第3章 基因与基因组的结构
遗传学第三章 基因的概念和结构
基因重叠方式
• Mis-reading for stop codon
( Q RNA virus 1973. A. Weiner )
400Nt
800Nt
AUG----------------------UGA-----------------------UAA
设有两个独立起源的隐性突变,具有类似的表现型。判断是属于同一个基因 突变,还是属于两个基因突变?即判断是否属于等位基因? ①建立双突变杂合二倍体; ②测定突变间有无互补作用。
• 顺反测验:顺式排列为对照(是两个突变座位位于同一条染色 体上),其表现型野生型。实质上是进行反式测验(反式排列是 两个突变座位位于不同的染色体上)。
① 反式排列为野生型:突变分属于两个基因位点; ② 反式排列为突变型:突变分属于同一基因位点。
Complementary assay
rII47 0
rII106 0
rII 47
rII106
rII106 0 rII51 0
rII106 rII51
Why?
plane E.coli K12
依据; One gene
2、假基因(pseudo gene)
• 假基因:同已知的基因相似,处于不同的位点,因缺失或突变而 不能转录或翻译,是没有功能的基因。
第五节、外显子和内含子
• 内含子(intron):DNA序列中不出现在成熟mRNA的片段; • 外显子(extron):DNA序列中出现在成熟mRNA中的片段。
Ovalbumin DNA X cDNA
5387 bp 11 genes 3 mRNA 9 peptides
第三章遗传物质的基础1

蛋白复杂得多,具种和组织特异性。
组蛋白 类型
H1 H2A H2B H3 H4
小牛胸腺染色体组蛋白的特点
碱性氨基酸
Lys
Arg
29%
1%
11%
9%
16%
6%
10%
13%
11%
14%
氨 基 酸 分子量
数
(Da)
215
23,000
129
13,960
125
13,775
135
15,340
102
11,280
•染色体的结构 分子生物学和生物化学研究表明,染色体基本结构单位为核 小体,核小体连接成染色质丝,经卷曲形成螺线管solenoid, (中期)后者进一步卷曲成超粗纤维,再进一步浓缩即为染 色体。高度浓缩的染色体长度只有DNA双螺旋的1/万左右。
第三章 遗传的物质基础
本章内容:
染色体 核酸 基因的组织与结构
遗传物质
生物性状
遗传物质的本质的揭示:
➢ 孟德尔(Mendel,G.J.)1856—1864年进行豌豆杂交 试验,发现分离和独立分配遗传规律
只是一种逻辑推理产物 没有任何物质内容
认为生物性状是受细胞里的颗粒性遗传因子控制
➢ 约翰生(Johannsen,W.L.)1909年用“基因” (gene)一词代替孟德尔的遗传因子概念
二、原核生物及病毒染色体结构
以大肠杆菌为例来阐明原核生 物染色体结构特点。
•大肠杆菌染色体以单个双链环状DNA分子构成,大约有 4.6×106bp。
•这种染色体组成了大肠杆菌的拟核(核质体)。
•在拟核中DNA占80%,其余为RNA和蛋白质。
基因和基因组(生物化学)

GC盒(GC Box)
位于-35bp ,GGCGG,与转录因 子SP1结合,促进转录的过程。
-35
-35
+1
12
(2).增强子(enhancer)
与转录因子特异性结合,增强 转录活性,在基因任意位置都有效、 无方向性。
promoter
CAAT box TATA box 5′ exon intron
5′--------AAUAAA ------AAAAAAAA 3′ 多聚腺苷酸化 14
mRNA
真核生物基因的结构
调控序列
promoter
Enhancer
5′
CAAT box TATA box
结构基因
调控序列
Poly(A) 加尾信号 UTR
UTR
exon intron
exon intron
exon TGA Stop 3′
内含子的 5′端以GT开始, 3′端以AG结束。
