中性pH条件下化能异养硝酸盐还原菌对亚铁离子的好氧与
水产养殖亚硝酸盐降解实用大全
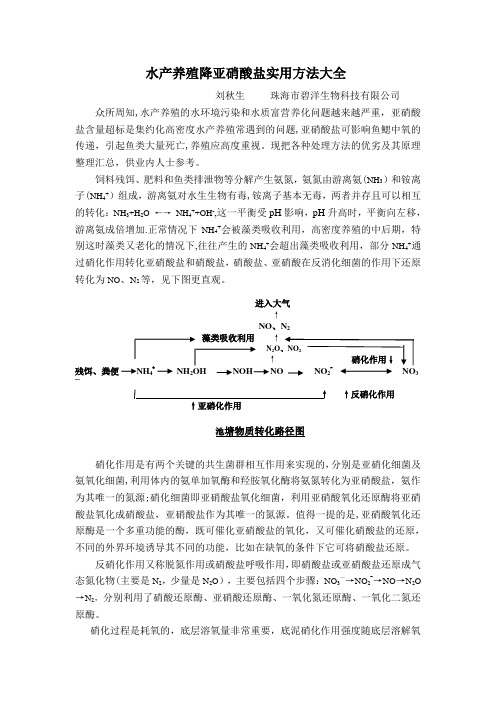
水产养殖降亚硝酸盐实用方法大全刘秋生珠海市碧洋生物科技有限公司众所周知,水产养殖的水环境污染和水质富营养化问题越来越严重,亚硝酸盐含量超标是集约化高密度水产养殖常遇到的问题,亚硝酸盐可影响鱼鳃中氧的传递,引起鱼类大量死亡,养殖应高度重视。
现把各种处理方法的优劣及其原理整理汇总,供业内人士参考。
饲料残饵、肥料和鱼类排泄物等分解产生氨氮,氨氮由游离氨(NH3)和铵离子(NH4+)组成,游离氨对水生生物有毒,铵离子基本无毒,两者并存且可以相互的转化:NH3+H2O ←→NH4++OH-,这一平衡受pH影响,pH升高时,平衡向左移,游离氨成倍增加.正常情况下NH4+会被藻类吸收利用,高密度养殖的中后期,特别这时藻类又老化的情况下,往往产生的NH4+会超出藻类吸收利用,部分NH4+通过硝化作用转化亚硝酸盐和硝酸盐,硝酸盐、亚硝酸在反消化细菌的作用下还原转化为NO、N2等,见下图更直观。
进入大气↑NO、N2↑N2O↑残饵、粪便NH42NOH 23—↑↑反硝化作用↑亚硝化作用池塘物质转化路径图硝化作用是有两个关键的共生菌群相互作用来实现的,分别是亚硝化细菌及氨氧化细菌,利用体内的氨单加氧酶和羟胺氧化酶将氨氮转化为亚硝酸盐,氨作为其唯一的氮源;硝化细菌即亚硝酸盐氧化细菌,利用亚硝酸氧化还原酶将亚硝酸盐氧化成硝酸盐,亚硝酸盐作为其唯一的氮源。
值得一提的是,亚硝酸氧化还原酶是一个多重功能的酶,既可催化亚硝酸盐的氧化,又可催化硝酸盐的还原,不同的外界环境诱导其不同的功能,比如在缺氧的条件下它可将硝酸盐还原。
反硝化作用又称脱氮作用或硝酸盐呼吸作用,即硝酸盐或亚硝酸盐还原成气态氮化物(主要是N2,少量是N2O),主要包括四个步骤:NO3—→NO2-→NO→N2O →N2,分别利用了硝酸还原酶、亚硝酸还原酶、一氧化氮还原酶、一氧化二氮还原酶。
硝化过程是耗氧的,底层溶氧量非常重要,底泥硝化作用强度随底层溶解氧浓度增加而显著增强.硝化细菌比亚硝化细菌对水体pH敏感,硝化细菌进行硝化作用的最适pH范围在8。
关于硝化反硝化的碳源、碱度的计算!

