基于三链DNA结构的01整数规划改进研究任晓玲
近100分的Nature顶级子刊重磅综述揭秘这个研究热点!快收藏!

近100分的Nature顶级子刊重磅综述揭秘这个研究热点!快收藏!近百分重磅综述本文首发于君莲书院,扫码入群抢先体验,第一时间学习前沿热点,高分综述。
非编码RNA在近几年最为火爆,从miRNA到lncRNA,再到circRNA,一直都是热潮。
今天我们就来回顾一下lncRNA的基因调控和生物学功能。
该综述由中科院陈玲玲教授和西班牙纳瓦拉大学Maite Huarte教授为共同通讯作者于今年2月份发表在Nature Review. Molecular Cell Biology期刊(最新影响因子为94.444分)上,截至10月23日该文已被引用147次(数据来源:PubMed)。
作者指出过去十年积累的证据表明,长非编码RNA(lncRNAs)广泛表达并在基因调控中发挥关键作用。
最近的研究已经开始揭示lncRNAs的生物发生与mRNAs的不同之处,并与其特定的亚细胞定位和功能相关。
lncRNAs根据其定位及其与DNA、RNA和蛋白质的特异性相互作用,可以调节染色质功能,调节无膜核体的组装和功能,改变细胞质mRNAs的稳定性和翻译,干扰信号通路。
其中许多功能最终会在不同的生物学和生理病理学环境中影响基因表达。
组织特异性和条件特异性表达模式表明lncRNAs是潜在的生物标记物,并为临床靶向它们提供了理论依据。
在这篇综述中,研究人员讨论了lncRNA 的生物发生机制、在转录、转录后和其他基因调控模式中的定位和功能,以及它们潜在的治疗应用。
现在就让我们来一起看看吧。
前言基因组被广泛转录并产生数千个lncRNAs,它们被定义为长度超过200个核苷酸的RNA,不能翻译成功能性蛋白质。
这一宽泛的定义涵盖了一个巨大且高度异质的转录物集合,这些转录物在其生物成因和基因组起源上存在差异。
来自人类基因编码的统计数据表明,人类基因组包含16000多个lncRNAs,但其他估计超过100000个人类lncRNAs。
这些主要包括RNA聚合酶II(Pol II)转录的lncRNAs,也包括其他RNA聚合酶转录的lncRNAs;基因间区的lncRNAs (lincRNAs)以及与其他基因重叠的正反义转录本。
数字金融的风险、挑战与应对

41CENTRAL ISSUE区域治理作者简介:任晓珠,生于1989年,副教授,金融硕士,研究方向为数字金融、金融市场、国际金融。
基金项目:重庆市教育委员会科学技术研究计划项目《基于区块链技术的重庆内陆国际金融中心建设研究》(项目编号:KJQN202002002)、重庆市教育委员会人文社会科学研究项目《“上云用数赋智”下商业银行小微金融数字化发展研究》(项目编号:21SKGH306)、《“双循环”视角下成渝双城经济圈建设中四位协同产业体系研究》(项目编号:21SKGH307)、4.重庆市高等教育教学改革研究项目《基于智慧学习工场的”135递阶“新金融人才培养研究与实践》(项目编号:202120)、重庆对外经贸学院教育教学改革研究产教融合专项《“数字经济专业站”助推经贸类人才培养模式研究》(项目编号:JGCJ202005)阶段性研究成果数字金融的风险、挑战与应对*重庆对外经贸学院 任晓珠摘要:一场疫情改变了人类生产生活方方面面,一次技术性变革推动数字经济新业态的到来。
当前,我国经济发展最为活跃的领域当属数字经济领域,与之相关的各种商业模式和各类技术创新速度可谓日新月异,数字金融正成为这其中的佼佼者。