AG GT AG
5′
GT
3′
intron1 intron2 exon3 exon1 exon2
6
(二) 转录调控序列
结构基因编码区两侧的一段不被翻译 的DNA片段(侧翼序列),参与转录调控。
侧翼序列
前导序列 5′
intron1 exon1 exon2
TATA盒(TATA Box): 位于-25~-30bp,TATAAAA/TATATAT 与TFII结合,启动基因转录。
-30
-25
+1
10
CAAT盒(CAAT Box)
位于-70~-80bp,GG C/ T CAATCT,
与CTF结合,决定启动子转录效率。
-80
《现代分子生物学》第三版 (朱玉贤 李毅 主编)课后习题答案 高等教育出版社

现代分子生物学课后习题及答案(共10章)第一章绪论1. 你对现代分子生物学的含义和包括的研究范围是怎么理解的?2. 分子生物学研究内容有哪些方面?3. 分子生物学发展前景如何?4. 人类基因组计划完成的社会意义和科学意义是什么?答案:1. 分子生物学是从分子水平研究生命本质的一门新兴边缘学科,它以核酸和蛋白质等生物大分子的结构及其在遗传信息和细胞信息传递中的作用为研究对象,是当前生命科学中发展最快并正在与其它学科广泛交叉与渗透的重要前沿领域。
狭义:偏重于核酸的分子生物学,主要研究基因或 DNA 的复制、转录、达和调节控制等过程,其中也涉及与这些过程有关的蛋白质和酶的结构与功能的研究。
分子生物学的发展为人类认识生命现象带来了前所未有的机会,也为人类利用和改造生物创造了极为广阔的前景。
所谓在分子水平上研究生命的本质主要是指对遗传、生殖、生长和发育等生命基本特征的分子机理的阐明,从而为利用和改造生物奠定理论基础和提供新的手段。
这里的分子水平指的是那些携带遗传信息的核酸和在遗传信息传递及细胞内、细胞间通讯过程中发挥着重要作用的蛋白质等生物大分子。
这些生物大分子均具有较大的分子量,由简单的小分子核苷酸或氨基酸排列组合以蕴藏各种信息,并且具有复杂的空间结构以形成精确的相互作用系统,由此构成生物的多样化和生物个体精确的生长发育和代谢调节控制系统。
阐明这些复杂的结构及结构与功能的关系是分子生物学的主要任务。
2. 分子生物学主要包含以下三部分研究内容:A.核酸的分子生物学,核酸的分子生物学研究核酸的结构及其功能。
由于核酸的主要作用是携带和传递遗传信息,因此分子遗传学(moleculargenetics)是其主要组成部分。
由于 50 年代以来的迅速发展,该领域已形成了比较完整的理论体系和研究技术,是目前分子生物学内容最丰富的一个领域。
研究内容包括核酸/基因组的结构、遗传信息的复制、转录与翻译,核酸存储的信息修复与突变,基因达调控和基因工程技术的发展和应用等。
(整理)第三章基因与基因组

第三章基因与基因组第一节基因概念的历史演变第二节DNA与基因第三节真核生物的割裂基因第四节基因大小第五节重叠基因第六节真核生物的基因组第七节真核生物DNA序列组织第八节细胞器基因组第九节基因鉴定第十节人类基因组计划第三章基因与基因组1 基因(gene)的概念基因是遗传的功能单位,DNA分子中不同排列顺序的DNA片段构成特定的功能单位;含有合成有功能的蛋白质多肽链或RNA所必需的全部核苷酸序列。
广义地说,基因是有功能的DNA片段。
第一节基因概念的历史演变2 基因概念的历史演变:(1)Mendel提出基因的存在(2)Morgan证实基因在染色体上(3)“一个基因一个酶”修正为“一个基因一个多肽链”“基因”一词的创立: 1909年,丹麦遗传学家约翰逊“基因”(gene)。
Gregor MendelThomas Hunt Morgan3 基因概念的理论基础3.1 一个基因一个酶1941年G W Beadle 和E L Tatum研究证实红色链孢霉各种突变体的异常代谢是一种酶的缺陷,产生这种酶缺陷的原因是单个基因的突变。
3.2 一个基因一条多肽链本世纪50年代,Yanofsky有些蛋白质不只由一种肽链组成,如血红蛋白和胰岛素,不同肽链由不同基因编码,因而又提出了“一个基因一条多肽链”的假设。