关于硝化反硝化的碳源、碱度的计算!1、硝化细菌硝化反应过程:在有氧条件下,氨氮被硝化细菌所氧化成为亚硝酸盐和硝酸盐。
他包括两个基本反应步骤:由亚硝酸菌(Nitrosomonas sp)参与将氨氮转化为亚硝酸盐的反应;硝酸菌(Nitrobacter sp)参与的将亚硝酸盐转化为硝酸盐的反应,亚硝酸菌和硝酸菌都是化能自养菌,它们利用CO2、CO32-、HCO3-等做为碳源,通过NH3、NH4+、或NO2-的氧化还原反应获得能量。
硝化反应过程需要在好氧(Aerobic或Oxic)条件下进行,并以氧做为电子受体,氮元素做为电子供体。
其相应的反应式为:亚硝化反应方程式:55NH4++76O2+109HCO3→C5H7O2N﹢54NO2-+57H2O+104H2CO3硝化反应方程式:400NO2-+195O2+NH4-+4H2CO3+HCO3-→C5H7O2N+400NO3-+3H2O硝化过程总反应式:NH4-+1.83O2+1.98HCO3→0.021C5H7O2N+0.98NO3-+1.04H2O+1.884H2CO 3通过上述反应过程的物料衡算可知,在硝化反应过程中,将1克氨氮氧化为硝酸盐氮需好氧4.57克(其中亚硝化反应需耗氧3.43克,硝化反应耗氧量为1.14克),同时约需耗7.14克重碳酸盐(以CaCO3计)碱度。
在硝化反应过程中,氮元素的转化经历了以下几个过程:氨离子NH4-→羟胺NH2OH→硝酰基NOH→亚硝酸盐NO2-→硝酸盐NO3-。
2、反硝化细菌反硝化反应过程:在缺氧条件下,利用反硝化菌将亚硝酸盐和硝酸盐还原为氮气而从无水中逸出,从而达到除氮的目的。
反硝化是将硝化反应过程中产生的硝酸盐和亚硝酸盐还原成氮气的过程,反硝化菌是一类化能异养兼性缺氧型微生物。
当有分子态氧存在时,反硝化菌氧化分解有机物,利用分子氧作为最终电子受体,当无分子态氧存在时,反硝化细菌利用硝酸盐和亚硝酸盐中的N3+和N5+做为电子受体,O2-作为受氢体生成水和OH-碱度,有机物则作为碳源提供电子供体提供能量并得到氧化稳定,由此可知反硝化反应须在缺氧条件下进行。
2021年注册公用设备工程师(给水排水)《专业知识考试(下)》真题及详解

2021年注册公用设备工程师(给水排水)《专业知识考试(下)》真题及详解一、单项选择题(共40题,每题1分,每题的备选项中只有1个符合题意)1.反映用水量变化情况的日变化系数,是指哪项?( )A .一年中最高日用水量与平均日用水量的比值B .一年中最高日最高时用水量与最高日平均时用水量的比值C .设计年限内最高日最高时用水量与本年最高日平均时用水量的比值D .一年中最高日最高时用水量与本年平均日平均时用水量的比值【答案】A【解析】在一年之中的最高日供水量和平均日供水量的比值,称为日变化系数K d 。
在一年之中供水最高日那一天的最大一小时的供水量(最高日最高时供水量或用水量)和该日平均时供水量或用水量的比值,称为供水时变化系数K h 。
故选择A 选项。
2.关于枝状给水管网水力计算的说法,正确的是哪项?( )A .多水源枝状管网按顺水流方向进行管段计算流量分配时,可有多个方案B .对于环状管网节点引出的枝状网,在环状管网平差计算时,枝状网的管段流量不受影响C .枝状管网控制点是指供水水源至管网节点水头损失最大的节点D .对于泵站和网后水塔联合供水的枝状管网,最高日最高时的最不利节点也是最大转输时的控制点【答案】B【解析】A 选项,枝状网每一管段只有唯一的流量值。
B 选项,枝状网的流量唯一,不受环状网平差的影响。
C 选项,控制点是水压最难满足的点,一般位于地势较高处或距离二级泵站最远处。
当控制点位于地势较高处,则水源至控制点的水头损失并不是最大。
D 选项,最大转输属于校核时的不利工况。
最大转输时的最不利点一般是水塔处,和高日高时的最不利点不在同一位置。
3.关于输水管(渠)设计计算的说法,正确的是哪项?( )A.当量摩阻计算公式()122=+d s s s ,仅适用于两根不同口径、长度相同且平行布置的并联输水管B .同材质的输水单管由不同管径管段组成,若途中输送流量发生变化时,该输水单管当量摩阻仍为各管径管段摩阻之和C .长度相同、管径不同的两条输水管并联敷设并设有连通管,当一段输水管段损坏时,事故时输水流量的计算与事故管段的摩阻无关D .当泵站的出站输水管发生突然爆管溢流时,输水管特征曲线将向左偏移,对应水泵扬程升高【答案】B【解析】A 选项,当量摩阻计算公式适用任何情况的两根并联管道。
