体基因组。
然后,使用高通量测序技术对扩增的线粒体基因组进行测序。
线粒体基因组数据的测序分析包括序列的拼接、基因注释、遗传多样性和进化分析等。
首先,将测序得到的片段序列拼接成完整的线粒体基因组。
接下来,使用基因注释软件对拼接序列进行注释,识别编码基因和非编码区域。
然后,可以计算线粒体基因组的总碱基对数、GC含量等测序相关信息,评估测序质量。
此外,还可以进行基因组间的比较分析,研究不同个体之间的遗传差异和多样性。
通过构建系统进化树,可以探究石斑鱼的亲缘关系和物种起源。
石斑鱼线粒体基因组测序与分析可以揭示石斑鱼的遗传特征、进化关系以及与其他物种的亲缘关系。
基于线粒体基因组的研究结果,可以为石斑鱼的遗传育种提供依据。
通过遗传多样性分析,可以评估不同种源个体的遗传差异,为养殖策略的优化提供参考。
此外,通过比较不同物种的线粒体基因组,可以进一步揭示鱼类的进化历程和系统分类学。
尽管线粒体基因组测序和分析在石斑鱼遗传研究中发挥着巨大的作用,但还有一些需要注意的问题。
首先,测序时需要选择适当的测序策略和测序深度,以确保获得具有高质量的线粒体基因组数据。
石斑鱼遗传多样性的研究进展

第24卷第8期2005年8月水产科学FISHERIES SCIE NCEV ol.24N o.8Aug.2005石斑鱼遗传多样性的研究进展尹绍武1,2,黄 海1,张 本1,陈国华1(1.海南大学海洋学院,海南 海口 570228;2.农业部渔业资源可持续利用重点开放实验室,山东 青岛 266071)关键词:石斑鱼属;表型;染色体;同工酶;DNA ;遗传多样性中图分类号:S917文献标识码:C文章编号:100321111(2005)0820046204 收稿日期:2005-01-06; 修回日期:2005-02-011 基金项目:农业部渔业资源可持续利用重点开放实验室开放课题资助(实开2004-08). 作者简介:尹绍武(1969-),男,副教授,博士;研究方向:鱼类遗传育种与发育生物学. 遗传多样性(genetics diversity )又称基因多样性(genes di 2versity )。
它不仅是生物多样性的核心组成部分,而且是物种多样性和生态系统多样性的基础,也是生命进化和物种分化的基础,更是评价自然生物资源的重要依据。
通常所说的遗传多样性是指种内不同种群之间或一个种群内不同个体之间遗传变异的总和,其表现是多层次、多水平的,主要包括表型多态、染色体多态、蛋白质多态以及DNA 多态。
鱼类遗传多样性是鱼类生存与遗传进化的基础。
开展鱼类遗传多样性的研究不仅有利于渔业资源的管理与保护,而且为鱼类遗传育种提供科学依据[1,2,6]。
石斑鱼是重要的世界性海水经济鱼类,也是海水增养殖的对象之一。
近年来,由于人类活动的不断加剧,掠夺性开发、过度捕捞、海洋环境的污染和破坏、养殖群体的近亲繁殖、外来种侵入等原因,鱼类资源日益衰退,许多重要的经济鱼类已经不再形成鱼汛,而且种质资源在不断退化[6,7]。
石斑鱼的种质资源现状如何?如何有效地保护与开发?这些问题都值得我们去探讨。
本文分别从表型、染色体、蛋白质、DNA 四个层次上简要概述国内外石斑鱼遗传多样性的研究进展,以期为今后开展石斑鱼的种质资源保护和遗传育种提供参考。
鞍带石斑鱼、棕点石斑鱼及其杂交子代DNA甲基化的MSAP分析

5 2 %, 2 3 . 4 4 %, 1 4 . 7 6 %, 杂交子代 的半 甲基化率显 著低于亲 本的半 甲基化率. 研究表 明 , 鞍带石斑 鱼与棕点
石斑鱼的杂交子代在基 因组层 面上 和双 亲相 比发 生 了较 大 的甲基化水 平 的调整 , 于不 同位点 , D N A甲基化 的增强或减弱对石斑鱼杂种优势可能会产生影 响. 关键词 : 石斑鱼 ; D N A 甲基化 ;甲基化敏感 扩增多态性 ;杂种 优势
第3 5 卷 第2 期