3.3 基因的化学本质是DNA(有时是RNA)1944年,O T Avery 证实了DNA是遗传物质。
有些病毒只含有RNA。
1953年沃森和克里克建立DNA分子的双螺旋结构模型。
3.4 基因顺反子(Cistron)的概念1955年,美国本兹尔(Benzer)提出顺反子的概念:是指编码一个蛋白质的全部组成所需信息的最短片段,即一个基因。
基因仅是一个功能单位,基因内部的碱基对才是重组单位和突变单位。
一对同源染色体上两突变(a和b)在同一染色体上时,称为顺式构型,在两个染色体上时,为反式构型;顺反互补测验(cis-trans test):比较顺式和反式构型个体的表型来判断两个突变是否发生在一个基因(顺反子)内的测验。
第三章--基因与基因组的结构PPT课件

-
4
③近20年来,由于重组DNA技术的完善和应 用,人们已经改变了从表型到基因型的传统 研究基因的途径,而能够直接从克隆目的基 因出发,研究基因的功能及其与表型之间的 关系,使基因的研究进入了反向生物学阶段。
-
5
• 反向生物学:指利用重组DNA技术和离体 定向诱变的方法研究已知结构的基因相应的 功能,在体外使基因突变,再导入体内,检 测突变的遗传效应即表型的过程。
• 例如,对于大肠杆菌和其他细菌,用三个小写
字母表示一个操纵子,接着的大写字母表示不
同基因座,lac 操纵子的基因座:lacZ,lacY, lacA;其表达产物蛋白质则是lacZ,lacY,
lacA。
-
37
• 3.质粒和其他染色体外成分的命名 • 自然产生的质粒,用三个正体字母表示,第—
个字母大写,例如:ColEⅠ;
血破裂而使血红蛋白计数减少,造成贫血。
• 其本质是其血红蛋白的β-链与正常野生型
β-链之间的第6位氨基酸,由Val取代了 Glu所致。
-
32
• 这种贫血病是由基因突变造成的一种分子病,
除溶血后发生贫血外,还会堵塞血管形成栓塞, 从而伤及多种器官。
• 它的纯合子(通过单倍体形成的纯系双倍体)患
者在童年就夭折。
-
40
• 6.线虫基因的命名
• 用三个小写斜体字母表示突变表型,如存
在不止一个基因座,则在连字符后用数字
表示,如基因unc-86,ced-9;蛋白UNC-
86;CED-9。
-
41
• 7.植物基因的命名
• 多数用1~3个小写英文斜体字母表示。
-
42
• 8.脊椎动物基因的命名
基因组
(1)几个结构基因的编码区无间隔:几个基因 几个结构基因的编码区无间隔: 的编码区是连续的、不间断的, 的编码区是连续的、不间断的,即编码一条多 肽链,翻译后切割成几个蛋白质。 肽链,翻译后切割成几个蛋白质。 mRNA没有 ′端帽子结构 没有5 (2)mRNA没有5 ′端帽子结构 5 ′ 端非编码区 RNA形成特殊的空间结构称翻译增强子 形成特殊的空间结构称翻译增强子。 的RNA形成特殊的空间结构称翻译增强子。 (3)结构基因本身没有翻译起始序列,必须在 结构基因本身没有翻译起始序列, 转录后进行加工、剪接,与病毒RNA5 ′端的帽 转录后进行加工、剪接,与病毒RNA5 ′端的帽 结构相连,或与其它基因的起始密码子连接, 结构相连,或与其它基因的起始密码子连接, 成为有翻译功能的完整mRNA。 成为有翻译功能的完整mRNA。
细菌多数基因按功能相关成串排列,组成操纵元的基因 细菌多数基因按功能相关成串排列, 表达调控的单元,共同开启或关闭, 表达调控的单元,共同开启或关闭,转录出多顺反子的 mRNA; mRNA;真核生物则是一个结构基因转录生成一条 mRNA, mRNA是单顺反子 mRNA,即mRNA是单顺反子,基本上没有操纵元的结 是单顺反子, 构,而真核细胞的许多活性蛋白是由相同和不同的多肽 形成的亚基构成的, 形成的亚基构成的,这就涉及到多个基因协调表达的问 真核生物基因协调表达要比原核生物复杂得多。 题,真核生物基因协调表达要比原核生物复杂得多。 