一株地衣芽孢杆菌的异养硝化-好氧反硝化特性研究开题报告

一株地衣芽孢杆菌的异养硝化-好氧反硝化特性研究开题报告研究背景:生物硝化-反硝化过程是地球上氮素循环的重要环节。
细菌在异养硝化过程中将氨或有机氮化合物氧化为亚硝酸盐,再将其硝化为硝酸盐,而在好氧条件下,反硝化菌将硝酸盐还原为亚硝酸盐和氮气。
硝化-反硝化过程对于维护水体、土壤等环境的氮素平衡起着至关重要的作用。
地衣芽孢杆菌是一种多功能细菌,可利用多种有机物和无机物作为氮源和碳源。
已有研究表明,地衣芽孢杆菌具有异养硝化和好氧反硝化的能力。
但是对于其异养硝化和好氧反硝化特性的研究仍不够深入。
研究目的:本研究旨在探究一株地衣芽孢杆菌的异养硝化和好氧反硝化特性,揭示其对氮素循环的贡献,为环境治理提供理论依据。
研究方法:1.分离出一株具有异养硝化和好氧反硝化能力的地衣芽孢杆菌,进行形态学、生理生化特性和分子生物学鉴定。
2.通过动态培养和化学分析法, 研究其异养硝化和好氧反硝化的特性,包括反应条件(温度、pH、营养物质含量等)对异养硝化和好氧反硝化的影响和产氮气的效率。
3.通过比较地衣芽孢杆菌和其他用于氮素循环的细菌的异养硝化和好氧反硝化特性,揭示其在氮素循环过程中的作用。
预期成果:1.明确一株具有异养硝化和好氧反硝化能力的地衣芽孢杆菌的形态学、生理生化特性及分子生物学鉴定结果。
2.揭示地衣芽孢杆菌的异养硝化和好氧反硝化特性,并分析其影响因素和产氮气效率,为环境治理提供理论依据。
3.比较地衣芽孢杆菌和其他用于氮素循环的细菌的异养硝化和好氧反硝化特性,探究其在氮素循环过程中的作用,为氮素循环的控制提供理论基础。
研究意义:本研究对于深入了解地衣芽孢杆菌的异养硝化和好氧反硝化特性,探究其在氮素循环过程中的作用,为氮素循环的控制提供理论基础,也为理解细菌在环境中的生态学过程提供了重要信息。
同时,本研究还为水体、土壤等环境的治理提供了理论基础。
上海应用技术学院(大学)微生物作业2答案

作业2 一、名词解释(选作10题,2*10=20分)1. 抗生素是一类由微生物或其它生物生命活动过程中合成的次生代谢产物或其人工衍生物,它们在很低浓度时就能抑制或干扰它种生物(包括病原菌、病毒、癌细胞等)的生命活动,因而可用作优良的化学治疗剂。
2. 抗代谢物一类在化学结构上与细胞内必要代谢物的结构相似,并可干扰正常代谢活动的化学物质。
3. 灭菌采用任何一种方法,将物体的表面和内部的微生物及其各种芽孢、孢子全部杀死。
一类采用强烈的理化因素使任何物体内外部的一切微生物彻底丧失其生长繁殖能力的措施。
4. 消毒利用某种方法杀死或灭活物质或物体中所有病原微生物的一种措施。
5. 恒浊器根据培养器内微生物的生长密度,借光电控制系统控制培养液流速,以达到菌体密度高、生长速率恒定的连续培养器。
6. 恒化器通过保持有一种生长限制因子的培养液的流速不变,可使微生物始终处在低于其最高生长速率的条件下进行长期生长繁殖的连续培养装置。
7. 兼性厌氧菌是一类以在有氧条件下的生长为主也可在无氧条件下生长的微生物。
特点是在有氧下借呼吸产能,而在无氧条件下可借发酵或无氧呼吸产能;细胞含超氧化物歧化酶和过氧化氢酶。
8. 耐氧菌即耐氧性厌氧菌的简称。
一类可在有氧条件下正常生长却不需氧,而仅借发酵和底物水平磷酸化产能的微生物。
9. 微好氧菌是一类只能在较低的氧分压(103~3*103 Pa)下才能正常生长的微生物。
10. 化学治疗指利用具有高选择毒力即对病原菌具有高度毒力而对其宿主基本无害的化学物质来抑制或杀死宿主体内病原微生物,借以达到治疗传染病的一种措施。
11. 次级代谢微生物在一定的生长时期,以初级代谢产物为前体,合成一些对于该微生物没有明显的生理功能且非其生长和繁殖所必需的物质的过程。
12. 同型乳酸发酵指一分子葡萄糖经EMP途径后仅产生两分子乳酸的发酵。