事实上任何新事物被人接受都需要一段从陌生到熟悉的过渡期,但是数字金融从出现到普及就目前来看需要思考一个问题,它是否过快了?本文在此背景下研究数字金融,以期对数字金融进行风险分析,并阐明可能遇到的挑战,并据此提出数字金融可持续发展的可行性对策。
关键词:数字金融;技术风险;金融监管中图分类号:F83文献标识码:A文章编号:2096-4595(2021)02-0041-0002一、数字金融的发展趋势(一)“数字金融1.0”时代数字金融数字技术与传统金融融合的新金融业态。
数字金融在中国乃至全世界都是新生事物。
在中国,数字金融可溯源至2004年,这一年支付宝出生,中国数字金融元年应当是2013年,此时余额宝正式上线。
中国沙棘HrNHX6基因的生物信息学分析及其在盐胁迫下的表达模式

㊀山东农业科学㊀2024ꎬ56(1):17~23ShandongAgriculturalSciences㊀DOI:10.14083/j.issn.1001-4942.2024.01.003收稿日期:2023-02-24基金项目:国家自然科学基金项目(31660071)ꎻ青海省 昆仑英才 高端创新创业人才 计划项目作者简介:肖莎莎(1999 )ꎬ女ꎬ重庆人ꎬ硕士研究生ꎬ主要从事林木遗传育种方面的研究ꎮE-mail:1521621909@qq.com通信作者:马玉花(1978 )ꎬ女ꎬ博士ꎬ教授ꎬ主要从事森林培育理论与技术㊁植物资源开发利用研究ꎮE-mail:qhxnmyh@163.com中国沙棘HrNHX6基因的生物信息学分析及其在盐胁迫下的表达模式肖莎莎ꎬ费凡ꎬ董佳伟ꎬ张丹ꎬ马玉花(青海大学农牧学院ꎬ青海西宁㊀810016)㊀㊀摘要:Na+/H+逆向转运蛋白(NHX)广泛存在于多种生物中ꎬ可调节植物响应盐胁迫ꎮ本研究对中国沙棘HrNHX6进行结构预测和分析ꎬ并采用qRT-PCR技术分析不同浓度NaCl胁迫下HrNHX6基因的表达情况ꎮ结果如下:①HrNHX6基因总长为1347bpꎬ与月季花等植物NHX6基因具有较高的同源性ꎮ②HrNHX6为稳定的疏水性蛋白ꎬ定位于高尔基体膜中ꎬ不具有信号肽ꎬ含7个跨膜螺旋结构ꎬ且α-螺旋和无规则卷曲为其二级结构的主要构成元件ꎬ具有多个修饰位点ꎮ③盐胁迫下中国沙棘根㊁茎㊁叶中的HrNHX6基因表达上调ꎬ说明HrNHX6基因在中国沙棘响应盐胁迫的过程中可能发挥着重要的调控作用ꎮ本研究结果可为阐明中国沙棘应对盐胁迫的机制奠定基础ꎮ关键词:中国沙棘ꎻ盐胁迫ꎻHrNHX6基因ꎻ荧光定量PCRꎻ基因表达中图分类号:S793.6ꎻQ786㊀㊀文献标识号:A㊀㊀文章编号:1001-4942(2024)01-0017-07BioinformaticsAnalysisofGeneHrNHX6inHippophaerhamnoidessubsp.sinensisanditsExpressionPatternunderSaltStressXiaoShashaꎬFeiFanꎬDongJiaweiꎬZhangDanꎬMaYuhua(CollegeofAgricultureandAnimalHusbandryꎬQinghaiUniversityꎬXining810016ꎬChina)Abstract㊀TheNa+/H+reversetransporter(NHX)proteinswhichcouldregulateplantresponsestosaltstressarewidelypresentinavarietyoforganisms.InthispaperꎬthestructureofHrNHX6fromHippophaerh ̄amnoidessubsp.sinensiswaspredictedandanalyzedꎬandtheexpressionofthegeneunderdifferentconcentra ̄tionsofNaClstresswasassayedbyqRT ̄PCR.Theresultswereasfollows.