长江口外海域微型和小型底栖生物群落结构和时空变化
长江口大型底栖动物群落的演变过程及原因探讨

长江口大型底栖动物群落的演变过程及原因探讨刘录三;郑丙辉;李宝泉;蔡文倩;韩庆喜;林岿璇【摘要】根据近30年来的长江口大型底栖动物群落的历史资料,以及2009年和2010年的现场调查数据,从长江口底栖动物群落结构变化特征出发,分析了其长期演变的过程和规律,旨在阐明底栖动物群落的演变趋势,识别其重要演变时段.同时结合长江口海域近50~60年来的入海径流量和携沙量变化、营养盐和DO等水质特征变化,分析底栖动物群落的变化原因.分析结果表明,长江口底栖生物群落的变化可大体分为3个阶段,第一阶段是20世纪90年代之前,底栖生物群落无论物种数、生物量都维持相对较高的水平;第二阶段发生在20世纪90年代初至2005年,由于受到气候变化和人类活动的综合影响,底栖生物群落的上述指标都有所降低,表明受到自然和人为因素干扰的加剧;第三阶段是2005年之后至现在,由于长江口水域的各项生态环境保护措施加强,底栖生物群落得到一定程度的恢复.通过对长江口海域各种环境因素的分析表明,底栖生物群落变化受到长江口区域生物因素和非生物因素的共同影响,变化趋势也与环境因素的变化比较吻合.%The macrobenthos samples collected from Changjiang Estuary, China in 2009 and 2010 were i-dentified to clarify the current state, and the historical data obtained from essentially the same sampling area were analyzed to get the long-term trends and their response to environmental changes over the past 30 years. The average total species number exhibited an obvious fluctuation over this 30 year period, which includes three periods, e. g. , period one-before 1990s, the total species number maintained relatively high value, period two-from 1990s to 2005, the total species number decreased markedly, and period three-after 2005, the total species number increasedrapidly. The average biomass and the abundance also had similar trends over the last 30 years. The trends of species composition of macrobenthos community was that some long-lived, larger sized, stress intolerant resident species had been replaced by some opportunistic, shortlived, small sized taxa, especially some opportunistic polychaeta species, which indicated the macrobenthos community of the study area was unhealthy and unstable. The results of MDS ordination also approximately coincide with the above results. The integrated impacts both from climate changes and from anthropogenic disturbances, such as aquaculture, coastal land reclamation and sewage discharge were the triggers for these long-term changes.【期刊名称】《海洋学报(中文版)》【年(卷),期】2012(034)003【总页数】12页(P134-145)【关键词】大型底栖动物;群落演变;长江口【作者】刘录三;郑丙辉;李宝泉;蔡文倩;韩庆喜;林岿璇【作者单位】中国环境科学研究院国家环境保护河口与海岸带环境重点实验室,北京100012;中国环境科学研究院国家环境保护河口与海岸带环境重点实验室,北京100012;中国科学院烟台海岸带研究所,山东烟台264003;中国环境科学研究院国家环境保护河口与海岸带环境重点实验室,北京100012;北京师范大学水科学研究院,北京100875;中国科学院烟台海岸带研究所,山东烟台264003;中国环境科学研究院国家环境保护河口与海岸带环境重点实验室,北京100012【正文语种】中文【中图分类】P714十.5底栖生物作为海洋生态系统中的重要组成部分,在海洋食物网和沉积物-水层界面的生物地球化学循环过程中起着重要的作用。
长江口张网鱼类群落结构特征及月相变化

长 江 口张 网鱼 类群 落 结构 特 征 及 月 相 变 化
史 赘荣 , 晁敏 , 沈 新 强
( 1 .中 国水 产 科 学 研 究 院 东 海 水 产研 究 所 , 上海 2 0 0 0 9 0 )
摘 要 :为 了解长 江 口鱼类群 落结构 的现状及 变化特征 , 用 张 网作业对 长 江 口 2 0 1 0年 5月 至 2 0 1 1年 4
41 9 3 .2 O1 4 . 02 . 0 09
ห้องสมุดไป่ตู้
S h i Yu n r o n g,C h a o Mi n,S h e n Xi n q i a n g .Ch a r a c t e r i s t i c s a n d mo n t h l y v a r i a t i o n s o f s e t n e t f i s h c o mmu n i t y s t r u c t u r e i n t h e C h a n  ̄i a n g
群落 生态学研 究 也是 国际热 点 研究 之 一 。关 于 长 江 口鱼类 群落生态 学 的研究 , 有 学者侧 重= F 鱼类资 源
收稿 日期 : 2 0 1 2 — 1 2 — 2 9 ; 修 订 日期 : 2 0 1 3 — 0 4 — 2 8 。
关键词 : 长江 口; 鱼类群 落结构 ; 时间变化 ; 环境 因素
中 图分 类 号 : ¥ 9 3 1 . 1 文献 标 志 码 : A 文章 编 号 : 0 2 5 3 — 4 1 9 3 ( 2 0 1 4 ) 0 2 — 0 0 8 1 — 1 2
1 引 言
河 口对栖息 在其 中的很 多生 物种 类有 着 重要 的 作用 , 例如 , 河 口不 仅是 很 多经 济鱼类 的育 幼场 口 - 。 ] ,
横沙东滩大型底栖动物多样性及其群落结构特征

(1Shanghai Investigation, Design & Research Institute Co., Ltd., Shanghai 200120, China; 2Yangtze Eco-Environment Engineering Research Center, China Three Gorges Corporation, Beijing 100038, China;
Y=ni / N×fi
(1)
d =(S - 1)/ log2 N
(2)
J = H′/ log2S
(3)
H′ = -∑(ni /N)× log2(ni /N)
(4)
式(2—4)中,S 为总种数,N 为总丰度,ni为第 i 种
的丰度。
- 73 -
资源·环境·植保
安徽农学通报 2024 年 04 期
1.4.3 群落结构 采用 Primer 5.0 软件进行聚类 分析(Cluster)和丰度/生物量曲线(ABC 曲线)分析, 将调查海域大型底栖动物丰度数据对数转化,得到
具有重要意义[5]。 横沙东滩位于横沙岛东端,面积约 52 km2,是长
江口发育较好的浅滩之一,与长兴岛和九段沙构成 了长江口的三大浅滩。随着河道整治与促淤圈围工 程的开展,横沙东滩的湿地面积逐年增加。近年来, 对长江口潮间带大型底栖动物的研究集中在九段沙 和崇明岛[6-10],而针对横沙东滩大型底栖动物多样性 和群落结构的研究相对较少。吕巍巍等[11]对围垦后 横沙东滩大型底栖动物群落结构进行了研究,发现 围垦导致该区大型底栖动物群落结构发生了明显改 变。为进一步了解横沙东滩围垦后潮间带大型底栖 动物群落结构和多样性,本研究于 2019 年 10 月、 2020 年 4 月分别对研究区成陆区和自然潮滩的大型 底栖动物开展了调查,为横沙东滩大型底栖动物深 入研究提供参考。
长江口新生沙洲底栖动物群落组成及多样性特征

2 研 究 区域
九段 沙是 由长 江水流 携带 的大量泥 沙在 人海 口受海水 、 汐的作用 沉积 而 戚 .它 由上沙 、 潮
收藕日期 : 加∞ 一l — 8 修订 日期: 0 1 3 0 2 0; 2 0 —0 —2 基 盘项 目: 教育部科技研究 20 年重点资助项 目( 0 1 ) 上海市重 点学科资助 项目(9 0 0 ) 00 K 20; B 7 32 作者■升 : 中(9 3 )男, 寰 1 6 一 , 四川省万振市^, 副教授 博士 . 从事生态 系统生态学和环境 生态学研兜
九段沙是长江口现代发育过程 中产生的一个新 生沙洲, 是继崇明岛、 长兴岛和横沙岛之后 的叉一成陆冲积沙洲, 是一块重要的河 口沙洲型滨海湿地【 . 3 九段沙受人类干扰较少, 】 其生物
资源保持 着天 然状态 作 为世界 上特 大 型河 口的新 生沙 洲 . 九段 沙 生物 的 出现 和演 替规 律 具 有其独特 性. 是进 行 生态系统 演替 和 生物 多样性 研究 的理 想基 地 .九段 沙 的底栖 动 物 群落及 生物多 样性 不仅与河 口理化 环境 密切 相关 , 且 明显受 沙洲 自然演变 的 影响 .我 们 曾经对 九 而 段沙底栖 动物 的 生态学特征进 行过 初步研 究Lj但 对 于九段 沙底 栖 动物 的多样性 特征 尚未 见 4. 报道 .本文根 据 19 99年 3月至 2 0 00年 9月 的实地 调研 , 九段 沙 底栖 动物群 落种 类 组 成及 对 多样性 特征进 行 了探 讨 . 本工作 不仅 对于 长江 口生物 多样性 的基 础研究 以及 生 态系统 的演 替 研究具 有意 义. 而且 可以为 长江 口沙 洲岛屿的 资源开 发和 自然保护 , 以及探 水航道 的建设 与治
【浙江省自然科学基金】_群落结构_期刊发文热词逐年推荐_20140811
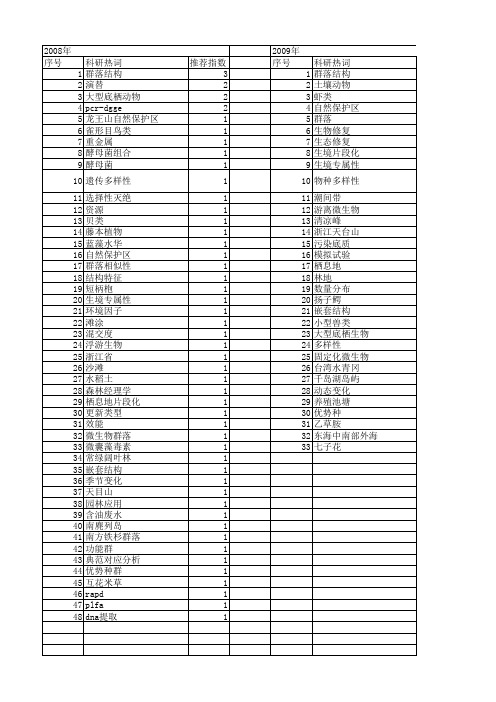
53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71