13. 呼吸作用葡萄糖在好氧和兼性好氧微生物中通过氧化作用放出电子,该电子经电子传递链传给外源电子受体分子氧或其它氧化型化合物生成水或其它还原型产物,并伴随有能量放出的生物学过程称为呼吸作用14. 硝酸盐呼吸又称反硝化作用,是在无氧条件下,某些兼性厌氧菌利用硝酸盐作为呼吸链的最终氢受体,使硝酸盐还原成亚硝酸盐、NO、N2O或N2的过程。
一株铁2f硝酸盐还原菌分离、特性鉴定及其与铁氧化物作用

致谢首先,我要深深感谢恩师陈天虎教授,两年多的时间,在恩师点拨思路、讨论问题下,我渐渐了解了基本的实验方法和研究思路。
陈老师为人和善、严谨、博学、敬业,热衷科学研究,耐心培育学生,可为师之典范。
在我心目中,他永远是我最尊敬的老师。
恩师难觅,师恩难报!回首两年多的师生之情,只有满心的愧疚!感谢课题组其他可亲可敬的老师们,你们是我的第二导师。
特别是在王进、周跃飞、岳正波、陈冬等老师的指点下,我对自己所做的研究有了更直接和深入的了解。
他们的指导犹如春风,唤起我沉睡的思维,使我的研究变得鲜活起来。
感谢李云霞老师对一些实验器材的指导和帮助。
我的毕业与诸位老师的教导点拨密不可分。
感谢课题组王国刚、章红、何光亚、刘畅、王菊、鲍腾、孔殿超等已经毕业和正在读博的师兄师姐们的关心和帮助。
感谢我的同窗姚敦璠、谢芸芸、滕沙沙和杨燕等,我们相互扶持,共同努力,渡过了这一段紧凑而难忘的时光。
感谢正在读研的李碧、程素春、杨露露、韦玲、李明明、罗筱箫、李青、胡雪静等师弟师妹们的帮助和支持,和你们在一起我感到快乐和生机(人太多不能一一叙述,望谅解!)。
在此一并表示由衷的感谢!感谢我的父母、哥哥、姐姐和姐夫的关心和支持。
作为最普通的农民,虽然贫困,但是父母仍以自己勤劳的双手、顽强的意志和乐观的态度培育了一名大学生和两名硕士研究生。
他们不仅给予了我们健康的体魄,也教育了我们健康健全的人格,其过程可谓呕心沥血。
每每想到艰辛的过程,看着他们渐渐年长的身躯,不觉潸然泪下!最后感谢各位答辩组委老师的莅临和指导!谢谢那些给予过我帮助的人,谢谢大家!在今后的工作、学习中我将加倍努力,期望能够取得更多成果回报大家、回报社会!作者:汪国威2013年4月8日目录第1章绪论 (1)1.1 铁(氢)氧化物矿物来源及其环境效应 (1)1.2 铁还原菌与铁(氢)氧化物矿物相互作用及元素耦合 (2)1.3 铁还原菌与铁(氢)氧化物相互作用 (4)1.3.1 铁还原菌概述 (5)1.3.2 铁还原菌还原铁(氢)氧化物电子传递机制 (5)1.3.3 铁还原菌与铁(氢)氧化物作用影响因素 (6)1.3.4 铁还原菌还原铁(氢)氧化物国内外研究现状 (9)1.4 铁还原菌与铁(氢)氧化物矿物作用环境意义 (10)1.5 研究目的、意义及研究内容 (12)第2章铁还原菌分离、鉴定及功能测定 (14)2.1 实验材料、设备及方法 (14)2.1.1 实验仪器 (14)2.1.2 实验材料 (14)2.1.3 实验测试项目及方法 (15)2.2 实验设计及方法 (15)2.3 结果及讨论 (17)2.3.1 分离细菌形态、系统发育和生理生化特征 (17)2.3.2 菌株的生长温度、pH范围和耐氧能力 (18)2.3.3 硝酸盐还原活性 (19)2.3.4 菌株还原铁的活性 (20)2.3.5 铁和硝酸盐共存体系下菌株还原 (21)2.4 讨论 (21)2.5 小结 (26)第3章菌种生长、产氮气条件优化及动力学 (27)3.1 实验材料、设备及方法 (27)3.1.1 实验仪器 (27)3.1.2 实验材料 (27)3.1.3 测试项目及方法 (27)3.2 实验方法 (28)3.3 结果及讨论 (30)3.3.1 温度对微生物生长的影响 (30)3.3.2 pH对微生物生长的影响 (31)3.3.3 pH对还原硝酸盐产氮气的影响 (32)3.3.4 温度对还原硝酸盐产氮气的影响 (33)3.3.5 动力学模型参数 (33)3.4 讨论 (33)3.5 小结 (37)第4章铁还原菌表面性质及絮凝特性 (38)4.1 实验材料、设备及方法 (38)4.1.1 实验仪器 (38)4.1.2 实验材料 (38)4.1.3 测试项目和方法 (38)4.2 实验方法 (39)4.3 结果及讨论 (40)4.3.1 细菌干重和OD600关系 (40)4.3.2 不同时期培养液对絮凝效果的影响 (40)4.3.3 培养液中不同组分对絮凝性能的影响 (41)4.3.4 溶解型和结合型EPS基本成分浓度 (41)4.4 小结 (42)第5章铁还原菌与强磁性铁氧化物矿物作用 (44)5.1 实验材料、设备及方法 (44)5.1.1 实验仪器 (44)5.1.2 实验材料 (44)5.2 实验方法 (45)5.3 结果及讨论 (46)5.3.1 体系中磁化率的变化 (46)5.3.2 体系中矿物变化 (48)5.4 小结 (53)第6章结论和展望 (54)6.1 结论 (54)6.2 展望 (54)参考文献 (56)攻读硕士学位期间发表的论文 (68)插图清单图1-1 异化还原铁耦合碳循环过程示意图 (3)图1-2 希瓦氏菌和土杆菌还原Fe(III)代谢过程示意图 (7)图2-1 菌种形态图像 (18)图2-2 16S rRNA基因琼脂糖电泳图像 (18)图2-3 分离菌种系统发育树状图 (19)图2-4 硝酸根为电子受体转化过程 (20)图2-5 亚铁浓度变化及底物分析 (22)图2-6 Fe(III)和硝酸盐共存时转化过程 (23)图2-7 Fe(III)和硝酸盐共存时代谢路径分析 (24)图3-1 不同温度下Pseudomonas stutzeri CW的生长代谢 (31)图3-2 不同pH值下Pseudomonas stutzeri CW的生长代谢 (32)图3-3 不同pH值下产氮气过程 (34)图3-4 不同温度下产氮气过程 (34)图3-5 不同浓度硝酸盐产氮气过程 (35)图3-6 21.5 mM硝酸盐体系产氮气过程 (36)图4-1 Pseudomonas stutzeri CW 干重与OD600关系 (40)图4-2 不同时期培养液絮凝效果 (41)图4-3 培养液不同成分絮凝效果 (42)图4-4 溶解型及结合型EPS部分成分分析 (42)图5-1 不同条件下铁氧化物磁化率变化 (47)图5-2 两种矿物体系中矿物变化 (49)图5-3 磁铁矿体系矿物与微生物形貌变化 (51)图5-4 磁赤铁矿体系矿物和微生物形貌变化 (53)表格清单表2-1微生物的生理生化特征 (19)表3-1 动力学参数 (36)表5-1 实验设计 (45)第1章绪论1.1铁(氢)氧化物矿物来源及其环境效应1 铁(氢)氧化物来源作为主要的矿体代表,铁是地球上含量最多的元素,在地壳中平均浓度为5%。
两株异养硝化—好氧反硝化细菌的分离、筛选、鉴定和特性研究

两株异养硝化—好氧反硝化细菌的分离、筛选、鉴定和特性研究一、本文概述本文旨在探讨两株异养硝化-好氧反硝化细菌的分离、筛选、鉴定及其特性研究。
异养硝化-好氧反硝化细菌是一类特殊的微生物,能够在好氧条件下进行硝化和反硝化过程,对于氮循环和环境保护具有重要意义。
本文首先通过分离和筛选方法,从自然环境中获取两株具有异养硝化-好氧反硝化功能的细菌,并对其进行初步的生理生化特性分析。
接着,采用分子生物学手段对这两株细菌进行鉴定,明确其分类地位和系统发育关系。
在此基础上,进一步深入研究这两株细菌的生长特性、硝化反硝化性能、以及环境因子对其生长和代谢的影响。
本文的研究结果不仅有助于深入了解异养硝化-好氧反硝化细菌的生物学特性和生态学功能,同时也为该类微生物在环境修复、污水处理等领域的应用提供理论支撑和实践指导。
二、材料与方法为了分离和筛选异养硝化—好氧反硝化细菌,我们从多个不同的生态环境中采集了土壤和水样,包括污水处理厂、河流、湖泊以及农田土壤等。
为了培养和筛选目标细菌,我们使用了多种培养基,包括常规的好氧反硝化培养基和异养硝化培养基。
这些培养基根据细菌的生长特性和需求进行了优化。
实验过程中使用了多种分子生物学试剂,如PCR引物、DNA提取试剂盒等。
同时,还使用了多种仪器,如PCR仪、凝胶电泳仪、微生物培养箱等。
采用稀释涂布法将采集的样品接种到含有相应培养基的平板上,通过观察菌落的形态、大小和颜色等特征,初步筛选出具有异养硝化—好氧反硝化能力的细菌。
通过形态学观察、生理生化特性分析以及分子生物学方法(如16S rDNA序列分析)对筛选出的细菌进行鉴定。
对筛选和鉴定出的细菌进行详细的特性研究,包括生长曲线测定、异养硝化速率测定、好氧反硝化速率测定等。