原核基因组的大部分序列都为基因编码, 基因组的大部分序列都为基因编码 原核基因组的大部分序列都为基因编码,而核酸杂交等 实验表明:哺乳类基因组中仅约10%的序列为蛋白质 基因组中仅约 的序列为蛋白质、 实验表明:哺乳类基因组中仅约10%的序列为蛋白质、 rRNA、tRNA等编码 其余约90%的序列功能至今还不 rRNA、tRNA等编码,其余约90%的序列功能至今还不 等编码, 清楚。 清楚。 原核生物的基因为蛋白质编码的序列绝大多数是连续的, 原核生物的基因为蛋白质编码的序列绝大多数是连续的, 而真核生物为蛋白质编码的基因绝大多数是不连续的, 而真核生物为蛋白质编码的基因绝大多数是不连续的, 即有外显子(exon)和内含子 和内含子(intron), 即有外显子(exon)和内含子(intron),转录后需经剪接 (splicing)去除内含子 才能翻译获得完整的蛋白质, (splicing)去除内含子,才能翻译获得完整的蛋白质,这 去除内含子, 就增加了基因表达调控的环节。 就增加了基因表达调控的环节。
第三章基因组作图
样品将分别取自亚、欧、非裔人群,我国将提供一半的亚裔样品,
占到研究样品总数的1/6。这意味着我国虽然只做10%的构建工作,
但可以得到几十份全基因组规模的单体型图,并且获得与其它族裔
的比对结果。我国样品将在北京师范大学进行采集。
(4)中国10%单体型图计划
O 北京、香港和台湾地区的科学家将共同参加这一重大国际合作科研项目,
进行国际交流,并统一向国际中心提交数据。
O 中国协调组由杨焕明院士任组长。香港大学、香港科技大学和香港中文大
学计划共同承担单体型图2%的任务,台湾中研院下属的生物医学科学研
究所计划承担约1.5%左右的工作。其余6.5%任务由大陆科学家承担。
(5)基因组单体型图的意义
①是人类基因组的遗传整合图,将对基因组的研究更为全面、有效、准
筛查后进行结构学和功能学验证
物理图构建
O
低精度物理作图:构建覆盖每条染色体的数百kb的大
片段的图谱。
O
高精度物理作图:构建覆盖每条染色体的几十个kb的
图谱。
物理图作图方法Ⅱ:荧光原位杂交(FISH)作图
O
将一组不同颜色的荧光标记探针与单个变性的染色体杂交,
分辨出每种杂交信号,从而测定出各探针序列的相对位置。
O
缺点:最终排序结果的拼接组装比较困难,尤其在部分重复序列较
高的地方难度较大。
2、逐个克隆法
O 对连续克隆系中排定的YAC克隆逐个进行亚克隆测序并进行组装:
O 遗传图-物理图-亚克隆测序-计算机拼装。
O 理想状况下,整条染色体就是由一个完整的重叠群构成。
基因组“草图”和“完成图
O
草图绘制的是整个基因组的框架,完成图则是基因组序列
生物第三章基因的本质知识点
生物第三章基因的本质知识点
生物第三章基因的本质主要包括以下知识点:
1. 基因的定义:基因是遗传信息的基本单位,是控制生物体形态、结构和功能的DNA 序列。
2. 基因的结构和组成:基因由DNA分子组成,包括编码区和非编码区。
编码区包含编码基因的信息,非编码区包含调控基因表达的元素。
3. 基因的功能:基因通过编码蛋白质来执行特定的功能,如调节细胞生长、发育和代
谢等。
4. DNA的复制:DNA分子在细胞分裂时通过复制过程来传递基因信息,确保每个新生细胞都有完整的基因组。
5. 基因的表达:基因表达是指基因转录为mRNA分子,并经过翻译过程产生蛋白质。
6. DNA的转录:DNA转录为mRNA过程包括启动子、RNA聚合酶、转录因子等多个
环节的参与。
7. 基因的翻译:mRNA通过核糖体和tRNA的参与,翻译成氨基酸序列,形成蛋白质。
8. 基因突变:基因突变指基因序列发生变化,包括点突变、插入突变、缺失突变等,
可能导致基因功能的改变。
9. 基因的遗传:基因通过遗传方式传递给下一代,确定了后代的表型和遗传特征。
10. 基因的调控:基因的表达可以受到内、外界环境的调控,通过启动子、转录因子等参与的调控元素来实现。
以上是关于生物第三章基因的本质的主要知识点,可以帮助我们理解基因的结构、功能和遗传规律。
第三章基因与基因组的结构
叶绿体基因组
叶绿体也属于半自主性的细 胞器,同mtDNA类似,叶绿体 DNA也以双链环状分子的形式 存在于细胞质中。