2011年 科研热词 推荐指数 群落结构 3 微生物群落结构 2 长江口 1 西瓜嫁接 1 茶园 1 聚糖菌(gaos) 1 聚磷菌(paos) 1 聚磷菌 1 群落多样性 1 群落丰富度 1 细菌群落 1 细菌 1 种类组成 1 社团结构 1 磷细菌 1 相关统计分析 1 电泳图谱 1 生成因子 1 生境片段化 1 甜槠 1 环境因子 1 牵制控制 1 溪流 1 渗滤液 1 浮游动物 1 浙江天台山 1 氮细菌 1 氨氮 1 榨菜 1 椒江口 1 森林群落结构 1 森林景观效果 1 根际微生物 1 枫香属 1 条件优化 1 景观生态学 1 时空分布 1 无脊椎动物 1 微生物多样性 1 强化生物除磷(ebpr) 1 强化生物除磷 1 异养细菌 1 常绿阔叶林 1 季节性变化 1 外貌 1 复杂网络 1 填埋垃圾生物反应器 1 堆肥 1 土壤节肢动物 1 土壤动物 1 变性梯度凝胶电泳(dgge) 1 反硝化细菌 1
推荐指数 4 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51
科研热词 群落结构 土壤动物 虾类 自然保护区 群落 生物修复 生态修复 生境片段化 生境专属性 物种多样性 潮间带 游离微生物 清凉峰 浙江天台山 污染底质 模拟试验 栖息地 林地 数量分布 扬子鳄 嵌套结构 小型兽类 大型底栖生物 多样性 固定化微生物 台湾水青冈 千岛湖岛屿 动态变化 养殖池塘 优势种 乙草胺 东海中南部外海 七子花
游泳动物对长江口新生盐沼湿地潮沟生境的利用

游泳动物对长江口新生盐沼湿地潮沟生境的利用3全为民 倪 勇 施利燕 陈亚瞿33(中国水产科学研究院东海水产研究所农业部海洋与河口渔业重点开放实验室,上海200090)摘 要 利用长袋网(fyke net )采集了长江口新生盐沼湿地潮沟内的游泳动物,分析了该类盐沼湿地的鱼类栖息地利用。
结果表明,3次调查共记录到游泳动物20种,其中鱼类15种,虾蟹类5种。
生态类群主要以淡水性种类(10种)和河口性种类(6种)为主,洄游性种类(3种)和海洋性种类(1种)较少。
从数量组成来看,游泳动物群落主要由安氏白虾(3514%)、棕刺虾虎鱼(1717%)、贝氏餐(1717%)、长蛇鮈(912%)和日本沼虾(416%)等少数几个物种占优势。
这些游泳动物大多为稚幼个体,表明长江口新生盐沼湿地是许多鱼类和甲壳动物的重要育幼场所。
关键词 游泳动物;盐沼湿地;生境;利用;长江口中图分类号 Q178.1 文献标识码 A 文章编号 1000-4890(2009)03-0560-05Utili za ti on of ti da l creek hab it a t by the nekton s i n a newly for m ed s a lt marsh a t Yangtze R i ver estuary .QUAN W ei 2m in,N I Yong,SH IL i 2yan,CHEN Ya 2qu (Key and O pen L aborato 2ry of M arine and Estuarine F isheries,M inistry of A g ricultu re,East China S ea F isheries R esea rch Institute,Ch inese A cade m y of F isheries S ciences,Shanghai 200090,China ).Chinese Journal of Ecology ,2009,28(3):560-564.Abstract:The nekt ons in an intertidal creek of a ne wly for med salt marsh at Yangtze R iver estu 2ary were sa mp led by Fyke net t o assess its habitat utilizati on of the salt marsh .A t otal of 20nek 2t on s pecies were recorded in three investigati ons,including 15fishes and 5crustaceans .Among the nekt ons,fresh water s pecies (10s pecies )were the most abundant ecol ogical gr oup s,foll owed by estuarine (6s pecies ),m igrat ory (3s pecies ),and coastal s pecies (1s pecies ).I n ter m of numeric abundance,the nekt ons were highly dom inated by Exopa lae m on annanda lei (35.4%),A can thogobius luridus (1717%),He m iculter leucisculus (1717%),Saurogobio dum erili (912%),and M arobrachium n ipponensis (416%).Most of the nekt ons sa mp led in this study were juveniles,suggesting that the ne wly f or med salt marsh was the i m portant nursery habitat for many fishes and crustaceans .Key words:nekt on;salt marshes;habitat;utilizati on;Yangtze R iver estuary .3上海青草沙原水工程有限公司项目、中央级公益性科研院所基本科研业务费专项资金(中国水产科学研究院东海水产研究所)资助项目(2007M03)。
长江口海域生态环境状况及保护对策

第33卷㊀第5期2020年5月环㊀境㊀科㊀学㊀研㊀究ResearchofEnvironmentalSciencesVol.33ꎬNo.5Mayꎬ2020收稿日期:2020 ̄02 ̄04㊀㊀㊀修订日期:2020 ̄03 ̄21作者简介:王孝程(1990 ̄)ꎬ男ꎬ黑龙江哈尔滨人ꎬ工程师ꎬ博士ꎬ主要从事海洋生态学研究ꎬxcwang@nmemc.org.cn.∗责任作者ꎬ李宏俊(1982 ̄)ꎬ男ꎬ辽宁丹东人ꎬ研究员ꎬ博士ꎬ主要从事海洋生态学研究ꎬhjli@nmemc.org.cn基金项目:自然资源部海洋灾害预报技术重点实验室开放基金项目(No.LOMF1805)ꎻ国家海洋环境监测中心博士科研启动经费项目SupportedbyOpenFoundationofKeyLaboratoryofMarineHazardsForecastingꎬMinistryofNaturalResourcesꎬChina(No.LOMF1805)ꎻDoctoralFoundationofNationalMarineEnvironmentalMonitoringCenterꎬChina长江口海域生态环境状况及保护对策王孝程1ꎬ2ꎬ解鹏飞1ꎬ李㊀晴1ꎬ张金勇1ꎬ李宏俊1∗1.国家海洋环境监测中心ꎬ辽宁大连㊀1160232.自然资源部海洋灾害预报技术重点实验室ꎬ北京㊀100081摘要:为加快推进长江口海域的生态环境保护和修复工作ꎬ结合长江经济带大保护ꎬ系统总结分析了近20年长江口环境质量和生态监控区的监测结果.结果表明:①长江口海域生态系统长期处于亚健康状态.②长江径流总量呈现波动变化ꎬ年均流量无明显的变化ꎬ而长江口海域海水环境状况一直较差.③营养盐污染严重ꎬ主要污染物是无机氮和活性磷酸盐ꎻ浮游生物和底栖生物群落结构不稳定ꎬ存在生境破碎化严重㊁外来生物入侵㊁赤潮频发㊁低氧区等诸多生态问题.为加强长江口海域生态环境的保护与修复ꎬ建议:①加强顶层设计ꎬ推进落实陆海统筹ꎻ②科学规划临港产业布局ꎬ加强涉海产业的污染管理ꎻ③加强污染物入海排放管控ꎬ提升海洋环境保护意识ꎻ④保障海洋生态建设资金ꎬ强化海洋生态保护与建设.关键词:长江口ꎻ生态环境ꎻ变化趋势ꎻ生态问题ꎻ保护对策中图分类号:X321㊀㊀㊀㊀㊀文章编号:1001 ̄6929(2020)05 ̄1197 ̄09文献标志码:ADOI:10 13198∕j issn 1001 ̄6929 2020 03 29EcologicalEnvironmentoftheYangtzeEstuaryandProtectionCountermeasuresWANGXiaocheng1ꎬ2ꎬXIEPengfei1ꎬLIQing1ꎬZHANGJinyong1ꎬLIHongjun1∗1.NationalMarineEnvironmentalMonitoringCenterꎬDalian116023ꎬChina2.KeyLaboratoryofMarineHazardsForecastingꎬMinistryofNaturalResourcesꎬBeijing100081ꎬChinaAbstract:InordertopromotetheprotectionandrestorationoftheYangtzeestuaryundertheprotectionstrategyofYangtzeRiverEconomicBeltꎬwesystematicallyanalyzedthemonitoringdataoftheYangtzeestuarymarineecologicalmonitoringprogramsinrecent20years.Theresultsindicatedthattheecosystemwasinasub ̄healthlong ̄termstate.Thetotalrunofffluctuatedwhiletherewasnoobviouschangeintheannualaverageflow.Howeverꎬitisnoteworthythattheconditionofseawaterwaspoor.Nutrientsweremainpollutants(i.e.inorganicnitrogenandphosphate).Manyotherproblemssuchasbiodiversitylossꎬdamagedhabitatꎬalieninvasionꎬfrequentredtideꎬandlow ̄oxygenzonesalsoexist.Werecommendimprovingtheprotectionandrestorationoftheecologicalenvironmentfromthefollowingaspects:(1)Strengthenthetop ̄leveldesignandpromotecoordinatedlandandseadevelopmentꎻ(2)Plananddesignthelayoutofport ̄vicinityindustryscientificallyandimprovethepollutionmanagementofsea ̄relatedindustriesꎻ(3)Tightenthecontrolofpollutantdischargeandincreasetheawarenessofmarineenvironmentalprotectionꎻ(4)Ensuresufficientfundsformarineecologicalconstructionandstrengthenitsprotectionandconstruction.Keywords:Yangtzeestuaryꎻecologicalenvironmentꎻhealthconditionꎻecologicalproblemꎻprotectioncountermeasure㊀㊀长江口是世界第三大河口ꎬ生态环境状况特殊[1].