还研究了环境因子(如温度、pH、碳源和氮源等)对细菌生长和硝化反硝化活性的影响。
实验数据采用统计学方法进行分析,以揭示细菌的生长规律和硝化反硝化特性。
还通过图表等形式直观地展示了实验结果。
微生物合成代谢与分解代谢的联系

在光合系统1中,叶绿素 分子P700吸收光子后被 激活,释放出一个高能 电子。这个高能电子传 递给铁氧还蛋白(Fd), 并使之还原。还原的铁 氧还蛋白将NADP+还原 为NADPH,用以还原 P700的电子来源于光合 系统2。在光合系统2中, 叶绿素分子P680吸收光 子后,释放出一个高能 电子,后者先传递给辅 酶Q,再传递给光合系 统1,使P700还原,失去 电子的P680,靠水的光解 产生的电子来补充。
ED途径 (Entner-Doudoroff pathway)
• ED途径是少数缺乏完整EMP途径的微 生物所具有的一种替代途径,在其他生 物中还没有发现 特点是 葡萄糖只经过4步反应即可快速获得由 EMP途径须经10步才能获得的丙酮酸
• •
• 在ED途径中的关键反应 是2-酮-3-脱氧-6-磷酸葡萄 糖酸的裂解
HMP途径
• 其净效应为:
• 总式为:
6
HMP途径的重要意义
ﯼ为核苷酸和核酸的生物合成提供戊糖-磷酸 ﯼ产生大量的NADPH2形式的还原剂 ﯼ通过EMP途径与本途径在果糖-1,6-二磷酸和 甘油醛-3-磷酸处的连接来加以调剂对戊糖的需 要 ﯼ作为自养微生物固定CO2的中介(Calvin循环) ﯼ由于在反应中存在着C3~C7的各种糖,使具有 HMP途径的微生物的碳源利用范围更广 ﯼ通过本途径而产生的重要发酵产物很多,例如 核苷酸、若干氨基酸
化学渗透假说要点:a 氧化磷酸化过程中,通过呼吸链酶系的作用, 将底物分子上的质子从膜 的内侧传递至外侧,从而造成质子在膜的 两侧分布的不均衡,亦即形成 了质子梯度差,这梯度差是产生ATP 能量的来源。 b 通过ATP酶的作用,把质子从膜的外侧再运到膜的内侧时,一方 面消除了质子梯度差,同时合成了ATP。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
中性pH条件下化能异养硝酸盐还原菌对亚铁离子的好氧与厌氧氧化Arch Microbiol 1998, 169:159-165 IF= 1.975摘要:百分之九十的厌氧富集培养都出自于一些淡水沉积物试样以及一些海洋沉积物,在pH 7.2、30℃条件下下,沉积物中的亚铁离子在硝酸盐和有机辅助物的矿物媒介中被氧化。
厌氧硝化的亚铁氧化是一个生物过程。
从苦咸水沉积物中分离出的一种微生物(HidR2,运动型不产芽孢的革兰氏阴性棒状菌),进一步研究在醋酸盐环境中对亚铁离子的氧化作用。
在微量醋酸盐(0.2~1.1 mM)环境中,pH 7.2、30℃条件下,HidR2菌能厌氧和好氧氧化0.7~4.9 mM的亚铁离子,其用于生长的能量来自于亚铁离子的厌氧硝化氧化,铁氧化比率是一个常数,取决于醋酸盐的量的给予。
中性条件下硝酸盐厌氧氧化亚铁离子的能力似乎是嗜温反硝化细菌的共有特性。
由于依赖硝酸盐的铁离子氧化关闭沉积物缺氧区的铁循环、铁离子的好氧氧化促进含氧区内的二价铁再氧化为三价铁,这两个过程都增加了铁离子在自然沉积物的有机质循环转化中作为过渡电子载体的重要性。
关键词:铁氧化亚铁离子高铁离子硝酸还原作用沉积物1 引言铁元素是地壳中含量最丰富的元素之一,也是含量第二丰富的金属元素。
由于其低水溶性,铁氧化物在水环境中通常以沉淀形式存在,如复杂的非晶形或晶体结构(Cornell and Schwertmann 1996)。
三价铁离子能够被异养菌还原为二价铁离子(Lovley 1991),在有氧条件下,亚铁离子可以在低pH值中被嗜酸细菌如氧化亚铁硫杆菌再次氧化(Blake et al. 1993),或者在近中性环境里,被铁锈色披毛菌氧化(Hallbeck et al. 1993),这两种细菌都是从氧化还原反应中获取能量而生长。
Fe3+/Fe2+的标准氧化还原电位为+770 mV,但是在自然环境中其实际的氧化还原电位很大程度上取决于pH值的变化(Widdelet al. 1993)。
在pH 7的碳酸氢盐环境下,FeOOH/FeCO3的氧化还原过渡电位E值约为+200 mV。