①ThetotallengthofHrNHX6genewas1347bpꎬwhichhadhigherhomologouswiththeNHX6ofRosachinensiet.②HrNHX6wasastablehy ̄drophobicproteinlocatedintheGolgimembranewithnosignalpeptidebutseventransmembranehelicalstruc ̄turesꎻα ̄helixandirregularcurlwerethemaincomponentsofitssecondarystructureꎬandithadmanymodifi ̄cationsites.③qRT ̄PCRanalysisshowedthattheexpressionsofHrNHX6genewereup ̄regulatedinrootsꎬstemsandleavesofH.rhamnoidesundersaltstressꎬwhichindicatedanimportantregulatoryroleofthegeneinresponsetosaltstress.TheresultsabovecouldlayafoundationforelucidatingtheresponsemechanismsofH.rhamnoidestosaltstress.Keywords㊀Hippophaerhamnoidessubsp.sinensisꎻSaltstressꎻHrNHX6geneꎻqRT ̄PCRꎻGeneexpression㊀㊀土壤盐碱化是影响农业发展的全球性问题ꎬ由于工业污染㊁化肥过量使用和灌溉不合理等因素的影响ꎬ土壤盐碱化的程度不断上升[1]ꎬ已成为影响植物生长的重要问题之一ꎮ盐胁迫不仅会抑制植物种子的萌发ꎬ还会加速植物根系的木栓化[2]ꎬ影响根系对水分和营养物质的摄入ꎬ从而抑制植物生长发育ꎮ植物耐盐性可表现为Na+被Na+/H+逆向转运蛋白(NHX)外排或区隔化以维持细胞内的低Na+水平[3]ꎮ研究表明ꎬ在盐胁迫下ꎬ植物利用NHXs将Na+分离到液泡ꎬ可以促使细胞从外界应激环境中吸收水分以维持渗透平衡ꎬ调节细胞质中的Na+浓度和pHꎬ保护细胞器[4-5]ꎮNHX基因已在许多植物中被鉴定出来ꎬ但不同植物其成员数量不同ꎬ例如ꎬ拟南芥中有8个NHX基因[6]ꎬ葡萄中有6个NHX基因[7]ꎬ棉花中有25个NHX基因[8]ꎮ拟南芥的8个NHX蛋白中ꎬAtNHX1 At ̄NHX4定位于液泡膜上ꎬAtNHX5和AtNHX6定位于胞体内膜ꎬAtNHX7(AtSOS1)和AtNHX8定位于质膜上ꎻ根据亚细胞定位ꎬ又可将它们分为液泡NHXs(vacuolarNHXs)㊁内膜NHXs(endosomalNHXs)㊁质膜NHXs(plasmamembraneNHXs)[6]ꎮAtNHX1 AtNHX4通过形成的跨膜质子梯度提供能量ꎬ介导Na+区隔化ꎬ以减少过量盐离子对细胞的伤害ꎬ稳定细胞渗透压ꎬ使植物适应盐旱环境ꎻAtNHX5和AtNHX6具体定位于跨高尔基体网络ꎬ可参与囊泡运输和细胞扩增活性ꎬ特别是在液泡运输中具有重要作用ꎬ并调节pH和K+ꎻAt ̄NHX7通过盐超敏感(saltoverlysensitivityꎬSOS)信号传导途径赋予植物耐盐性ꎬSOS途径包括SOS1(质膜Na+/H+反向运输蛋白)㊁SOS2(丝氨酸/苏氨酸蛋白激酶)和SOS3(钙结合蛋白)ꎬ其中SOS2与SOS3的复合物可调节SOS1以去除细胞内的Na+[9-10]ꎻAtNHX8可以参与Li+/H+反向转运[11-12]ꎮ但目前有关NHXs在中国沙棘应对盐胁迫时的作用机制研究报道相对较少ꎮ中国沙棘(Hippophaerhamnoidessubsp.sinen ̄sis)是一种胡颓子科(Elaeagnaceae)沙棘属(Hipp ̄ophae)植物ꎬ具有耐旱㊁抗寒㊁抗风沙等特性ꎬ在我国西北被广泛用于水土保持ꎮ研究表明ꎬ在盐胁迫条件下ꎬ沙棘渗透压降低ꎬ体内脯氨酸㊁可溶性糖等渗透介质的含量增加ꎬ渗透调节能力增强ꎬ以使植株适应外界环境[13]ꎻ另一方面ꎬ沙棘体内抗氧化酶(POD㊁CAT㊁SOD等)活性增强ꎬ可减轻活性氧对植株的伤害[14]ꎬ从而提高沙棘的耐盐性ꎮ本研究以中国沙棘的抗盐分子调节机制为切入点ꎬ对耐盐相关基因HrNHX6及其编码蛋白进行结构分析和亚细胞定位预测ꎬ同时分析其在不同浓度NaCl胁迫下的表达模式ꎬ旨在为中国沙棘应对盐胁迫的机制阐明奠定基础ꎬ同时为高效耐盐基因的发掘提供依据ꎮ1㊀材料与方法1.1㊀试验材料供试植物材料为长势一致的中国沙棘两年生扦插苗ꎬ由青海省西宁市大通县城关苗圃(37ʎ14ᶄ45ᵡNꎬ101ʎ30ᶄ15ᵡEꎬ海拔2920m)提供ꎮ2019年7月28日开始盐胁迫试验ꎬ将18盆中国沙棘扦插苗随机分为6组ꎬ分别用500mL浓度为0㊁200㊁400㊁600㊁800㊁1000mmol/L的NaCl溶液浇灌ꎬ3天后再浇灌一次ꎮ每组处理的3盆作为3个重复ꎬ试验时在花盆底放置一个托盘ꎬ及时将漏出的溶液倒回花盆中ꎬ以防止水分和盐分的流失ꎮ8月2日采集根㊁茎㊁叶ꎬ用超纯水冲洗后用滤纸吸干水分ꎬ装入冻存管ꎬ置于液氮中速冻ꎬ随后放入-80ħ冰箱保存备用ꎮ1.2㊀试验方法将中国沙棘根㊁茎和叶在液氮中研磨成细粉ꎬ依照植物RNA试剂盒说明书快速提取总RNAꎬ使用PrimeScript Ⅱ1stStrandcDNASynthesisKit反转录质量检测合格的优质RNAꎬ并通过荧光定量PCR法分析表达模式ꎬ引物(HrNHX6-F:CAATGGATATGTTGCTCCTTCGTꎬHrNHX6-R:CTAATGCTGTAAATGATGCTGTGCT)由宝生物工程(大连)有限公司合成ꎮ使用2-ΔΔCt法对基因的相对表达量进行分析ꎬ使用MicrosoftExcel2010和SPSS22软件进行数据整理与统计分析ꎮHrNHX6基因的全长序列通过转录组测序获得ꎮ用DNASTAR进行核苷酸序列分析和同源比对ꎻ用MEME进行保守基序分析ꎻ用ExPASyProt ̄Param进行蛋白理化性质分析ꎻ用PredictProtein进行亚细胞定位ꎻ用NetOGlyc4.0Server进行信号肽和跨膜螺旋区分析ꎻI-TASSER用于预测蛋白二级结构ꎬSWISS-MODEL用于预测三级结构ꎻPredictProtein用于蛋白功能位点的预测ꎮ2㊀结果与分析2.1㊀HrNHX6基因序列分析2.1.1㊀HrNHX6基因的核苷酸序列分析㊀HrN ̄HX6基因序列长度为1347bpꎬ含343个A㊁288个G㊁470个T㊁246个CꎬA+T的含量(60.36%)高于G+C的含量(39.64%)ꎮ81山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第56卷㊀2.1.2㊀HrNHX6蛋白的氨基酸序列分析㊀HrN ̄HX6基因编码448个氨基酸ꎬ其中ꎬ酸性氨基酸(D㊁E)36个ꎬ碱性氨基酸(K㊁R)22个ꎬ极性氨基酸(N㊁C㊁Q㊁S㊁T㊁Y)119个ꎬ疏水性氨基酸(A㊁I㊁L㊁F㊁W㊁V)190个ꎬ分别占氨基酸总量的8.0%㊁5.0%㊁26.6%㊁42.4%ꎮ在该蛋白中Leu(L)和Ser(S)含量较高ꎬ所占比例分别为12.3%和9.8%ꎮ2.1.3㊀HrNHX6基因的核苷酸和氨基酸序列同源性分析㊀将HrNHX6与10种植物NHX6基因的核苷酸和氨基酸序列进行同源比对分析ꎬ结果(图1㊁图2)表明ꎬHrNHX6与其他植物NHX6基因核苷酸序列的同源性为82.9%~85.7%ꎬ氨基酸序列的同源性则达到84.9%~89.7%ꎬ其中中国沙棘与月季花的序列同源性最高ꎮ中国沙棘:Hippophaerhamnoidesꎻ茶树:Camelliasinensisꎻ橡胶树:Heveabrasiliensisꎻ麻疯树:Jatrophacurcasꎻ欧洲野苹果:Malussylvestrisꎻ杧果:Mangiferaindicaꎻ木薯:Manihotesculentaꎻ阿月浑子:Pistaciaveraꎻ毛白杨:Populustomentosaꎻ山谷白栎:Quercuslobataꎻ月季花:Rosachinensisꎮ下图同ꎮ图1㊀中国沙棘与其他10种植物NHX6基因的同源比对图2㊀中国沙棘与其他10种植物NHX6蛋白的同源比对2.1.4㊀HrNHX6基因保守基序分析㊀通过MEME软件对HrNHX6的保守基序进行分析ꎬ共得到15个保守结构域ꎬ每个结构域的长度为50个保守氨基酸ꎬ具有11个位点ꎬ且E值都显著ꎮHrNHX6保守基序与橡胶树㊁山谷白栎㊁欧洲野苹果等植物NHX6基因保守基序完全一致(图3)ꎮ2.2㊀HrNHX6蛋白结构预测2.2.1㊀HrNHX6蛋白理化性质及亚细胞定位㊀HrNHX6蛋白分子式为C2260H3436N552O636S20ꎬ分子量为49156.84Daꎬ理论等电点为5.475ꎬ脂肪系91㊀第1期㊀㊀㊀㊀肖莎莎ꎬ等:中国沙棘HrNHX6基因的生物信息学分析及其在盐胁迫下的表达模式数为99.22ꎬ蛋白质不稳定系数为35.02ꎬ为稳定蛋白ꎮHrNHX6蛋白的疏水分布区域较大ꎬ且有多个连续疏水区ꎬ疏水性最大值为3.589ꎬ而亲水区域较小ꎬ亲水性最大值为-2.511(图4)ꎬ具有跨膜蛋白疏水性较强的特征ꎬ表明该蛋白是一种疏水性蛋白ꎮ图3㊀NHX6的保守基序02山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第56卷㊀图4㊀HrNHX6蛋白亲水性2.2.2㊀信号肽预测㊀NetOGlyc4.0Server分析表明ꎬ切割位点C的最大值为0.243ꎬ位于第33位氨基酸ꎻ信号肽分数S的最大值为0.179ꎬ位于第44位氨基酸ꎻ合并切割位点Y的最大值为0.164ꎬ位于第33位氨基酸(图5)ꎮS的平均值为0.117ꎬ预测剪切点在1~28位氨基酸ꎮS的平均值远低于0.5ꎬ表明HrNHX6蛋白是一种没有潜在信号肽位点的非分泌蛋白ꎮ2.2.3㊀HrNHX6蛋白结构及功能位点预测㊀HrN ̄HX6蛋白二级结构中α-螺旋㊁无规则卷曲㊁延伸链㊁β-折叠分别占49.33%㊁33.26%㊁13.62%㊁3.79%ꎬα-螺旋和无规则卷曲是其主要构成成分(图6)ꎮ蛋白三维构象如图7所示ꎮ跨膜区预测结果(图8)显示ꎬHrNHX6含7个跨膜螺旋结构ꎬ属于跨膜蛋白ꎮHrNHX6蛋白功能位点的预测结果显示该蛋白包含多个修饰位点:2个N-糖基化位点ꎬ即始于215位氨基酸的NLSE和始于375位的NESFꎻ2个蛋白激酶C磷酸化位点ꎬ分别为始于22位的SPK和始于220位的SQRꎻ5个酪蛋白激酶Ⅱ磷酸化位点ꎬ分别为始于77位的SATDꎬ始于233位的SLAEꎬ始于317位的SIHDꎬ始于350位的TM ̄LEꎬ始于414位的TALDꎻ9个N-豆寇酰基化位点ꎬ分别为始于31位的GAIVTFꎬ始于40位的GTFIASꎬ始于73位的GSLISAꎬ始于92位的GSD ̄VNLꎬ始于145位的GSLSAGꎬ始于152位的GVG ̄FTSꎬ始于192位的GLGLSGꎬ始于304位的GLR ̄GAMꎬ始于345位的GGSTGTꎮ图5㊀HrNHX6的信号肽预测图6㊀HrNHX6蛋白二级结构预测2.3㊀不同浓度NaCl胁迫下HrNHX6基因的表达模式分析NaCl胁迫下ꎬ中国沙棘根㊁茎㊁叶中的HrNHX6基因均上调表达(图9)ꎮ在中国沙棘叶中ꎬ其表达量随着NaCl浓度的增加逐渐升高ꎬ当NaCl浓度超过600mmo/L后ꎬ表达量显著升高ꎻ在茎中ꎬ其表达量呈现先升高后降低的趋势ꎬ在400mmol/LNaCl处理下最高ꎬNaCl浓度继续升高ꎬ其表达量略有下降且变化趋于平缓ꎬ但均显著高于0~200mmol/LNaCl处理ꎻ在根中ꎬ其表达量则表现为波动上升趋势ꎬ在1000mmol/LNaCl处理下达到最高ꎬ除与600mmol/LNaCl处理差异不显著外ꎬ显著高于其他处理ꎮ12㊀第1期㊀㊀㊀㊀肖莎莎ꎬ等:中国沙棘HrNHX6基因的生物信息学分析及其在盐胁迫下的表达模式图7㊀HrNHX6蛋白三维结构模型图8㊀HrNHX6蛋白跨膜结构域图9㊀不同浓度NaCl胁迫下HrNHX6基因在㊀㊀中国沙棘不同部位中的表达情况3㊀讨论与结论本研究结果表明ꎬ中国沙棘HrNHX6基因序列全长为1347bpꎬ有15个保守结构域ꎻHrNHX6为稳定蛋白ꎬ定位于高尔基体膜中ꎬ这与拟南芥中AtNHX6定位于胞体如高尔基体内膜[15]的结论一致ꎮ高尔基体网络被认为是细胞成分运输的中心ꎬ连接到内质网㊁液泡和质膜[16]ꎮ在盐胁迫下ꎬ植物产生大量的次生代谢物并分离破坏细胞结构的盐离子ꎬ这都取决于高尔基体网络的作用ꎮ因此推测HrNHX6的耐盐作用机制可能依赖于高尔基体网络的功能ꎮHrNHX6蛋白的疏水性较强ꎬ具有多个疏水区ꎬ且含有7个跨膜结构域ꎮ此外ꎬHrNHX6蛋白为非分泌蛋白ꎬ没有潜在的信号肽位点ꎬ这与杨平等[17]的研究结果一致ꎬ符合植物NHX的基本特征ꎮHrNHX6蛋白二级结构的主要构成元件为α-螺旋与无规则卷曲ꎬ分别占49.33%和33.26%ꎮα-螺旋靠链内氢键维持ꎬ两侧分别由疏水性和亲水性氨基酸组成ꎮ疏水侧通过疏水键与磷脂双分子层相互作用将蛋白固定在膜上使得HrNHX6蛋白具有稳定跨膜结构域ꎬ亲水侧则形成空桶状结构使得HrNHX6蛋白可进行水分跨膜运输ꎮ这些结构特征赋予了HrNHX6蛋白高效转运水分的功能ꎬ与该蛋白具有7个跨膜螺旋结构的分析结果一致ꎮ此外ꎬ功能位点预测表明ꎬ中国沙棘HrN ̄HX6具有2个N-糖基化位点㊁2个蛋白激酶C磷酸化位点㊁5个酪蛋白激酶Ⅱ磷酸化位点㊁9个N-豆寇酰基化位点ꎬ说明HrNHX6蛋白可通过糖基化㊁磷酸化㊁豆蔻酰化等修饰调节该蛋白的功能和构造进而完成Na+的转运ꎮ植物NHX在耐盐㊁调节pH㊁保持K+稳态以及细胞扩张㊁细胞囊泡的转运等方面发挥着关键作用ꎬ可增强石榴[18]㊁玉米[19]㊁桃[20]等的耐盐性ꎮ盐胁迫下ꎬ苜蓿MtNHX6基因在不同组织中的表达水平不同ꎬ且在叶片和根部的表达量较高[21]ꎻ玉米ZmNHX6基因在根中显著表达[19]ꎻ而在拟南芥幼苗中几乎检测不到AtNHX6基因的表达[22]ꎮ这些结果表明不同植物的NHX6基因在盐胁迫下行使功能的主要组织部位不同ꎮ本研究结果表明ꎬNaCl胁迫下HrNHX6基因在中国沙棘根㊁茎㊁叶中均上调表达ꎬ这与在石榴[18]和拟南芥[23]等植物中的研究结果一致ꎻ但不同NaCl浓度下其表达量差异较大ꎬ且同一胁迫浓度下其在不同组织中的表达也不同ꎮ此外ꎬ中国沙棘HrN ̄HX6基因受盐胁迫显著诱导ꎬ表明中国沙棘HrN ̄HX6基因在盐胁迫中行使重要的生物学功能ꎮSandhu等[21]研究发现ꎬ盐胁迫下MtNHX6基因在苜蓿根部上调表达的幅度显著高于其他组织ꎮ本研究亦得到相似结果ꎬ这可能是由于根组织最先接触到盐胁迫的缘故ꎮ另外ꎬ根中HrNHX6基因表达显著上升有助于根中细胞的Na+外排或区隔22山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第56卷㊀化ꎬ以维持细胞内低Na+水平ꎬ从而减轻盐胁迫对植物地上部分的伤害ꎬ同时通过渗透调节促进根系吸水ꎮ本研究结果可为中国沙棘HrNHX6基因功能研究提供一定的理论参考ꎬ并为中国沙棘耐盐育种基因的挖掘奠定基础ꎮ参㊀考㊀文㊀献:[1]㊀ZhuJK.Plantsalttolerance[J].TrendsinPlantScienceꎬ2001ꎬ6(2):66-71.[2]㊀TattiniMꎬGucciRꎬCoradeschiMAꎬetal.Growthꎬgasex ̄changeandioncontentinOleaeuropaeaplantsduringsalinitystressandsubsequentrelief[J].PhysiologiaPlantarumꎬ1995ꎬ95(2):203-210.[3]㊀ApseMPꎬBlumwaldE.Na+transportinplants[J].FEBSLettersꎬ2007ꎬ581(12):2247-2254.[4]㊀YamaguchiTꎬFukada ̄TanakaSꎬInagakiYꎬetal.Genesen ̄codingthevacuolarNa+/H+exchangerandflowercoloration[J].PlantandCellPhysiologyꎬ2001ꎬ42(5):451-461. [5]㊀ApseMPꎬAharonGSꎬSneddenWAꎬetal.SalttoleranceconferredbyoverexpressionofavacuolarNa+/H+antiportinArabidopsis[J].Scienceꎬ1999ꎬ285(5431):1256-1258. [6]㊀BrettCLꎬDonowitzMꎬRaoR.Evolutionaryoriginsofeukary ̄oticsodium/protonexchangers[J].AmericanJournalofPhysi ̄ology:CellPhysiologyꎬ2005ꎬ288(2):C223-C239. [7]㊀AyadiMꎬMartinsVꎬBenAyedRꎬetal.Genomewideidenti ̄ficationꎬmolecularcharacterizationꎬandgeneexpressionana ̄lysesofgrapevineNHXantiporterssuggesttheirinvolvementingrowthꎬripeningꎬseeddormancyꎬandstressresponse[J].Bi ̄ochemicalGeneticsꎬ2020ꎬ58:102-128.