长江口海域在海洋水团的共同作用下ꎬ水温状况复杂多变ꎬ营养盐丰富ꎬ生产力高ꎬ磷酸盐㊁硝酸盐和硅酸盐显著高于我国其他河口海域[2 ̄3].营养盐含量从近海向河口区逐渐递增ꎬ导致河口海域成为高生产力区[4 ̄5].长江径流带来的营养物质ꎬ孕育了大量的浮游生物和滩涂植物ꎬ为水生动物和底栖生物提供了充足的食源[6 ̄7]ꎬ是众多溯河性和降河性长途洄游性物种ꎬ如中华鲟(Acipensersinensis)㊁鳗鲡(Anguillajaponica)等鱼类的必经通道[8 ̄11]ꎬ是我国凤鲚(Coiliamystus)和中华绒螯蟹(Eriocheirsinensis)的最主要产卵场之一ꎬ还是珍稀物种中华鲟幼鲟的集中分布区[12 ̄17].滩涂湿地是鸟类亚太迁徙路线中的重要驿站[18].但是随着人类干扰的不断增多ꎬ长江口海域的㊀㊀㊀环㊀境㊀科㊀学㊀研㊀究第33卷生态环境状况也受到了严重影响ꎬ生境破碎化严重ꎬ生态系统长期处于亚健康状态ꎬ其保护和修复工作亟需更高质量的推进.中共中央㊁国务院高度重视长江生态环境保护工作ꎬ推动长江经济带发展是党中央作出的重大决策ꎬ是关系国家发展全局的重大战略.随着长江大保护的持续推进ꎬ长江经济带地表水环境质量呈好转趋势ꎬ总体优于全国平均水平ꎬ并且生态环境质量正逐渐好转ꎬ保护和修复成果显著.而海纳百川ꎬ长江最终于崇明岛以东汇入我国东海ꎬ海洋是其保护成效的最终体现者之一ꎬ长江口作为重要的陆海连接区域ꎬ是长江保护和修复成效的重要体现者ꎬ所以长江口海域的生态环境质量评价工作对于评估长江保护和修复的成效具有重要意义ꎬ其生态环境状况尤为重要.该研究系统总结了近20年来长江口海域的业务化监测结果ꎬ对生态环境状况及其变化趋势进行了分析ꎬ剖析长江口海域存在的主要生态问题ꎬ并提出了相应的保护修复和管理对策ꎬ以期为长江经济带的保护成效评估提供参考ꎬ为长江口海域的保护和修复工作提供科学依据.1㊀长江口海域生态环境状况及其变化趋势1 1㊀长江口海域水体和沉积物环境1 1 1㊀长江口径流和泥沙特性长江口是我国最大的河口ꎬ近10年来ꎬ长江流域及长三角区域经济发展迅速㊁人口相对集中㊁海上倾废㊁海洋运输㊁污染物的排放及水利工程的建设等对河口及其邻近海域水动力和水环境条件㊁地貌演变等都产生了重要影响.长江口的水体环境与流域自然因素和人类活动影响密切ꎬ而在长江经济带的发展中ꎬ人类活动加剧ꎬ长江上游兴建了大量的水利水电工程ꎬ特别是三峡工程的关闸蓄水ꎬ中下游实施了大量的诸如滩涂围垦㊁河道整治㊁取排水㊁采砂㊁深水航道建设等工程ꎬ在一定程度上对长江的水文㊁泥沙特性产生了影响[19].长江三峡水利枢纽工程是中国也是世界上最大的水利枢纽工程ꎬ具有巨大的防洪㊁发电㊁航运㊁水资源利用等综合效益.但是三峡工程的建设和运营并未对长江年径流量和日均流量产生明显影响ꎬ自20世纪50年代至今ꎬ长江年径流量和日均流量均呈现波动变化ꎬ总体趋势和周期变化不明显[20]ꎬ2003年以前大通站年均流量㊁年最大流量㊁年最小流量的历史平均值分别为28635㊁60114和8428m3∕sꎬ2003年后历史平均值分别为26443㊁52191和9486m3∕sꎬ可见三峡工程运营以来ꎬ年均流量变幅不显著ꎬ年最大流量减少ꎬ年最小流量增加[21].对于最大日流量ꎬ2003年为最大日流量的显著拐点.2003年前ꎬ最大日流量呈现增加趋势ꎻ而2003年后ꎬ最大日流量值明显小于历史平均ꎬ且具有下降趋势.而日均流量在2003年前后并未发生显著差异ꎬ其趋势也不明显[21].而由于人为控制水文动力过程ꎬ三峡工程对径流年内变化趋势㊁突变特性和分配特征产生了一定的影响ꎬ洪枯季和最大日流量都有明显变化趋势ꎬ流量年内分配不均ꎬ主要集中于洪季ꎬ枯季占比较小.大通站流量丰枯率(为汛期与非汛期径流总量的比值ꎬ体现径流量年内分配)在20世纪五六十年代均较大ꎻ60年代中期到80年代末期有所减小ꎻ90年代增大ꎬ且在90年代末出现极大值ꎻ进入21世纪初以来ꎬ开始减少ꎬ并保持于一个相对较小值内[22].三峡工程的修建拦截了一部分径流ꎬ同时ꎬ水土保持及水库建成等造成的截沙效应超过水土流失造成的增沙效应ꎬ入河口输沙量降低[19]ꎬ直接影响长江口的径流来沙量ꎬ下游来沙量大幅减少ꎬ且这种减少也不是简单的数量减少[23].据统计ꎬ2003年三峡工程蓄水以来ꎬ60%~70%的上游来沙被拦截在库内ꎬ尽管坝下游河床冲刷补偿了一部分泥沙ꎬ但入河口输沙量较之前仍约下降了1∕3[19].蓄水后ꎬ长江口水文泥沙特性发生了明显变化ꎬ洪季泥沙中值粒径大于枯季ꎬ汛初流量增大阶段泥沙粗于汛末流量减小阶段ꎬ多年平均中值粒径基本不变ꎬ但泥沙有逐年变粗的趋势[19].1 1 2㊀长江口海域水质状况和沉积物质量长江口海域一直是我国近岸海域水质状况污染较严重的区域.近15年来ꎬ长江口严重污染海域主要集中在近岸ꎬ长江口北支到杭州湾南岸区域均为GB3097 1997«海水水质标准»劣Ⅳ类水质ꎬ而优良(Ⅰ类和Ⅱ类)水质面积占比不足50%(见图1).1999 2018年长江口海域主要环境要素的年际变化如图2所示.近20年来ꎬ长江口海域海水盐度整体呈下降趋势ꎬ1999 2003年波动较大ꎬ变化范围为6 88~33 16ꎬ2003年后整体趋于稳定ꎬ并呈逐年递减的趋势ꎬ2004 2018年盐度变化范围为17 00~26 79ꎬ由2004年的26 02降至2018年的18 41ꎻ海水DO年均浓度呈波动变化ꎬ整体呈上升趋势ꎬ由1999年的6 45mg∕L升至2018年的8 13mg∕Lꎬ变化范围为5 67~8 13mg∕Lꎬ其中2002年最低ꎬ2018年最高ꎻpH较稳定ꎬ变化范围为7 89~8 60ꎻ无机氮和活性磷酸盐年均浓度呈波动变化ꎬ但其年均浓度总体较高ꎬ且整体均呈上升趋势.无机氮年均浓度除20008911第5期王孝程等:长江口海域生态环境状况及保护对策㊀㊀㊀注:数据来源于2005 2018年«中国海洋环境状况公报»ꎻⅠ㊁Ⅱ㊁Ⅲ㊁Ⅳ㊁劣Ⅳ类均为GB3097 1997«海水水质标准»水质等级.图1㊀2005—2018年长江口海域水质状况趋势分布Fig.1TrendmapofwaterqualityintheYangtzeestuaryfrom2005to2018年㊁2002年和2018年外均高于0 5mg∕Lꎬ显示长江口海域长期属于GB3097 1997劣Ⅳ类水质ꎬ活性磷酸盐年均浓度2003年后长期高于0 03mg∕Lꎬ显示其多数时期属于GB3097 1997Ⅳ类水质.盐度㊁DO㊁pH㊁活性磷酸盐和无机氮等主要指标浓度在2003年前年际波动均较大ꎬ而2003年后相对较小(见图2)ꎬ这可能与人为活动的干扰有关.2003年ꎬ三峡水库开始进行一期蓄水ꎬ自蓄水后ꎬ整个长江口海域的主要指标较之前明显稳定ꎬ这可能是由于水利工程人为干预了长江径流量ꎬ从而使得长江口海域的长江径流输入㊁盐度和其他指标更加趋于稳定ꎬ长江水利工程的建设在一定程度上也对保持长江口海域水环境的稳定起到了重要作用.多年连续监测结果表明ꎬ长江口海域表层海水环境状况较差ꎬ营养盐污染严重ꎬ尤其是无机氮超标严重.长江及钱塘江径流携带东海沿岸发达的工农业生产所产生的大量污染物入海ꎬ同时每年径流也携带了大量的营养盐类ꎬ海水氮㊁磷及化学需氧量浓度超标ꎬ是造成长江口海域大面积污染的主要原因.根据«中国海洋环境状况公报»的监测结果ꎬ长江口沉积物类型为粘土质粉砂和粉砂ꎬ2005 2018年ꎬ长江口海洋沉积环境总体质量状况良好ꎬ综合质量等级年际变化基本稳定ꎬ绝大部分站位的沉积物质量最多只有一项超标要素ꎬ超标率低ꎬ而2015 20189911㊀㊀㊀环㊀境㊀科㊀学㊀研㊀究第33卷注:数据来源于1999 2004年长江口海域业务化监测结果和2005 2018年«中国海洋环境状况公报».图2㊀1999—2018年长江口海域主要环境要素的年际变化Fig.2Inter ̄annualchangeofmajorenvironmentalfactorsintheYangtzeestuaryfrom1999to2018年ꎬ长江口沉积物质量良好点位的比例已连续4年达到100%.1 2㊀长江口海域海洋生物群落和生态健康状况2011 2018年长江口海洋生物状况主要指标的年际变化如图3所示.由图3可见ꎬ浮游植物群落密度自2011年起有明显降低ꎬ2015年后有所波动ꎬ并呈逐年上升的趋势.浮游植物多样性指数呈波动状态ꎬ2011 2018年浮游植物多样性指数变化范围为0 91~2 18ꎬ整体多样性水平较低ꎬ这与逐渐增高的赤潮发生率表现出一定的相关性.综合以往的研究结果ꎬ近35年来长江口区浮游植物群落结构不断演变ꎬ种类组成趋向简单ꎬ种类个体数量分布不均匀[24]ꎬ少数优势种类(如中肋骨条藻)在环境条件合适时易大量增殖形成赤潮[25].群落结构中硅藻为浮游植物中主要类群ꎬ数量上占绝对优势ꎬ但多年来其占比呈缓慢下降趋势ꎬ甲藻种类占比缓慢增加[24].2011 2018年浮游动物密度年际波动较大ꎬ整体呈上升趋势ꎬ变化范围为288~2942ind.∕m3.浮游动物多样性指数波动较小ꎬ变化范围为1 81~2 41ꎬ多样性水平相对较高ꎬ但整体呈下降趋势.综合以往的研究结果ꎬ近35年来浮游动物群落结构趋向简单化ꎬ优势种以桡足类为主ꎬ且桡足类的组成比例有下降趋势[24]ꎬ其百分比的降低ꎬ显示浮游动物的群落结构正逐渐发生变化ꎬ这与长江口海域生境条件的日益恶化有很大关系.2011 2018年大型底栖生物密度和多样性指数年际波动较大ꎬ变化范围分别为53~175ind.∕m3㊁1 30~2 48ꎬ整体呈上升趋势.长江口及其邻近海域是我国最大的河口渔场ꎬ在我国渔业生产中居重要地位.淡水渔业资源ꎬ如凤鲚㊁刀鲚(Coiliaectenes)㊁前额间银鱼(Hemisalanxprognathus)㊁鳗鲡㊁白虾(Exopalaemon)和中华绒螯蟹ꎬ素有长江口六大渔业之称[25]ꎻ海水渔业资源ꎬ如带鱼(Trichiurusjaponicus)㊁小黄鱼(Larimichthyspolyactis)㊁大黄鱼(Larimichthyscrocea)和银鲳(Pampusargenteus)等均属该区域海洋渔业的主要捕捞对象[26].近10年来ꎬ长江口及邻近海域渔业资源因过度捕捞㊁水域生态环境和水质恶化而受到严重损害ꎬ刀鲚㊁凤鲚㊁带鱼㊁大黄鱼和小黄鱼等资源量急剧下降ꎬ低龄化和小型化明显[27]ꎬ鱼类资源量的衰退可能使甲壳类资源量相对增加[28 ̄29].由于长江口及其邻近海域受到重金属和有机物的污染ꎬ2000 2002年该海域生态环境总体质量处于重污染水平[30]ꎬ污染导致该海域渔业资源衰退[31].2005年后杭州湾可能已经成为长江口海域重金属元素重要的沉积 汇 ꎬ而长江口及其邻近海域表层沉积物中重金属0021第5期王孝程等:长江口海域生态环境状况及保护对策㊀㊀㊀注:数据来源于2011 2018年«中国海洋环境状况公报».图3㊀2011 2018年长江口海域海洋生物状况主要指标的年际变化Fig.3Inter ̄annualchangeofmajorindicatorsofmarineorganismintheYangtzeestuaryfrom2011to2018元素含量整体上均呈逐步降低的趋势ꎬ生态环境总体质量有所恢复[32].