在较低的氧化还原电位下,亚铁离子也能够充当给电子体,从而在厌氧环境中发生还原反应,最近有研究者可以分离和描述能利用亚铁离子作为电子供体的不产氧光合细菌(Widdel et al. 1993; Ehrenreich and Widdel 1994)。
+200 mV下的释放电子也可以用作微生物的硝酸盐异化作用,最近报道了一种嗜温细菌(Straub et al. 1996)和一种极端嗜热的古细菌(Hafenbradl et al. 1996)都具有这种新陈代谢功能。
这一过程将铁循环和氮循环联系上,因此也显示了微生物厌氧环境中铁离子作为电子受体的重要性。
本文主要研究在兼性厌氧硝酸还原菌辅助作用下亚铁离子的氧化过程。
2 材料和方法2.1 微生物的来源由于采取加富培养,各种沉积物试样被作为接种源,有淡水源(V orsee,Mittlerer Buchensee,and Lake Constance,Germany)、海水源(Venice, Italy; Ameland and Groningen, Netherlands)、盐水源(Hiddensee,Germany)以及市政污水处理中使用的活性污泥源(Ko nstanz,Germany)。
2.2 培养介质和培养方法对于淡水和海水中细菌的加富培养,使用在缺氧条件下的碳酸氢盐缓冲溶液作矿质媒介,且加入1mM的硫酸盐作为硫源,不需加还原剂,NaCl和MgCl2的浓度则与微生物本身的环境相适应,淡水中NaCl和MgCl2 · 6H2O的量分别为1.0 g和0.4 g,盐水中为7.0 g和1.0 g 海水中则为20.0 g和3.0 g。
经高压灭菌,N2/CO2(80:20, v/v)冷却后,每升溶液中分别加入1 ml微量元素溶液SL9(Tschech and Pfennig 1984)和7-维生素溶液(Widdel and Pfennig 1981),并将pH调至7.2,。
而对于需氧培养实验,培养基仍和上述方法一样,所不同的是缓冲溶液由羟乙基哌嗪乙硫磺酸所替代。
2.3 富集与分离将50ml 高压灭菌后的培养基装入120ml规格的血清瓶中,用丁基橡皮塞密封好,在缺氧条件下往里加入约5ml的接种体。
并无菌地向基底分别注入浓度为4mM 硫酸亚铁和5mM 硝酸盐。
每个接种体均在pH为6.5、7.0、8.0,温度为16和30℃下富集培养。
培养周期为2~4周,以棕色锈状沉淀形成为一个周期。
苦咸水培养的细菌的纯化是用的琼脂稀释法(Pfennig and Trüper 1981):60℃下向25ml 的试管中加入3ml 3%(w/v)的琼脂溶液,再加入8 mM 硫酸亚铁溶液,小心混匀,然后,加入7ml 42℃预热的含5 mM硝酸盐培养基,得到的亚铁离子最终浓度为4 mM。
每根稀释后的试管都经过密封、接种、水域冷凝以及N2/CO2 (80:20, v/v)气体吹扫,最后置于避光的30℃恒温箱中培养。
在进入液体培养基前,微生物都是在琼脂上生长繁殖的。
淡水和海水微生物的纯培养在平板基上分离。
平板基的基质也是上述培养基加入0.01M 丙磺酸作为缓冲溶液、1.5%(w/v)的琼脂、5mM 硝酸盐和3 mM 醋酸盐。
在30℃,N2/CO2(80:20, v/v)气氛下进行避光平板接种。
培养好的菌种转移到呈有淡水培养基的血清瓶(60ml)中。
只有在瓶子中进行亚铁离子氧化的微生物才在琼脂板上显示多于两条条纹从而得以纯化,并且将其转移到液体培养基中培养。
在含有矿物质元素和0.2%的酵母膏的培养基中培养后的微生物在显微镜下进行纯化检查。
培养期间需定期用显微镜检查微生物是否被污染。
检测鞭毛是否形成,使用的方法是布雷登-戈登堡银侵染法。
2.4 增长实验细菌在装有300ml培养基的瓶(500ml)中培养,未接种的空白试验瓶在相同条件下培养至少两个月,但是始终未见到有能氧化二价铁离子的微生物产生。
经过震荡后的样品放入具塞瓶中,用N2/CO2 (80:20, v/v)气预吹,然后转移到盐酸或者磷酸缓冲溶液中准备下一步分析。
为保证在氧气梯度管中的生长实验,将4ml 缺氧培养基装入22ml试管中,培养基中含有1.5% (w/v)琼脂糖、15 mM 亚铁离子、1 mM醋酸盐。