一般而言,一个顺反子就是一个基因, 大约1500个核苷酸。
它是由一群突变单位和重组单位组成 的线形结构(因为任何一个基因都是突变 体或重组体)。
基因是DNA分子上的功能单位,在染 色体或DNA分子上,基因成串排列,它既 是遗传的功能单位,同时也是交换单位 和突变单位。
但并非所有的DNA序列都是基因,只 有某些特定的多核苷酸区段才是基因的 编码区。
染色体上存在大量无转录活 性的重复DNA序列,主要有两 种形式:
➢ 串联重复的DNA,如卫星DNA,
成簇存在于染色体的特定区域;
➢ 分散重复的DNA,分散在染色体 的各个位点。
卫星DNA
有些高度重复的DNA序列 的碱基组成和浮力密度与主体 DNA不同,在CsCl密度梯度离 心时,可形成相对独立于主 DNA带的卫星带。
例如rRNA和某些tRNA基因以及重 复程度更高的卫星DNA等。
不同生物中 非重复基因占基 因组的比例差别 很大。原核生物 含有完全不重复 的DNA,低等真 核生物的大部分 DNA是非重复的, 高等真核生物中 非重复基因所占 基因组的比例更 低。
从不同种类的原核生物的Cot曲线可以看出, 各种原核生物的Cot曲线形状都很相似,跨度一 般只有两个数量级,只是Cot1/2值不同。表明各
➢ 单拷贝序列
➢ 轻度重复序列
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第3章基因与基因组的结构1.主要内容1)断裂基因构成性质2)重叠基因种类3)C值矛盾4)原核生物与真核生物基因组的区别5)真核生物染色体的结构6)真核生物DNA序列的4种类型7)基因家族、基因簇、卫星DNA、分散重复DNA 序列8)人类基因组计划2.教学要求1)掌握基因,断裂基因,顺反子,C值矛盾,重叠基因,基因家族,重复序列,卫星DNA等基本概念;2)熟悉原核生物和真核生物基因组结构特点与功能;3)了解人类基因组的重复顺序、人类基因组计划。
第1节基因的概念第2节基因命名简介第3节真核生物的断裂基因第4节基因及基因组的大小与C值矛盾第5节重叠基因第6节基因组第7节真核生物DNA序列组织第8节基因家族第9节人类基因组研究进展第1节基因的概念●基因:带有特定遗传信息的核酸分子片段。
包括结构基因:编码蛋白质tRNA rRNA调控基因:●基因研究的发展染色体分子反向生物学●基因位于染色体和细胞器的DNA分子上•基因和顺反子•1955,Benzer用以表述T4 具溶菌功能的区的2个亚区: rⅡA rⅡB•现代分子生物学文献中,顺反子和基因这两个术语互相通用。
第2节基因命名简介•表示基因3个小写斜体字母,lac•表示基因座3个小写斜体字母+ 1个大写斜体字母。
lacZ•表示质粒自然质粒 3 个正体字母,首字母大写重组质粒在2个大写字母前面加小写p•基因为斜体,蛋白质为正体•人类基因为大写斜体第3节真核生物的断裂基因•一、割裂基因的发现•1977,通过成熟mRNA(或cDNA)与编码基因的DNA杂交试验而发现•真核生物的基因是不连续的,大大改变了原来对基因结构的看法,现在知道大多数真核生物的基因都是不连续基因或割裂基因(split gene)。
•割裂基因的概念——是编码序列在DNA分子上不连续排列而被不编码的序列所隔开的基因。
•割裂基因的构成•构成割裂基因的DNA序列被分为两类:•基因中编码的序列称为外显子(exon),外显子是基因中对应于信使RNA序列的区域;•不编码的间隔序列称为内含子(intron),内含子是从信使RNA中消失的区域。
•割裂基因由一系列交替存在的外显子和内含子构成,基因两端起始和结束于外显子,对应于其转录产物RNA的5’和3’端。
如果一个基因有n个内含子,则相应地含有n+1个外显子。