注:数据来源于2006 2018年«中国海洋环境状况公报».图4㊀2006 2018年长江口海域生态系统的健康状况Fig.4MarineecosystemhealthintheYangtzeestuaryfrom2006to20182006 2018年ꎬ长江口海域生态系统处于亚健康状态(见图4)ꎬ生态健康评价指数一直呈波动变化ꎬ范围为52 8~71 3ꎬ均低于90ꎬ其中2016年最低ꎬ2014年最高.生态健康的评价主要包含5种指标ꎬ即水环境㊁沉积环境㊁生物质量㊁栖息地和生物群落.长江口海域水环境和沉积环境基本稳定ꎬ其中沉积环境较好ꎬ而水环境一直处于较差状态ꎬ这使得栖息地环境受到威胁ꎬ由于水生生物对环境非常敏感ꎬ对水环境和栖息地的变化反应较强烈ꎬ长期处于恶劣的水质和栖息地环境下ꎬ导致生物质量整体较低ꎬ生物多样性水平较差ꎬ群落结构不稳定ꎬ生态系统健康状况处于亚健康状态.2㊀长江口海域主要的生态问题2 1㊀海水污染严重ꎬ水环境质量较差长江㊁钱塘江等江河的径流每年携带了大量的营养盐类进入长江口海域ꎬ该海域水体污染物浓度较高ꎬ氮㊁磷及化学需氧量浓度均超过GB3097 1997Ⅳ类水质标准限值[33 ̄35].无机氮年均浓度显示长江口海域长期属于劣Ⅳ类水质ꎬ而活性磷酸盐年均浓度显示其多数时期属于Ⅳ类水质.目前ꎬ长江口海域是我国海水水质极差的海域之一.除多年水质极差外ꎬ«中国海洋环境状况公报»显示ꎬ长江口海域生物体内的油类㊁总汞㊁砷㊁铅和滴滴涕等指标浓度也普遍超标.环境质量差是致使长江口海域多年来处于亚健康的主要原因之一.2 2㊀海洋工程和人类活动干扰强烈ꎬ生境破坏严重上海长江隧桥工程㊁杭州湾大桥工程㊁长兴岛造船基地工程㊁长兴 崇明 启东桥隧工程项目㊁长江口深水航道三期疏浚工程和洋山深水港工程等工程1021㊀㊀㊀环㊀境㊀科㊀学㊀研㊀究第33卷的施工和完成ꎬ导致长江口海区海洋生物栖息地严重破碎化.另外ꎬ滩涂养殖的过度发展ꎬ也使余姚和慈溪沿岸的滩涂生物简单化ꎬ基本形成了由单一养殖物种组成的滩涂湿地生物结构ꎬ大大降低了滩涂湿地的物种多样性.同时海洋工程占用了海洋生物的生存空间及洄游路线ꎬ使多个自然洄游通道遭到不同程度的破坏.生境的破碎化和洄游通道的阻断ꎬ加之大型船只频繁穿梭等干扰(包括噪声污染等)ꎬ不仅影响一般过河口性和定居性生物的产卵㊁育幼㊁生长和生存ꎬ而且经常造成许多珍稀动物的非正常死亡.2 3㊀低氧区长期存在ꎬ成为生态安全的重要潜在威胁长江口海域水体中DO浓度虽然近20年有所升高ꎬ但是仍监测到低氧区的存在[36 ̄38].2002年ꎬ科学家们在长江口及其邻近海域底层发现存在面积约为13700km2㊁DO浓度小于2mg∕L的低DO区域ꎬ最低处仅为1mg∕L[39]ꎬ而2007年在长江口外海区发现了一个更大的近20000km2的低氧区域[40].研究[41]发现ꎬ20世纪90年代后ꎬ低氧现象的发生概率已逐渐升至90%.低氧区的存在ꎬ可导致大量海洋生物窒息死亡ꎬ而低氧区消除和恢复则需要漫长的时间ꎬ但迄今未见有消除和恢复迹象.随着长江口海域水体中DO浓度的变化ꎬ低氧区的范围和程度可能进一步扩大和加剧ꎬ成为长江口海域生态系统的重要潜在威胁ꎬ最终成为长江口生态系统中的生物死亡区或无生物区.2 4㊀生物群落状况较差ꎬ生态系统健康总体欠佳由于长江口海域生境条件的日益恶化ꎬ浮游植物群落种类组成发生明显变化ꎬ浮游植物中硅藻的占比有所下降ꎬ甲藻有所上升[24]ꎬ赤潮种类数量异常增殖引发赤潮ꎻ浮游动物种类明显减少ꎬ密度普遍偏低ꎬ原来的优势种类桡足类的种类和数量均呈下降趋势ꎬ结构趋于简单化[42 ̄44]ꎬ2004年桡足类占浮游动物种类数的50%ꎬ2005年㊁2006年分别降至46%和42%ꎬ2007年降至30%以下ꎬ2008年因种类数㊁生物量和密度均呈较大幅度升高ꎬ桡足类的占比也有所反弹ꎬ2009年之后一直在较低水平波动[24].渔业资源衰退明显ꎬ长江口及杭州湾传统渔场接近消失边缘[45].长江口海域生态系统健康状况欠佳ꎬ其主要原因是:①捕捞压力过大ꎬ近10年来优质渔业资源严重衰退ꎻ长三角海域近岸鳗鱼苗网密布ꎬ对近岸鱼类产卵场㊁索饵场及洄游通道影响极大.②近年来ꎬ三峡水利工程建设和上游工农业用水量增大ꎬ虽对年均径流量无明显影响ꎬ但人为的干预对径流年内变化趋势㊁突变特性和分配特征产生了一定的影响ꎬ使得水流对于岸滩的冲击作用发生改变ꎬ严重地改变了河口生境ꎬ导致产卵场和育幼场功能逐渐丧失㊁鱼类等生物生殖及生长洄游通道受阻ꎬ河口生态系统的生态服务功能丧失严重.③海洋生物饵料来源不稳定ꎬ磷酸盐和无机氮污染严重ꎬ饵料生物的种类组成和优势种类年际变化较大.2 5㊀外来生物入侵ꎬ赤潮频发随着上海国际航运中心的确立和运营ꎬ洋山港和北仑港大型港口经由远洋船只压舱水携带等途径带来的外来海洋生物日益增多ꎬ特别是外来浮游植物入侵种类的数量越来越多ꎬ土著硅藻种类占比日趋减少ꎬ甲藻类中的有毒赤潮生物的种类和数量不断增多ꎬ时常引发赤潮[46 ̄47]ꎬ其主要原因是:①由于长江口生态系统日趋恶化和脆弱化ꎬ为外来种提供了生存㊁增殖和引发赤潮的条件ꎻ②环境条件的变化致使土著种类不再具有适宜的生境条件ꎬ多数土著种类的种群数量减少甚至消失ꎬ但对于少数土著种类ꎬ如广生性和耐污性较强的中肋骨条藻ꎬ在环境条件合适时也会大量增殖ꎬ并形成赤潮.总体而言ꎬ浮游植物种类多样性明显下降ꎬ群落结构趋向简单化且不稳定.3㊀长江口海域保护修复及管理对策3 1㊀加强顶层设计ꎬ推进落实陆海统筹通过对长江口海域生态环境质量现状的分析和科学评价ꎬ认为在长江口海域生态环境管理中ꎬ应高度重视陆海统筹与区域协调机制的建设. 湾区经济 已经成为带动全球经济发展的增长极ꎬ推动湾区发展已然成为世界各国发展开发型经济㊁确立战略优势的重要经验.长江口海域作为我国极其重要的流域㊁海域交汇区ꎬ其良好的生态环境质量不仅关乎海洋生态环境ꎬ更关乎整个区域的经济社会发展.对长江口海域的生态环境治理必然要加强落实陆海统筹的顶层设计.a)规划引领.规划是进行区域调控和管理的重要工具ꎬ具有前瞻性㊁战略性㊁地域性和约束力.落实«中共中央国务院关于加快推进生态文明建设的意见»和«水污染防治行动计划»部署ꎬ按照«长江经济带生态环境保护规划»的要求ꎬ依据有关海洋环境保护法律法规㊁生态市建设规划和海洋经济发展规划等ꎬ编制海洋生态环境保护与建设相关专项规划ꎬ通过规划引领区域环境合作行动.b)建立区域协调机制.2018年的机构改革ꎬ在生态环境保护领域打通了陆地和海洋ꎬ破除了陆域㊁海域环境保护与管理之间的体制壁垒ꎬ为生态环境保2021第5期王孝程等:长江口海域生态环境状况及保护对策㊀㊀㊀护管理的陆海统筹奠定了良好基础.应充分发挥我国生态环境领域改革的制度优势ꎬ整合和发挥生态系统整体性的经济规模效应和污染治理的规模效应ꎬ建立区域协调机制ꎬ全流域 一盘棋 考虑ꎬ加快促进河(湖)长制㊁湾长制等流域㊁海域环境治理协调机制在治理对象㊁治理范围㊁技术标准等方面的有效衔接ꎬ倒逼和统筹河流㊁海域的污染控制目标和考核指标ꎬ突破现有陆海污染物管控不衔接问题ꎬ进一步制定落实流域㊁海域生态环境管理的政策措施体系ꎬ实施河口海湾区域生态环境治理的合理规划㊁共治共管ꎬ强化不同环境政策之间的协同和协调ꎬ为海洋环境保护奠定区域环境合作的政策基础.c)强化科技创新有效供给.充分发挥国家长江生态环境保护修复联合研究中心的平台枢纽作用ꎬ切实强化长江流域科技创新的有效性供给ꎬ推动国家水体污染控制与治理科技重大专项等重大专项成果转化ꎬ重点强化污染物来源解析与综合诊断技术ꎬ地表 地下㊁河 海多过程协同的流域水环境调控技术研究ꎻ加强农业农村污染防治㊁生态保护修复适用技术推荐ꎻ以污染物及其生态效应管控为目标ꎬ开展陆域㊁水体统筹兼顾的治理优先区识别ꎬ引领投资与保护方向.3 2㊀科学规划临港产业空间布局ꎬ完善陆海统筹的治污体系临港产业布局事关海洋经济的长远发展ꎬ事关人民群众福祉.合理的临港产业布局有利于充分利用各种要素资源ꎬ发挥比较优势ꎬ有利于防止生态环境污染ꎬ维持生态平衡ꎬ提高土地集约利用ꎬ是区域经济持续㊁健康发展的必要条件之一ꎬ对区域经济发展具有非常显著的影响.应科学规划临港产业空间布局ꎬ完善陆海统筹的治污体系.a)优化临港产业空间布局规划.按照生态环保优先㊁人与自然和谐㊁陆地与海洋统筹㊁海洋生态环境保护与临海产业发展统筹安排的原则ꎬ做好临港产业布局顶层设计ꎬ统筹产业发展规划ꎬ从源头控制临港产业海洋环境污染.针对临港产业布局现状ꎬ客观分析存在的问题ꎬ进一步调整优化临港产业布局ꎬ以实现海洋经济建设与海洋生态环境保护更为协调发展.b)加强涉海产业的污染管理.将长江口流域的污染治理与海洋环境保护结合起来ꎬ建立陆海统筹的生态修复与污染防治联动机制ꎬ分清轻重缓急ꎬ分级分区实现精准施策.依据长江口流域㊁海域生态环境污染防治的特征ꎬ系统全面推进水污染综合治理ꎬ加大在治水体制和生态补偿机制等方面的技术与政策支持ꎬ加快流域㊁海域水环境质量的全面改善.禁止在沿岸及岛屿新建㊁扩建污染海洋生态环境的项目ꎬ对现有的企业事业单位超过标准排放污染物的ꎬ要依法限期治理ꎬ对污染严重㊁难于治理或治理后仍达不到要求的涉海产业ꎬ要按照管理权限坚决依法予以关停.3 3㊀加强污染物入海排放管控ꎬ提升海洋环境保护意识通过实施环评㊁总量控制等制度ꎬ优化排污口布局ꎬ严格管理围填海活动ꎬ加强污染物入海排放管控ꎬ逐步减少入海污染物总量.具体措施包括:①严格海洋环评制度.发展海洋经济必须以环境容量为前提ꎬ要加强涉海工程的建设监督管理ꎬ严格执行海洋经济发展规划与项目的环境影响评价和环保设施 三同时 制度ꎬ排放非达标项目坚决一票否决ꎬ确保海洋经济可持续发展.②严格管理围填海活动.严格围填海项目审查ꎬ严格执行围填海禁填限填要求ꎬ从严限制单纯获取土地性质的围填海项目ꎬ制定并严格执行围填海规划ꎬ除政府组织的海域海岸带整治少量填海外ꎬ在港口航道附近和港湾区域要禁止围填海.③严格涉海产业准入.制订严格的涉海产业准入标准ꎬ项目选址要进行科学论证ꎬ特别是要强化对布局密集㊁规模庞大的化工㊁钢铁㊁火电㊁炼油项目环评论证ꎬ严格落实涉海产业准入和环保要求ꎬ择优发展临港工业ꎬ禁止高污染㊁高排放企业在临港落户.④对主要工业污水实行深度处理和废水回用ꎬ提高污水处理脱氮㊁脱磷效率ꎬ实现工业污水达标排放和有毒有害污染物 零排海 .加强城市污水处理设施㊁沿岸污水管网系统和中水回用系统建设ꎬ提升生活污水处理能力ꎬ实现城市污水100%处理ꎬ再生水100%回用.重视农业面源污染的治理ꎬ发展高效农业和先进的施肥方式ꎬ降低化肥㊁农药使用量.⑤以 三磷 综合整治㊁城镇污水收集与治理能力提升为抓手ꎬ继续强化磷污染工业和生活点源污染全过程防控.与此同时ꎬ大力推进重点区域面源污染综合管控.结合面源普查㊁污染通量测算等结果ꎬ宜将湖北省㊁湖南省㊁江苏省㊁安徽省㊁江西省5个省份作为重点区域ꎬ将汛期水质恶化河流∕湖泊作为重点对象ꎬ切实强化污染治理.⑥合理调整养殖布局和结构ꎬ控制养殖自身污染.推进生态渔业建设ꎬ建立和优化鱼㊁贝㊁藻间养和轮养复合生态养殖模式ꎬ重点鼓励发展浅海藻类养殖ꎬ根据养殖环境容量ꎬ调整和优化海水网箱养殖布局ꎬ开展养殖网箱标准化改造建设ꎬ推广应用配合饲料.3 4㊀保障海洋生态建设资金ꎬ强化海洋生态保护与建设3021。
长江口中华鲟自然保护区底栖动物

长江口中华鲟自然保护区底栖动物张凤英;庄平;徐兆礼;王云龙;朱江兴【期刊名称】《生态学杂志》【年(卷),期】2007(26)8【摘要】根据2004年5、8、11月和2005年2月在长江口中华鲟自然保护区附近水域(31°19.58′—31°38′N;121°32.08′—122°11.65′E)4个航次的海洋综合调查资料,分析了该水域底栖动物种类组成、数量变动和优势种时空分布以及与中华鲟幼鱼食性的关系。