2 0 1 7年 6月
海 南 大 学 学 报 自 然 科 学 版
NATURAL S CI ENCE J OURNAL OF HAI NAN UNI VERS I TY
V0 1 . 3 5 N o . 2
J u n . 2 0l 7
文章编号 : 1 0 0 4—1 7 2 9 ( 2 0 1 7 ) 0 2— 0 1 4 5— 0 7
鞍 带石 斑 鱼 、 棕 点 石 斑 鱼 及 其 杂 交 子 代 D N A 甲基 化 的 MS A P分 析
我国海洋鱼类染色体组型研究

我国海洋鱼类染色体组型研究摘要:染色体组型研究是细胞遗传学的基础, 随着细胞遗传学的深入发展, 愈来愈多的生物学家认识到作为遗传信息载体的染色体,非但其数目和形态结构具有物种的特征,而且还反映出生物进化的历史。
对其的研究有着十分重要的意义。
关键词:海洋鱼类染色体组型遗传资源染色体组型又称核型(Karyo type)。
染色体组型研究是细胞遗传学的基础, 随着细胞遗传学的深入发展, 愈来愈多的生物学家认识到作为遗传信息载体的染色体, 非但其数目和形态结构具有物种的特征, 而且还反映出生物进化的历史。
从现存物种的染色体组型研究和比较分析中来探讨种群的进化路线和亲缘关系, 不仅对于分类学和系统发生研究有着十分重要的意义; 而且对于现代分子生物学中的基因定位、原位杂交、种间杂交鉴定和多倍体育种等方面也有重要意义。
我国海域辽阔, 沿岸曲折, 栖息着丰富的海洋生物, 有成千上万种的鱼类和贝类, 开展鱼类和贝类染色体组型研究的前景十分广阔。
一、我国海水鱼类染色体组型特征1.染色体数目我国海水鱼类的染色体数目特点有: ①染色体数目较少,无多倍化。
②染色体数目变异较小,分布范围相对狭窄,呈连续分布。
染色体数目可分8种类型,淡水鱼类由于多倍化,部分类群的染色体数目可达260条。
海水鱼类染色体的数目变异可能主要来自罗伯逊易位(Robertsonian translocation)。
原始基本型数经罗伯逊易位,少数染色体着丝粒融合,不断特化演变成染色体数等类型。
2.染色体形态海水鱼类染色体形态的共同特征之一是端部、亚端部着丝粒染色体较多,而中部、亚中部着丝粒染色体较少。
在研究的50种海水鱼类中,除鳗鲡目外,其他6个目都明显体现了这一特性。
鲱形目、鲻形目、鲈形目和鲽形目等中的许多种类的染色体全部为端部、亚端部着丝粒染色体。
染色体结构重排和异染色质扩增,是引起染色体形态变化的主要原因。
而臂间重排会直接引起染色体的臂比变化,导致染色体形态发生相应变化。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
六种石斑鱼核型特征比较和染色体进化研究采用常规核型分析、C-带、Ag-NORs、DAPI荧光染料染色以及几种重复序列(5SrDNA、18S rDNA和(TTAGGG)_n)的荧光原位杂交技术对斜带石斑鱼E.coioides、青石斑鱼E.awara、拟青石斑鱼E.fasciatomaculosus、赤点石斑鱼E.akaara、褐石斑鱼E.bruneus和镶点石斑鱼E.amblycephalus的核型特征进行描述比较和研究,结果如下:1、6种石斑鱼二倍体染色体数目均为48,但其染色体组组成具有一定的差异,核型公式分别为:斜带石斑鱼2n=48,2sm+46t,NF=50;青石斑鱼
2n=48,48t,NF=48;拟青石斑鱼2n=48,1st+47t,NF=48;赤点石斑鱼
2n=48,2sm+8st+38t,NF=50;褐石斑鱼2n=48,2m+4sm+42t,NF=54;镶点石斑鱼
2n=48,2m+46t,NF=50。
2、分析了C-带和DAPI荧光带在6种石斑鱼染色体上的
分布,6种石斑鱼染色体C-带和DAPI荧光带的分布模式均不同。