●割裂基因的性质•Splitting Gene 的普遍性•外显子和内含子各有特点•Splitting gene 概念的相对性●Splitting Gene 的普遍性a) 真核生物(Eukaryots)中•绝大部分结构基因•tDNA, rDNA•mtDNA, cpDNAb) 原核生物(Prokaryots)中•SV40 大T 抗原gene•小t 抗原gene•Splitting gene 并非真核生物所特有●外显子和内含子各有特点•割裂基因的外显子在基因中的排列顺序和它在成熟mRNA产物中的排列顺序是相同的;•某种割裂基因在所有组织中都具有相同的内含子成分;•核基因的内含子通常在所有的可读框中都含有无义密码子(nonsense codon),因此一般没有编码功能。
•内含子上发生的突变不能影响蛋白质的结构,所以其突变往往对生物体是没有影响的;•但也有例外,例如一些发生在内含子上的突变可通过抑制外显子的相互剪接阻止信使RNA的产生。
•利用结构基因的特殊DNA限制片段作为探针,我们可以检测基因组中与之有亲缘关系的序列,结果表明一个基因的外显子常与其他基因的外显子有亲缘关系。
•两个相关基因内含子之间的亲缘关系远远不如其外显子之间的亲缘关系紧密。
•这是因为在进化过程中,相关基因的内含子比外显子变化得更快。
●Splitting gene 概念的相对性a)Intron 并非“含而不露”Yeast 细胞色素b基因Intron II 编码成熟酶b)Exon 并非“表里如一”人类尿激酶原基因Exon I 不编码氨基酸序列c) 并非真核生物所有的结构基因均为splitting geneHistone gene family干扰素Yeast 中多数基因(ADH…)第4节基因及基因组的大小与C值矛盾•由于割裂基因的存在,人们认识到基因比实际编码蛋白质的序列要大得多。
•外显子的大小与基因的大小没有必然的联系。
•不同种类的生物体中外显子的大小并没有明显的不同,基因可能是由一些小的、编码较小的独立蛋白质分区的单位在进化过程中加合起来的。
●基因的大小取决于它所包含的内含子的长度•内含子之间有很大不同,它们的大小从200个碱基对左右到上万个碱基对。
在一些极端的例子里,甚至有50- 60 kb的内含子。
•由于基因的大小取决于内含子的长度和数目,导致酵母和高等真核生物的基因大小有很大的不同。
•大多数酵母基因小于2 kb,很少有超过5 kb的。
•与此相反,在高等的真核生物中,开始出现长的基因,蝇类和哺乳动物基因很少小于2 kb,大多数长度在5~100 kb之间。
•但当基因的长度大到一定程度后,DNA的复杂性与生物体的复杂性之间开始失去必然的联系。
•例如虽然属于同一个门,果蝇细胞的DNA总量较小而家蝇细胞的DNA总量却是它的6倍。
●基因组•狭义:单倍体细胞中的全套染色体(人:22条常染色体+ X,Y + 线粒体DNA)。
•广义:一物种的全部遗传物质及其携带的遗传信息。
●基因组大小与C值矛盾•一个单倍体基因组的全部DNA含量总是恒定的。
这是物种的一个特征,通常称为该物种的C值。
•不同物种的C值差异很大,最小的枝原体只有直106bp,而最大的如某些显花植物和两栖动物可达lO11bp。
Range of genome size in different phyla门植物动物真菌等细菌•由图表可见,随着生物的进化,生物体的结构和功能越来越复杂,其C值就越大,例如真菌和高等植物同属于真核生物,而后者的C值就大得多。
这一点是不难理解的,因为结构和功能越复杂,需要的基因产物的种类越多,也就是说需要的基因越多,因而C值越大。
•然而另一方面,随着进一步的进化,生物体复杂性和DNA 含量之间的关系变模糊了,出现了很多令人不解的现象。
一些生物类群基因组大小的变化范围很窄,而另一些类群的变化范围则很宽。
•突出的例子是两栖动物,C值小的可以低至109bp以下,C值大的可以高达1011bp。
而哺乳动物的C值均为109bp的数量级。
人们很难相信不同的两栖动物,所需基因的数量会有100倍的差别,而且两栖动物的结构和功能会比哺乳动物更复杂。
•由于人们无法用已知功能来解释基因组的DNA含量,所以产生了C值矛盾(C value paradox,又称C值悖理)。
•它表现在两个方面:一个方面是,与预期的编码蛋白质的基因的数量相比,基因组DNA的含量过多。