结果表明:调查区共有底栖动物48种,种类组成和优势种均有明显的季节更替,四季皆为优势种的仅纵肋织纹螺(Nassariusvariciferus)和狭颚绒螯蟹(Eriocheirleptognathus)2种;夏季总生物量在四季中最高,春季次之,冬季最低;夏季总栖息密度大于其它3个季节,冬季次之,春季和秋季基本一致;夏季是底栖动物总生物量和总栖息密度较高的时期,也是中华鲟幼鱼在长江口停留觅食时期。
与2002年的研究结果进行比较表明,虽然底栖动物优势种组成在深水航道工程后基本稳定,但生物量和栖息密度均呈下降趋势,这直接威胁到保护区内中华鲟幼鱼的饵料基础状况。
【总页数】6页(P1244-1249)【关键词】长江口;中华鲟幼鱼;底栖动物【作者】张凤英;庄平;徐兆礼;王云龙;朱江兴【作者单位】中国水产科学研究院东海水产研究所农业部海洋与河口渔业重点开放实验室【正文语种】中文【中图分类】S963【相关文献】1.长江口中华鲟自然保护区管理办法出台:中华鲟享受特殊"待遇" [J],2.上海市长江口中华鲟自然保护区顺利实施中华鲟放流 [J],3.拯救国宝中华鲟促进长江大保护上海长江口中华鲟自然保护区专注中华鲟保护13年 [J],4.70尾大中型中华鲟放流长江口长江口中华鲟自然保护区基地动工建设 [J], 吴铠5.上海中华鲟自然保护区在长江口再次放流1500多尾中华鲟 [J],因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第35卷第15期2015年8月生态学报ACTA ECOLOGICA SINICA Vol.35,No.15Aug.,2015基金项目:国家重点基础研究发展计划(973)项目(2011CB403604);中国科学院知识创新工程重要方向项目(KSCX2⁃EW⁃Z⁃5);国家自然科学基金项目(41306153);中国科学院海洋研究所 一三五”专项资助项目(2012IO060104)联合资助收稿日期:2014⁃01⁃02; 修订日期:2014⁃12⁃21*通讯作者Corresponding author.E⁃mail:kxu@ DOI :10.5846/stxb201401020008周百灵,孟昭翠,赵峰,徐奎栋.长江口外海域微型和小型底栖生物群落结构和时空变化.生态学报,2015,35(15):5050⁃5063.Zhou B L,Meng Z C,Zhao F,Xu K munity structure and spatio⁃temporal variation of microbenthos and meiobenthos in sediments off the Yangtze Estuary.Acta Ecologica Sinica,2015,35(15):5050⁃5063.长江口外海域微型和小型底栖生物群落结构和时空变化周百灵1,2,孟昭翠1,赵 峰1,徐奎栋1,*1中国科学院海洋研究所,海洋生物分类与系统演化实验室,青岛 2660712中国科学院大学,北京 100049摘要:利用DAPI 荧光计数法和Ludox⁃QPS 方法并结合环境因子的分析,研究了2010年和2011年夏季(7月)及秋季(11月)长江口至济州岛沿线的CJ 断面沉积物中的微型及小型底栖生物的组成㊁丰度和生物量及分布特点㊂结果表明,表层5cm 沉积物中,细菌(108个/cm 3)㊁蓝细菌(106个/cm 3)㊁自养(106个/cm 3)和异养微型鞭毛虫(105 106个/cm 3)的丰度较其他类群高3个数量级以上;细菌(19 24μg C /cm 3)㊁自养(13 31μg C /cm 3)和异养(5 44μg C /cm 3)微型鞭毛虫的生物量明显较蓝细菌(1 2μg C /cm 3)㊁小型底栖生物(0.9 1μg C /cm 3)㊁纤毛虫(0.04 0.2μg C /cm 3)㊁异养小鞭毛虫(0.02 0.08μg C /cm 3)及硅藻(0.001 0.008μg C /cm 3)高㊂除细菌和蓝细菌外,2011年微型底栖生物和小型底栖生物各类群的丰度和生物量大多高于2010年,且大多为秋季高于夏季㊂发现的166种纤毛虫中,肉食性纤毛虫所占现存量的比例最大,菌食性次之,藻食性和杂食性最小;肉食性纤毛虫在秋季所占比例高于夏季,而菌食性则相反㊂统计分析表明,微型底栖生物丰度和生物量年度间具有极显著差异,基于物种⁃丰度的纤毛虫群落结构年度间呈极显著差异,而小型底栖生物则未见显著差异㊂微型和小型底栖生物各类群夏季和秋季的分布受多因子而非单一环境因子的影响,长江冲淡水对该断面的微型和小型底栖生物的影响不显著㊂对微型底栖生物各类群及水体中相应类群的比较分析表明,该海域的微型底栖生物具有明显的数量和功能重要性㊂关键词:微型底栖生物;小型底栖生物;丰度;生物量;分布;东海Community structure and spatio⁃temporal variation of microbenthos and meiobenthos in sediments off the Yangtze EstuaryZHOU Bailing 1,2,MENG Zhaocui 1,ZHAO Feng 1,XU Kuidong 1,*1Department of Marine Organism Taxonomy and Phylogeny ,Institute of Oceanology ,Chinese Academy of Sciences ,Qingdao 266071,China 2University of Chinese Academy of Sciences ,Beijing 100049,China Abstract :Using epifluorescence microscopy and the Ludox⁃QPS method,we investigated the community composition,abundance,and biomass as well as distribution of microbenthos and meiobenthos in combination with the analysis of benthic environmental variables at four stations off the Yangtze Estuary in the summer (July)and fall (November)of 2010and2011,respectively.The abundances of bacteria (108cells /cm 3),cyanobacteria (106cells /cm 3),and phototrophic (PNF,106cells /cm 3)and heterotrophic nanoflagellates (HNF,105 106cells /cm 3)were much higher than those of other components in the upper 5cm of sediments.The biomass partitioning indicates the primary importance of benthic bacteria (19 24μg C /cm 3),followed by PNF (13 31μg C /cm 3),HNF (5 44μg C /cm 3),cyanobacteria (1 2μg C /cm 3),meiobenthos (0.9 1μg C /cm 3),ciliates (0.04 0.2μg C /cm 3),heterotrophic microflagellates (HMF,0.020.08μg C /cm 3),and diatoms (0.001 0.008μg C /cm 3).The standing crops of microbenthic and meiobenthic assemblages,except of bacteria and cyanobacteria,were generally higher in 2011than in 2010and usually higher in the fall than in the summer.About 84%of diatoms,82%of ciliates,65%of HMF,and 77%of meiobenthos were distributed in the upper 2cm of the sediments,whereas no distinct vertical distribution was observed for bacteria,cyanobacteria,PNF,and HNF.The vertical distribution had little variation between the years and seasons.The estimated productivity of diatoms ranged from 0.05to 0.4mg C m-2d -1and contributed only 0.01% 0.1%of the primary productivity.Thus,most of the primary productivity was contributed by the pico⁃and nano⁃sized benthos.The estimated productivity of bacteria was 80 101mg C m-2d -1,about 17% 21%of the primary productivity.The biomass of bacteria was nine to 18times higher than that of the meiobenthos,whereas the productivity was 30 61times higher than that of the meiobenthos (2 3mg C m -2d -1).The biomass of ciliates was much lower than that of the meiobenthos,while the ciliate productivity (3 23mg C m -2d -1)was one to 14times higher than that of the meiobenthos.The relative metabolic rates indicate that benthic heterotrophic nanoflagellates constitute the most important group in sediment respiration,accounting for 92.9% 99.1%of the estimated combined metabolic rate of the microbenthic and meiobenthic consumers.Among the 166identified ciliate species,carnivorous