根据染色体组中是否绝大多数染色体的着丝粒区域为C—带异染色质这一
特征,可将6种石斑鱼分为2种类型:(1)染色体组中绝大多数染色体具有着丝粒带,此类型中包括斜带石斑鱼和褐石斑鱼;(2)染色体组中绝大多数染色体不具有着丝粒带,此类型又可根据是否具有恒定的异染色质带细分为:a、绝大多数染色体无着丝粒带,仅双臂染色体的整条短臂为恒定的异染色质带,如拟青石斑鱼和
赤点石斑鱼,b、绝大多数染色体无着丝粒带,仅在部分染色体的着丝点位置具有变动带,此类型包括青石斑鱼和镶点石斑鱼。
DAPI荧光带的研究结果表明,除斜
带石斑鱼外,其它5种石斑鱼均具有DAPI荧光亮带,具体模式如下:斜带石斑鱼各条染色体被均匀染色,无明显DAPI荧光带;青石斑鱼与褐石斑鱼在其着丝点位置具有荧光亮带,但青石斑鱼的荧光带信号非常微弱而褐石斑鱼则较强;拟青石斑
鱼和赤点石斑鱼染色体组中双臂染色体的整个短臂被DAPI染成明亮的荧光带;
镶点石斑鱼在一些染色体的着丝点位置和第24对染色体上靠近端粒位置显示为荧光亮带。
褐石斑鱼绝大多数染色体着丝粒位置,拟青石斑鱼和赤点石斑鱼所有双臂染色体的整个短臂既表现为C带阳性也表现为DAPI荧光亮带,说明这些位置染色体的结构和碱基组成相似,可能具有同源性。
3、不论是常规的Giemsa染色,还是C —带以及DAPI荧光染色体,拟青石斑鱼染色体组中第1对染色体为异形。
该对染色体由1条端部着丝粒染色体和1条亚端部着丝粒染色体组成,C—带和DAPI荧光带均表明该对染色体中的亚端部着丝粒染色体,其整个短臂为染色体组中唯一1条可以检测到C带阳性以及DAPI荧光亮带的染色体,而同源染色体中的另外1条则既无异染色质带也无DAPI荧光带,可见该对同源染色体为1对异形染色体。
这种同源染色体异形的现象在石斑鱼属中属首次发现。
4、银染结果表明,在6种石斑鱼种内和种间,NORs数目和分布模式具有多态性。
青石斑鱼、拟青石斑鱼、赤点石斑鱼和镶点石斑鱼染色体组中仅有1对核仁组织区位于第24对染色体靠近着丝粒的位置上;斜带石斑鱼仅在第24对染色体的短臂上具有1对NORs;褐石斑鱼染色体组中核仁组织区数目2~5个,位于双臂染色体,既第2、9、24对染色体短臂上。
5、利用荧光原位杂交技术研究5S rDNA和18S rDNA在6种石斑鱼染色体上的分布模式。
结果表明5S rDNA在6种石斑鱼染色体组中,均位于1对中等大小端部着丝粒染色体且靠近着丝点的位置上,种间和种内未观察到差异,在染色体上的分布模式基本一致。
这表明5S rDNA在石斑鱼属鱼类染色体上的分布模式具有较高的保守性;6种石斑鱼染色体组中,具有5S rDNA的染色体具有同源性。
利用FISH研究18S
rDNA在6种石斑鱼染色体上的分布模式,结果表明:褐石斑鱼在3对(第2,9和24对)染色体上具有明显杂交信号,并且在一些染色体的末端也有较小和微弱的信号,检测到的rDNA位点多于银染检测到的位点数;其它5种石斑鱼只能在第24对染色体上检测到唯一1对杂交信号,与银染结果一致。
6种石斑鱼第24对染色体上均具有NORs说明第24对染色体在6种石斑鱼中具有同源性。
6、利用FISH技术检测端粒序列在6种石斑鱼染色体上的分布,端粒探针杂交信号仅局限在所有染色体的端粒位置,在染色体上的中间部位未检测到杂交信号。
但在褐石斑鱼中,发现有10对染色体的端粒位置,杂交信号强度明显比其它位点的强度高,信号大小也比其它染色体上的大,说明这些位置上的端粒序列拷贝数明显高于其它位点。
7、为了探索石斑鱼属鱼类进化分歧过程中染色体进化趋势与途径,本文综合了6种石斑鱼核型特征信息建立了系统进化树。
通过与前人利用分子标记构建的系统进化树进行对比,发现2种进化树之间具有较好的一一对应关系。
此外,系统进化树还揭示了石斑鱼属染色体的进化规律:石斑鱼属系统进化树中不同分支中的石斑鱼染色体进化采用不同形式的染色体重组以实现进化分歧。