另一个方面是一些物种之间的复杂性变化范围并不大,但是C值却有很大的变化范围。
这些问题的解决有待于进一步的研究。
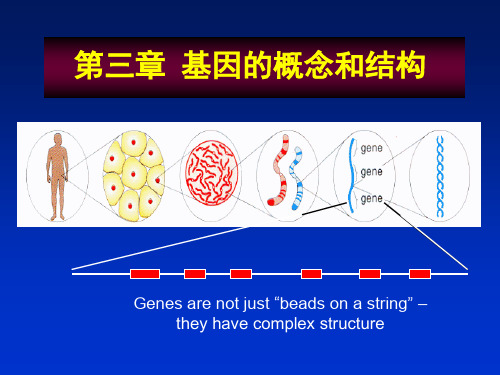
第5节重叠基因●莲人在绿杨津采一玉漱声歌新阙●采莲人在绿杨津,在绿杨津一阙新;一阙新歌声漱玉,歌声漱玉采莲人。
一、原核生物的重叠基因 (overlapping gene)•在细胞基因中,一般一段DNA序列只以三种蛋白质可读框的一种被阅读,但是在一些病毒或线粒体基因中,两个邻近的基因以一种巧妙的方式发生重叠,并以不同的可读框被阅读并表达,因此一段相同的DNA序列可以编码两个非同源蛋白质。
φXl74的DNA序列组织上有重叠基因(overlap-ping gene)和基因内基因重叠基因有以下几种情况:①一个基因完全在另一个基因内部如:B和A*E和D其读码结构互不相同重叠基因基因内基因部分重叠基因一个碱基重叠二、真核生物的重叠基因•通常割裂基因的每个外显子编码一段单一的氨基酸序列,对应于整个蛋白质上的相应部分,而内含子不在最终的蛋白质产物中表达,二者的作用是迥然不同的。
但是有些基因中内含子和外显子的定义是相对的,与它表达的途径有关。
•在这些基因中,选择性的基因表达模式引起了外显子连接途径的转变。
•一个特定的外显子可能选择性地与不同的外显子连接形成信使RNA 。
•这种选择性形式产生的两种蛋白质中,一部分相同而其他部分不同。
一段区域以一种途径表达时作为外显子,而以另一种途径表达时作为内含子。
•因为此时一段DNA序列通常以多种方式起作用,所以不能被简单地称为外显子或内含子。
第6节基因组一、原核生物的染色体基因组二、真核生物基因组一、原核生物的染色体基因组(一)细菌染色体基因组结构的一般特点1. 细菌的染色体基因组通常仅由一条环状双链DNA分子组成,细菌的染色体相对聚集在一起,形成一个较为致密的区域,称为类核(nucleoid)。
类核无核膜与胞浆分开,类核的中央部分由RNA和支架蛋白组成,外围是双链闭环的DNA超螺旋。
染色体DNA通常与细胞膜相连,连接点的数量随细菌生长状况和不同的生活周期而异。
Fig.Typical bacterial cell●E.coli genome is a single double-stranded DNA molecule of 1. 6 mm.●B ut E. coli is only ~ 2 μm in length●D NA is ~ 1000 larger than the size of the cell!●T his is achieved by super-coiling the DNA.●D NA gyrase旋转酶introduces negative-superhelical twists into the DNA. Thedegree of supercoiling of the chromosome is strictly regulated.Fig.The structure of E.coli nucleoid (图大肠杆菌拟核的结构)2. 具有操纵子(trnascriptional operon)结构, 其中的结构基因为多顺反子(polycistron),即数个功能相关的结构基因串联在一起,受同一个调节区的调节。
数个操纵子还可以由一个共同的调节基因(regulatorygene)即调节子(regulon)所调控。
φX174 D-E-J-F-G-H mRNA 外壳蛋白J、F、G、H组装蛋白D 裂解蛋白EE.coli 色氨酸操纵子 9个顺反子 9个酶真核很少,如18s 5.8s 及28s rRNA 基因3. 在大多数情况下,结构基因在细菌染色体基因组中都是单拷贝。
4. 不编码的DNA部份所占比例比真核细胞基因组少得多。
5.具有编码同工酶的同源基因(isogene)例如,在大肠杆菌基因组中有两个编码分支酸(chorismicacid)变位酶的基因,两个编码乙酰乳酸(acetolactate)合成酶的基因。