ciliates constituted the primary feeding type in standing crops,followed by bacterivores,algivores,and omnivores.Carnivorous ciliates contributed a relatively higher proportion of standing crops in the fall than in the summer,whereas the contribution of bacterivorous ciliates was inverse.The abundance and biomass of the microbenthos as well as the ciliate community structure based on species⁃abundance varied significantly between the years,while no significant difference was observed for meiobenthos.Statistical analyses show that the distribution of microbenthos and meiobenthos in the summer and fall was affected by multiple factors rather than a single environmental factor,and diluted water of the Yangtze River did not show an obvious impact on the microbenthos and meiobenthos.BIOENV analysis indicates that in the summer,the microbenthic abundance had the highest correlation with the chlorophyll a concentration and ciliate species⁃abundance had the highest correlation with the combination of the bottom water temperature,chlorophyll a concentration,and organic matter content,while no correlation was found for meiobenthos.In the fall,the meiobenthic abundance had the highest correlation with the median grain size of the sediments,while no correlation was found for the microbenthos and parative analyses between the microbenthic assemblages and corresponding ones in the upper pelagic waters suggest that the microbenthos had numerical and functional importance in the study area.Key Words :microbenthos;meiobenthos;abundance;biomass;distribution;East China Sea海洋底栖生物是通常指生活史的全部或大部分时间栖息于沉积物内㊁底表或水域底部的生物总称,按粒径大小可分为微型㊁小型和大型底栖生物㊂微型底栖生物(microbenthos)主要包括原核微生物㊁真核微藻和原生动物等[1];小型底栖生物(meiobenthos)是指能通过500μm 孔径网筛,而被42μm 或31μm 孔径网筛所阻留的后生动物,主要包括线虫㊁桡足类㊁多毛类㊁动吻类等类群[2]㊂微型和小型底栖生物作为底栖微/小食物网重要组成部分,通过复杂的营养关系形成一个有机整体,在底栖生态系统中起着重要作用[3]㊂原核微生物及微藻在底栖生态系统的初级生产㊁有机质利用及营养物质循环中起着基础作用,原生动物和小型动物是底栖微食物网中由初级生产力向高营养级转换的枢纽㊂微型和小型底栖生物的群落结构㊁现存量及分布格局的变动会对底栖微食物网及近岸浅海生态系统产生影响㊂微型底栖生物由于个体微小且定量困难,需从泥沙中有效分离和提取出来才能进行定性和定量分析,国际上对海洋微型底栖生物各类群的研究少,且主要针对潮间带生境[4⁃6],我国对海洋微型底栖生物的现存量及生态特点仅有零星报道[7⁃8]㊂国际上对小型底栖生物的研究较为深入[9],国内相关研究主要集中在黄渤海,对长江口海域研究相对较少[10⁃11]㊂长江是我国流入东海的最大河流,长江口及其邻近海域在长江巨量径流㊁复杂流系结构和特珠地形的共1505 15期 周百灵 等:长江口外海域微型和小型底栖生物群落结构和时空变化 同影响下形成独特的生态特征㊂有研究表明,夏季长江冲淡水123°E 附近存在浮游植物生物量和初级生产力高值区[12],浮游纤毛虫丰度较外围海域高1倍[13];小型底栖生物在冲淡水区域丰度也较外围海域高[11]㊂夏季台湾暖流水的入侵可致使浙江三门湾大型底栖生物群落种类数减少[14]㊂目前,尚无冲淡水及台湾暖流对微型底栖生物群落结构影响的报道㊂此外,2011年夏季黄㊁东海出现水母旺发,且可对纤毛虫和鞭毛虫等微小型浮游生物产生明显影响[15],但对底栖生态系统的影响尚不可知㊂本研究旨在了解长江口外海区微型和小型底栖生物的类群构成㊁丰度㊁生物量及分布的时空变化,分析其时空变化与环境因子的关系,并探讨环境因子对其分布的影响,为长江口底栖生态系统尤其是极为欠缺的微型底栖生物生态学研究提供必要资料㊂1 材料与方法1.1 样品采集本研究搭载中国科学院海洋研究所2010年㊁2011年7月(夏季)和11月(秋季)开放共享航次,对黄㊁东海的交界处 长江口至韩国济州岛沿线的CJ 断面进行了底栖生物采样调查,在CJ2,CJ5,CJ6和CJ7四个站位成功采集到微型和小型底栖生物样品及用于环境因子分析的样品㊂其中,2010年秋季CJ6站位未成功采到相关样品,CJ5站位仅采到纤毛虫㊁小鞭毛虫㊁硅藻及小型底栖生物样品(图1)㊂图1 黄、东海交汇区域典型断面采样站位图 Fig.1 Sampling stations along a representative transect in the boundary area between the Yellow Sea and the East China Sea CJ2,CJ5,CJ6,CJ7:采样点利用0.1m 2改进型Gray⁃Ohara 箱式采泥器在每一站位采集3箱未受扰动的沉积物样品,使用内径为1.6cm 的注射器改造的采样管随机采集5cm 长芯样4个(前两箱各采1管,第三箱采集2管),每2个芯样按0 2cm㊁2 5cm 分层合并分别移入50mL 离心管中,加入经滤膜(孔径0.22μm)过滤的海水配制的2%冷戊二醛至40mL 进行固定㊂从离心管中各取20mL 按照分层合并共计40mL,4℃避光保存用于细菌㊁蓝细菌及微型鞭毛虫分析㊂另外,采用内径为2.3cm 的注射器改造的采样管采集8cm 长芯样8个,按0 2cm㊁25cm㊁5 8cm 进行分层,其中4个芯样的各分层分别瓶装,用终浓度为2%冷戊二醛溶液固定,4℃避光保存用于纤毛虫㊁硅藻㊁小型鞭毛虫和小型底栖生物分析;另外4个芯样的相应分层合并后,分别装入封口袋,-20℃冷冻保存,用于沉积物粒度㊁叶绿素a㊁脱镁叶绿素a㊁有机质及总氮测定㊂1.2 样品及环境因子分析水深及底层水温度㊁盐度数据由随船温盐深测定仪(CTD)现场测定㊂沉积物粒度分析采用Cilas (940L)型激光粒度仪测定㊂叶绿素a 和脱镁叶绿素a 含量利用Turner II 荧光光度计测定㊂有机碳和总氮含量利用元素分析仪Vario Macro Cube (Elementar,Germany)测定,有机质含量为有机碳含量乘以系数1.724㊂沉积物中的细菌㊁蓝细菌和微型鞭毛虫经过样品分散㊁超声波处理㊁稀释后,经DAPI 荧光染色后镜检计数[7],其中微型鞭毛虫根据其色素颗粒的有无对自养(PNF)与异养微型鞭毛虫(HNF)分别计数,并按2 5μm㊁5 10μm㊁10 20μm 3个粒级进行计数㊂沉积物中底栖硅藻㊁纤毛虫和异养小鞭毛虫(HMF)及小型底栖生物利用Ludox⁃QPS 染色方法进行分析[16],主要步骤包括:密度梯度离心提取,定量蛋白银染色(QPS),制片,镜检,对微型和小型底栖生物进行鉴定和计数㊂微型和小型底栖生物的生物量通过生物体积换算法和相应转换系数进行计算[1,17⁃18]㊂对沉积物中初级生产力的估算按照Davis [19]等:P =9.57+0.35Chl a,P 为初级生产力总量(mg C m -2h -1),Chl a 为沉积物中叶绿素a 含量(mg /m 2)㊂底栖硅藻的生产力通过其增长率(μ)和生物量(B )来估2505 生 态 学 报 35卷 算:P =μ×B ,式中增长率的估算依据Montagnes 等[20]:μ=-2.40×0.85T -0.0056log V ×T +1.10,式中μ为增长率(/d),V 为硅藻体积(m m 3),T 为温度(°C)㊂底栖细菌生产力的估算依据Cole 等[21]:log P =2.054+1.08Log B ,式中P 为生产力(μg C m -2d -1),B 为细菌生物量(mg C /m 2)㊂底栖纤毛虫的生产力通过其增长率和生物量来估算,增长率的估算依据Müller [22]:ln μm =1.52×ln T -0.27×ln V -1.44,式中μm 为增长率(/d),T 为温度(°C),V 为纤毛虫体积(m m 3)㊂小型底栖生物的生产力以P =9B 估算[23]㊂依据Fenchel 中R =a W -0.25对纤毛虫㊁异养鞭毛虫和小型底栖生物的相对新陈代谢率进行估算,式中纤毛虫和异养鞭毛虫的a 为10-1.94,小型底栖动物的a 为10-1.64,W 为估算的平均个体重(约为总生物量/总丰度)[24]㊂表层0 2cm 沉积物中纤毛虫对细菌的摄食率按37 525个细菌纤毛虫-1h -1[4,25⁃26]进行估算,纤毛虫对硅藻的摄食率估算按17个硅藻纤毛虫-1㊃h -1[4]㊂1.3 数据统计分析采用Surfer 8.0绘制站位图,采用Excel 绘制柱状图㊂采用SPSS 15.0统计软件对微型和小型底栖生物各类群丰度与环境因子进行相关分析㊂利用PRIMER 6(Plymouth Routines in Multivariate Ecological Research)软件包中的CLUSTER 分析各研究站位的环境因子㊁微型和小型底栖生物丰度和生物量的等级聚类;利用ANOSIM 分析环境因子㊁各类群丰度和生物量年度㊁季节和站位间差异;利用BIOENV 分析找出与生物群落结构相关性最高的环境因子组合㊂为使数据正态分布,原始数据经过log (x +1)转化处理㊂2 结果2.1 采样站位的环境因子所研究4个站位的水深从CJ2到CJ7逐渐加深,依次为28,43,45,58m (图2)㊂2010年和2011年夏季4个站位底层水温度平均值差异不大,依次为15.4±3.2(平均值±标准差)℃㊁(15.6±3.5)℃;两年秋季底层水温度均高于夏季,依次为(17.9±0.4)℃㊁(19.7±0.7)℃㊂从CJ2到CJ7夏季各站位底层水温度逐渐降低,而两年秋季各站位温度先降低后升高,但总体上各站差异不大(图2)㊂2010年和2011年夏季各站底层水盐度分别为31.8±0.6㊁32.2±0.6,秋季底层水盐度分别为32.7±0.7㊁32.4±0.8,年度和季节之间的变化均不大,从CJ2到CJ7站底层水盐度先降低后升高,但CJ5站2010年秋季同比下一年度明显较高(图2)㊂研究海域4个站位的沉积物类型主要为粉砂,中值粒径为5 185m m㊂年度和季节间的变化均不大,从CJ2到CJ7站中值粒径先增大后减小,从粉砂变为粉砂质砂再变为粘土质粉砂,其中2010年夏季CJ7站中值粒径高达185m m,远大于其他站位(图2)㊂表层2cm 沉积物中,2010年和2011年夏季各站叶绿素a 含量依次为(0.67±0.37)m g /g 沉积物干重㊁(0.45±0.03)m g /g,脱镁叶绿素a 含量依次为(3.64±1.19)m g /g㊁(5.46±1.03)m g /g;,2010年和2011年秋季叶绿素a 含量依次为(0.52±0.05)m g /g㊁(0.46±0.18)m g /g,脱镁叶绿素a 含量依次为(5.49±0.65)m g /g㊁(6.26±3.07)m g /g㊂叶绿素a 含量夏季略高于秋季,2010年略高于2011年,而脱镁叶绿素a 含量的变化却相反㊂从CJ2到CJ7叶绿素a 含量逐渐降低,而脱镁叶绿素a 含量先增高后降低(图2)㊂2010年和2011年各站夏季沉积物有机质含量分别为(0.56±0.23)%㊁(0.59±0.19)%,秋季含量分别为(0.56±0.18)%㊁(0.63±0.43)%㊂年度和季节间变化均不大,从CJ2到CJ7有机质含量先升高后降低,其中2011年秋季CJ6站有机质含量高于其他站位(图2)㊂2010年夏季未获得沉积物总氮数据,2011年夏季各站沉积物总氮含量为(0.04±0.02)%,2010年和2011年秋季含量分别为(0.05±0.03)%㊁(0.09±0.03)%㊂从CJ2到CJ7总氮含量先升高后降低,2011年秋季CJ6站总氮含量高于其他站位(图2)㊂综合环境因子各变量,经CLUSTER 聚类分析显示,基本上相同站位聚集在一起㊂单因子相似性检验3505 15期 周百灵 等:长江口外海域微型和小型底栖生物群落结构和时空变化 图2 长江口外海域各研究站位(CJ2⁃7)的环境因子Fig.2 Benthic environmental variables at the stations CJ2⁃7off the Yangtze Estuary(One⁃Way ANOSIM)结果显示,站位差异>年度差异>季节差异,不同站位间有极显著差异(Global R =0.456,P =0.002),年度间和季节间均无显著差异㊂2.2 微型底栖生物丰度和生物量的时空变化2010年和2011年夏季长江口外海区表层0 5cm 沉积物中,各站细菌丰度分别为(5.47±0.50)×108个/cm 3和(4.62±1.02)×108个/cm 3,生物量为(24.05±2.19)μg C /cm 3和(20.33±4.50)μg C /cm 3,两年秋季丰度为(4.97±0.72)×108个/cm 3和(4.42±1.09)×108个/cm 3,生物量为(21.88±3.17)μg C /cm 3和(19.46±4.78)μg C /cm 3㊂从CJ2站到CJ7站细菌丰度和生物量呈现先降低后增高的趋势,最低值多出现在CJ5站,最高值多出现CJ6站(图3,图4)㊂4505 生 态 学 报 35卷 图3 长江口外海域各研究站位(CJ2⁃7)微型和小型底栖生物各类群的丰度Fig.3 The abundances of microbenthos and meiobenthos at the stations CJ2⁃7off the Yangtze Estuary 在表层0 5cm 沉积物中,2010年和2011年夏季各站蓝细菌丰度为(1.50±0.34)×106个/cm 3㊁(1.36±0.31)×106个/cm 3,生物量为(1.95±0.45)μg C /cm 3㊁(1.77±0.40)μg C /cm 3,两年秋季丰度为(1.15±0.27)×106个/cm 3㊁(1.09±0.25)×106个/cm 3,生物量为(1.50±0.35)μg C /cm 3㊁(1.42±0.33)μg C /cm 3㊂从CJ2站到CJ7站蓝细菌丰度和生物量呈现逐渐降低趋势,最低值多出现在CJ7站(图3,图4)㊂自养微型鞭毛虫(PNF)在0 5cm 沉积物中的丰度,两年夏季依次为(2.84±0.10)×106个/cm 3㊁(3.15±0.29)×106个/cm 3,生物量依次为(16.52±3.22)μg C /cm 3㊁(17.52±3.63)μg C /cm 3,两年秋季各站丰度依次为(2.09±0.21)×106个/cm 3㊁(3.63±0.41)×106个/cm 3,生物量依次为(12.51±2.78)μg C /cm 3㊁5505 15期 周百灵 等:长江口外海域微型和小型底栖生物群落结构和时空变化 (30.92±7.72)μg C /cm 3㊂从CJ2站到CJ7站PNF 丰度呈逐渐增高的趋势,最低值多出现在CJ2站(图3,图4)㊂按鞭毛虫大小看,2 5μm 鞭毛虫的丰度最高(可达106个/cm 3),5 10μm 鞭毛虫丰度次之(105个/cm 3),10 20μm 鞭毛虫丰度最小(104个/cm 3)㊂图4 长江口外海域各研究站位(CJ2⁃7)微型和小型底栖生物各类群的生物量Fig.4 The biomasses of microbenthos and meiobenthos at the stations CJ2⁃7off the Yangtze Estuary异养微型鞭毛虫(HNF)在0 5cm 沉积物中的丰度,各站两年夏季依次为(3.12±2.39)×105个/cm 3㊁(1.95±0.20)×106个/cm 3,生物量依次为(5.23±3.67)μg C /cm 3㊁(30.60±5.09)μg C /cm 3,两年秋季丰度依次为(5.48±0.20)×105个/cm 3㊁(4.24±1.43)×106个/cm 3,生物量依次为(7.19±0.82)μg C /cm 3㊁(43.72±13.87)μg C /cm 3㊂从CJ2站到CJ7站HNF 丰度呈现先增高后降低的趋势,最高值多出现在CJ5站(图3,6505 生 态 学 报 35卷 图4)㊂各粒级异养微型鞭毛虫丰度同样也是2 5μm 粒级最高(105 106个/cm 3),其次是5 10μm 粒级(多为104 105个/cm 3),10 20μm 粒级丰度最小(多为104个/cm 3)㊂在表层0 8cm 沉积物中,2010年和2011年夏季各站异养小鞭毛虫(HMF)的丰度分别为(5±5)个/cm 3㊁(10±4)个/cm 3,生物量为(0.0041±0.0036)μg C /cm 3㊁(0.0073±0.0029)μg C /cm 3㊂两年秋季丰度分别为(10±4)个/cm 3㊁(32±12)个/cm 3,生物量为(0.0079±0.0032)μg C /cm 3㊁(0.024±0.0091)μg C /cm 3㊂从CJ2站到CJ7站HMF 丰度呈现逐渐增高的趋势,但2011秋季逐渐降低,其中2011秋季CJ2站和CJ5站HMF 丰度远高于其他站位(图3,图4)㊂在表层0 8cm 沉积物中,2010年和2011年夏季各站硅藻的丰度分别为(5±4)个/cm 3㊁(24±7)个/cm 3,生物量为(0.0010±0.00076)μg C /cm 3㊁(0.0080±0.0032)μg C /cm 3㊂两年秋季丰度为(4±5)个/cm 3㊁(37±19)个/cm 3,生物量为(0.0011±0.0015)μg C /cm 3㊁(0.0072±0.0033)μg C /cm 3㊂从CJ2站到CJ7站硅藻丰度呈现先升高后降低的趋势,CJ5站多出现最高值(图3,图4)㊂本研究中共发现硅藻7属9种,优势类群均为曲舟藻(Pleurosigma )㊁菱形藻(Nitzschia )和舟形藻(Navicula )㊂在表层0 8cm 沉积物中,2010年和2011年夏季各站纤毛虫的丰度分别为(5±3)个/cm 3㊁(7±4)个/cm 3,生物量为(0.036±0.010)μg C /cm 3㊁(0.059±0.047)μg C /cm 3㊂两年秋季丰度为(5±3)个/cm 3㊁(19±10)个/cm 3,生物量为(0.066±0.058)μg C /cm 3㊁(0.17±0.16)μg C /cm 3㊂从CJ2站到CJ7站夏季时丰度先降低后增高,秋季时呈现逐渐降低的趋势㊂最低值多出现在CJ5站,但2011秋季此站出现各站次纤毛虫丰度最高值(图3,图4)㊂年度变化上,除细菌㊁蓝细菌外微型生物各类群丰度和生物量多为2011年高于2010年,其中硅藻丰度2011年秋季较2010年秋季高9倍;季节变化上,除细菌㊁蓝细菌外微型生物各类群丰度和生物量多为秋季高于夏季,其中2011年秋季HMF 的丰度是夏季的3倍㊂基于Bray⁃Curtis 相似性矩阵对微型底栖生物各类群的丰度进行CLUSTER 聚类分析,结果表明,在94.96%相似性水平上,2010年和2011年各自聚为一组,经SIMPROF 检验两个组具有极显著差异(P =0.002),且同航次的聚在一起,但不同季节间和不同站位间均无显著差异㊂2.3 小型底栖生物丰度和生物量的水平分布与时空变化2010年和2011年夏季表层0 8cm 沉积物中,各站小型底栖生物的丰度分别为(21±13)个/cm 3(相当于1653个/10cm 2)㊁(21±6)个/cm 3(1692个/10cm 2),生物量为(0.90±0.57)μg C /cm 3(72μg C /10cm 2)㊁(1.48±0.46)μg C /cm 3(118μg C /10cm 2)㊂2011年夏季生物量增加主要是CJ5㊁CJ6两站位多毛类增多㊂两年秋季丰度为(20±17)个/cm 3(1617个/10cm 2)㊁(27±12)个/cm 3(2146个/10cm 2),生物量为(0.93±0.94)μg C /cm 3(75μg C /10cm 2)㊁(0.94±0.39)μg C /cm 3(75μg C /10cm 2)㊂从CJ2站到CJ7站,2010年丰度先降低后增高,最低值出现在CJ5站;与此不同,2011年呈现逐渐降低的趋势,最高值出现在CJ2站(图3,图4)㊂在鉴定出的11个小型底栖生物类群,线虫在丰度上占绝对优势,其分布趋势与小型底栖生物一致,在小型底栖生物中所占的比例基本上从CJ2站到CJ7站呈逐渐降低的趋势㊂2010年和2011年夏季各站线虫丰度分别为(18±11)个/cm 3(1461个/10cm 2)㊁(18±7)个/cm 3(1469个/10cm 2),生物量为(0.54±0.35)μg C /cm 3(43μg C /10cm 2)㊁(0.39±0.30)μg C /cm 3(32μg C /10cm 2)㊂两年秋季丰度为(18±16)个/cm 3(1466个/10cm 2)㊁(25±11)个/cm 3(1970个/10cm 2),生物量为(0.44±0.41)μg