在氮气保护下,等第一层凝结后,再加入第二层培养基(0.5% (w/v)琼脂糖),将微生物接种在这一层上。
等到第二层氮气保护下凝固好以后再往上面加入1ml原培养基不含琼脂糖,为的是防止第二层琼脂糖的干结。
试管用铝盖轻轻盖上,垂直放入暗箱中在30℃下,振荡培养(80rpm)。
以上所有试验重复一遍2.5 分析方法亚铁离子的量用亚铁嗪分光光度法来测定(Stookey 1970)。
总铁离子的浓度测定也是用同样的方法即加入盐酸羟胺将所有的铁离子都还原成亚铁离子态(Stookey 1970)。
三价铁离子则有上述二者差值得到。
为得到亚硝酸盐的含量,样品与一体积500mM的磷酸缓冲溶液在厌氧环境下混合,并在室温下反应15分钟,然后在离心机上用最大转速离心5分钟。
用1ml 1M的盐酸将其又恢复粒子态。
这样处理可以避免亚硝酸盐在分析前和分析过程中被亚铁离子所氧化。
如果试样中含有超过1mM的亚硝酸盐,则这个过程必须重复至少两次。
由于铁离子分析是在梯度管中进行的,所以琼脂糖要用刀片切成2mm后的薄片并加入1ml 5M的盐酸。
在40℃水浴中静置15分钟后,薄片已被酸化成液体,铁离子在上述过程中被测量完毕。
硝酸盐和亚硝酸盐用带有格鲁姆阴离子交换柱(Grom, Herrenberg, Germany)的高效液相色谱来进行定量分析,在220nm下进行吸收检测。
为了抑制铁离子和亚硝酸盐的相互反应,并且为保护色谱柱,含铁样品中得加入磷酸缓冲溶液,上清液用于硝酸盐亚硝酸盐的检测。
这种方法的对亚硝酸盐的检出限大约是30 μM。
醋酸盐用气相色谱(GC6000 Vega Series 2; Carlo Erba, Milan, Italy)分析,使用填充柱(2 m×2mm; 60/80 Carbopack C/0.3% carbowax 20 M/0.1% H3PO4),火焰离子检测器。
为了保护柱子,醋酸盐试样也按上述方法准备。
氧化二氮也用上述气相色谱分析,使用的填充柱[2 m×2 mm; 60/80 Carbosieve SII (Supelco, Bellefonte, Pa., USA)]和检测器(热导池检测器)不一样。
根据Hall 和Aller (1992),铵盐用流动注射分析。
蛋白质由以下分析:测定含铁样品中蛋白质的含量,需要准备含有试样中相同铁量的蛋白质标样,为的是测定蛋白质化学分析中与铁离子的可能反应。
由于几种HidR2菌所对应的不同的光密度的蛋白质含量与干重呈一种相关关系,根据这种关系,可以来计算蛋白质的干重。
梯度管中的氧含量用三维显微操作器驱动氧微电极而测定。
3 结果3.1 富集培养在7~10天的培养下,除了威尼斯海峡采集的沉积物以外,所有的富集培养都形成了棕色锈状沉淀。
通过化学分析,这些沉淀都被确定为铁的氢氧化物,而在未富集培养的培养基里都未发现亚铁的氧化作用。
经过三到四次转移培养,需加入醋酸盐或者琥珀酸盐作为有机补给物以维持亚铁离子的氧化作用环境。
只有从格罗宁根市海洋沉淀物中富集培养的培养基不用加入补给物而能不断氧化亚铁离子,即使重复转移一年以上也能进行反应。
在比较所有富集培养后发现,无论是在16℃或30℃还是pH为6.5或8,亚铁离子的转化率和潜伏程度都没有明显的改变。
3.2 纯培养的特征从所有能发生亚铁离子氧化反应的富集培养中来看,纯培养独立的显示了亚铁离子厌氧氧化过程中醋酸盐是作为有机补给物而硝酸盐则是作为电子受体。
从波罗的海沉积物中分离出来的HidR2菌种将被作为进一步研究的对象。
图1HidR2菌在醋酸盐和硝酸盐厌氧生长中的相差显微镜照片(图中横杠表示10微米)HidR2菌属于革兰氏阴性,不产孢子的杆菌,大小为3~6×1μm(图1),单极鞭毛运动型,对氧化酶和过氧化氢酶产生极性。
通过相差显微镜观察到细胞两端有微小深色包涵体,用三氯甲烷处理后,这些包涵体消失,可能是由于聚羟基脂肪酸的富集产生。
在过量醋酸盐(>3 mM)中培养后,HidR2菌每一批能产生约10~50个细胞。
在含有亚铁离子、醋酸盐和硝酸盐的液体培养基中,通过4,6-二脒基-2-苯吲哚盐酸染色法,可以发现绝大多数细胞都含有絮状的三氧化铁或者肉眼可见的姜铁矿,但氢氧化铁并非包裹在细胞外部。