C /cm 3(35μg C /10cm 2)㊁(0.44±0.29)μg C /cm 3(36μg C /10cm 2)㊂秋季小个体线虫比例增加㊂其他常见类群为桡足类㊁多毛类㊁动吻类等㊂小型底栖生物各类群丰度和生物量年度季节间变化均不大,多为2011年略高于2010年,线虫和腹毛类为秋季略高于夏季㊂基于Bray⁃Curtis 相似性矩阵对小型底栖生物各类群的丰度进行聚类分析(CLUSTER),结果显示无明显的聚类分组㊂2.4 微型和小型底栖生物的垂直分布2010年及2011年四航次微型和小型底栖生物的丰度及生物量随沉积物深度的增加而减少,年度和季节7505 15期 周百灵 等:长江口外海域微型和小型底栖生物群落结构和时空变化 8505 生 态 学 报 35卷 间变化均不大㊂沉积物表层0 2cm中细菌㊁蓝细菌㊁PNF㊁HNF的数量分别占各自总数的50%㊁54%㊁51%㊁54%,2 5cm分层中各类群数量分别占各自总数的50%㊁46%㊁49%㊁46%㊂HMF垂直分层较上述四类群明显,表层0 2cm分层中其数量占总数的65%,22%分布在2 5cm,13%分布在5 8cm分层㊂硅藻和纤毛虫分布在0 2cm沉积物表层的数量分别占各自总数的84%和82%,仅4%的硅藻和5%的纤毛虫分布在5 8cm分层㊂四航次各站位表层0 2cm沉积物中的纤毛虫数量的变动直接影响0 8cm沉积物中纤毛虫丰度的变化趋势㊂秋季表层0 2cm中纤毛虫数量占总数量的比值CJ2㊁CJ7略高于夏季,CJ5㊁CJ6略低于夏季㊂小型底栖生物同样主要分布在沉积物表层0 2cm(占总数的77%),17%分布在2 5cm分层,仅有6%分布在5 8cm分层㊂2.5 微型和小型底栖生物的生产力和新陈代谢率表层5cm沉积物中,通过沉积物叶绿素a含量换算的初级生产力总量为444 602mg C m-2d-1;底栖硅藻的生产力估算值为0.05 0.4mg C m-2d-1,仅占初级生产力的0.009% 0.09%㊂底栖细菌的生产力为80 101mg C m-2d-1,相当于初级生产力的17% 21%;底栖纤毛虫的平均生产力为3 23mg C m-2d-1;小型底栖生物的平均生产力为2 3mg C m-2d-1㊂就微型和小型异养类群的新陈代谢率来看,异养鞭毛虫所占的比例最高(72.4% 99.6%),其次是小型底栖生物(0.3% 27.0%),纤毛虫仅占0.01% 0.7%(表1)㊂相较2010年,异养鞭毛虫的贡献在2011年明显增大㊂表1 纤毛虫㊁异养鞭毛虫及小型底栖生物生物量及新陈代谢率的相对贡献Table1 The relative contribution of ciliates,heterotrophic flagellates and meiobenthos in biomass and metabolic rate季节Seasons生物量Biomass*/%新陈代谢率Metabolic rate*/%2010⁃07 1.3∶78.4∶20.30.3∶92.9∶6.82010⁃11 1.6∶76.9∶21.50.3∶93.0∶6.62011⁃070.2∶92.8∶7.00.1∶98.2∶1.82011⁃110.7∶96.3∶2.90.1∶99.1∶0.8 *纤毛虫∶异养鞭毛虫∶小型底栖生物;所有均为0 5cm沉积物中的数据2.6 底栖纤毛虫群落结构特征本研究中共发现纤毛虫166种,隶属14纲/亚纲,83属㊂钩刺亚纲(Haptoria)种类数最多(46种),其次是前口纲(Prostomatea,34种)㊁盾纤亚纲(Scuticociliatia,26种)㊁腹毛亚纲(Hypotrichia,14)和核残迹纲(Karyorelictea,13种)㊂前口纲在丰度及生物量上均为最优势的类群,其次为钩刺亚纲,盾纤亚纲,腹毛亚纲,篮口纲(Nassophorea)等,各类群丰度和生物量所占比例年度季节间变化均不大㊂纤毛虫在各航次㊁各站位检获的物种数范围为6 66,最小值和最大值分别出现在2011年夏季和秋季的CJ5站㊂Margalef多样性指数为2.3 12.1,Shannon⁃Wiener多样性指数在1.58 3.48之间㊂各多样性指数2011年高于2010年,秋季高于夏季㊂依据相关文献及对纤毛虫食物泡的镜检观察,检获肉食性纤毛虫74种,菌食性69种,藻食性15种,杂食性8种㊂肉食性纤毛虫主要是前口纲的裸口虫属(Holophrya)㊁钩刺亚纲及核残迹纲的代表种类,在夏季0 8cm沉积物中其丰度占总丰度的39% 54%,生物量占总生物量的36% 83%;秋季其丰度和生物量所占比例均增加(丰度占52% 62%,生物量占68% 86%)㊂菌食性纤毛虫主要为楯纤虫属(Aspidisca)㊁游仆虫属(Euplotes)㊁中圆虫属(Metacystis)及盾纤亚纲的一些类群,夏季其丰度占33% 56%,生物量占10% 61%;秋季其丰度和生物量所占比例均减小(丰度占32% 40%,生物量占9% 26%)㊂藻食性纤毛虫主要类群为佛瑞环须虫(Peritromus faurei)及带膜虫属(Zosterodasys)的类群,夏季其丰度仅占5% 8%,生物量占3% 4%;秋季比例变化不大(丰度占3% 6%,生物量占1% 5%)㊂杂食性纤毛虫丰度和生物量所占比例均最小,夏季丰度仅占0.4% 5%,生物量占0.2% 2%;秋季丰度占1% 3%,生物量占1% 3%(图5)㊂图5 长江口外海域研究站位纤毛虫各营养类型的丰度及生物量百分比Fig.5 Relative contribution of the four trophic types to ciliate abundance and biomass at the stations off the Yangtze Estuary就各站位来看,在0 8cm 沉积物中,CJ6站在2011年的夏季和秋季均以菌食性纤毛虫所占比例最多,在2010夏季则以肉食性纤毛虫为最多;除2011年夏季的CJ2站和2010年秋季的CJ5站以菌食性纤毛虫生物量所占比例最高外,这两个站位的其他航次样品中均以肉食性纤毛虫所占比例最高㊂CJ7站各航次均为肉食性所占比例最高,秋季所占比例较夏季高(图6)㊂在表层2cm 沉积物中,即使采用最大的细菌摄食率,纤毛虫仅利用细菌日现存量的0.002% 0.08%,对硅藻日现存量的消耗均达到100%以上(450% 21969%,2010秋季CJ2和2011夏季CJ5站均未检获藻食性纤毛虫)㊂图6 长江口外海域各研究站位(CJ2⁃7)纤毛虫各营养类型生物量的相对百分比Fig.6 Relative contribution of the trophic types to ciliate biomass at the stations CJ2⁃7off the Yangtze Estuary对研究站位纤毛虫群落结构进行CLUSTER 聚类分析,基本上同年度同季节的样品聚合在一起㊂单因子相似性检验(One⁃Way ANOSIM)分析结果显示,年度差异大于季节差异,大于站位差异,不同年度间(Global R =0.328,P =0.006)有极显著差异,不同季节间和不同站位间无显著差异㊂9505 15期 周百灵 等:长江口外海域微型和小型底栖生物群落结构和时空变化 2.7 微型和小型底栖生物各类群丰度与环境因子的关系Spearman相关分析表明,夏季时,PNF丰度和温度呈负相关(P=0.047),和水深呈正相关(P=0.021),纤毛虫丰度和中值粒径呈负相关关系(P=0.047),HNF和HMF丰度均与叶绿素a含量呈负相关关系(P= 0.037,P=0.047),PNF和HNF丰度与总氮含量呈正相关(P=0),桡足类㊁多毛类丰度和总氮含量呈显著正相关(P=0.016,P=0.001)㊂BIOENV分析表明,微型底栖生物群落结构与叶绿素a含量最相关(R= 0.529,P=0.006);其中,纤毛虫群落结构与底层水温㊁叶绿素a含量和有机质含量的组合最相关(R=0.414, P=0.015)㊂秋季时,HMF丰度和底层水温呈正相关(P=0.014),细菌丰度和叶绿素a含量㊁有机质含量呈正相关关系(P=0.019,P=0.005),蓝细菌丰度和水深呈负相关关系(P=0.019),与中值粒径呈正相关关系(P= 0.042),小型底栖生物丰度和线虫丰度均与盐度呈负相关(P=0.023,P=0.023)㊂BIOENV分析表明,小型底栖生物群落结构与中值粒径最相关(R=0.668,P=0.004);微型底栖生物群落结构和纤毛虫群落结构未见最相关的因子组合㊂3 讨论3.1 微型和小型底栖生物的数量和功能重要性在长江口及东海海域,对底栖细菌丰度和生物量的报道较少㊂季倩等采用流式细胞仪技术研究发现长江口及邻近海域底栖细菌的丰度为106个/g沉积物[27],其结果明显低于本研究自表层5cm沉积物中细菌的丰度,原因可能是采用流式细胞仪计数沉积物中细菌会造成对其数量的低估[8]㊂本研究自长江口外海区获得的细菌丰度略高于同海域2009年春秋季底栖细菌丰度[8],与黄海陆架区夏季细菌丰度接近[7]㊂据报道,东海水体异养细菌的丰度为105个/cm3[28],本研究单位体积沉积物中细菌较水体中细菌丰度高3个数量级㊂由此,即使在表层5cm的沉积物中,底栖细菌的总量和50m水深中的细菌相当㊂由于细菌在沉积物中的分布很深,且所研究区域大多低于50m,因此底栖细菌在数量上无疑更多㊂蓝细菌作为初级生产者在生态系统中具有重要作用,但目前较少工作涉及底栖蓝细菌的生态学研究㊂国外已有研究表明,不足4m的浅水区域沉积物中蓝细菌的丰度为107 108个/cm3[6],明显较本研究所获蓝细菌丰度高,主要原因可能是所研区域水体透明度不高,且平均水深逾40m,海底光线弱,不利于蓝细菌的繁殖㊂与同海域研究结果相较,本研究所获蓝细菌数量高于2009年春季的数据[8],与秋季的数据较为接近,但在平均值上略低㊂此外,与黄海陆架区夏季数据较为接近[7],平均值较后者略高㊂季倩报道长江口海域夏季水体表层蓝细菌的丰度为10 105个/mL[27],相较于此,本研究单位体积沉积物中蓝细菌的数量高1 5个数量级㊂在潮间带,底栖异养鞭毛虫的丰度为103 105个/cm3,主要为微型鞭毛虫[6]㊂本研究结果与其相一致,但丰度(105 106个/cm3)较潮间带的高㊂本研究所获微型鞭毛虫丰度和生物量与同海域春秋季(丰度105 106个/cm3,生物量4 21μg C/cm3)[8]和黄海陆架区夏季(丰度106个/cm3,生物量19 41μg C/ cm3)[7]的数据接近,而HMF丰度相较偏低㊂对黄东海春夏季浮游鞭毛虫的研究显示,各水层微型鞭毛虫的总丰度为102个/mL,生物量为20μg C/L[15]㊂如此,本研究单位体积沉积物中微型鞭毛虫较水体中浮游鞭毛虫丰度高3 4个数量级,生物量较后者高3个数量级㊂由此,即使表层5cm沉积物中,该海域底栖鞭毛虫的绝对数量也明显较水柱中的总量高㊂在浮游生态系统中,异养鞭毛虫是重要的细菌摄食者,影响细菌的现存量及生态系统的净生产量㊂已有研究表明,在海洋水体中细菌和HNF丰度比值多为1000∶1,在此比值下两者在食物链中维持在一个稳定的状态[29]㊂而在海洋潮间带的研究表明,细菌和HNF丰度比值可达104 106∶1,鞭毛虫摄食不是影响细菌生产力的重要因素[4,30⁃31]㊂与浮游生态系统相似,本研究中2010年夏季和秋季细菌和HNF丰度比值量级为1000∶1,而2011年夏季和秋季两者比值均约为100∶1,在这种情况下HNF可对细菌的现存量产生明显的摄食影响㊂。