[8]㊀AkramUꎬSongYꎬLiangCꎬetal.Genome ̄widecharacteriza ̄tionandexpressionanalysisofNHXgenefamilyundersalinitystressinGossypiumbarbadenseanditscomparisonwithGossypi ̄umhirsutum[J].Genesꎬ2020ꎬ11(7):803. [9]㊀QiuQSꎬGuoYꎬDietrichMAꎬetal.RegulationofSOS1ꎬaplasmamembraneNa+/H+exchangerinArabidopsisthalianaꎬbySOS2andSOS3[J].ProceedingsoftheNationalAcademyofSciencesꎬ2002ꎬ99(12):8436-8441.[10]QuinteroFJꎬOhtaMꎬShiHꎬetal.ReconstitutioninyeastoftheArabidopsisSOSsignalingpathwayforNa+homeostasis[J].ProceedingsoftheNationalAcademyofSciencesꎬ2002ꎬ99(13):9061-9066.[11]WangMꎬZhengQSꎬShenQRꎬetal.Thecriticalroleofpo ̄tassiuminplantstressresponse[J].InternationalJournalofMolecularSciencesꎬ2013ꎬ14(4):7370-7390.[12]MengKꎬWuY.FootprintsofdivergentevolutionintwoNa+/H+typeantiportergenefamilies(NHXandSOS1)inthege ̄nusPopulus[J].TreePhysiologyꎬ2018ꎬ38(6):813-824. [13]阮成江ꎬ谢庆良.盐胁迫下沙棘的渗透调节效应[J].植物资源与环境学报ꎬ2002ꎬ11(2):45-47.[14]乔枫ꎬ耿贵工.盐碱胁迫对沙棘种子萌发及幼苗抗氧化酶活性的影响[J].东北林业大学学报ꎬ2012ꎬ40(2):17-19. [15]BassilEꎬOhtoMꎬEsumiTꎬetal.TheArabidopsisintracellularNa+/H+antiportersNHX5andNHX6areendosomeassociatedandnecessaryforplantgrowthanddevelopment[J].ThePlantCellꎬ2011ꎬ23(1):224-239.[16]LarkinHꎬCostantinoSꎬSeamanMNJꎬetal.Calnucfunctioninendosomalsortingoflysosomalreceptors[J].Trafficꎬ2016ꎬ17(4):416-432.[17]杨平ꎬ蔡小宁ꎬ贲爱玲.盐芥ThNHX1基因的克隆及序列分析[J].江苏农业学报ꎬ2007ꎬ23(6):556-563.[18]DongJMꎬLiuCYꎬWangYYꎬetal.Genome ̄wideidentifi ̄cationoftheNHXgenefamilyinPunicagranatumL.andtheirexpressionalpatternsundersaltstress[J].Agronomyꎬ2021ꎬ11(2):264.[19]ZörbCꎬNollAꎬKarlSꎬetal.MolecularcharacterizationofNa+/H+antiporters(ZmNHX)ofmaize(ZeamaysL.)andtheirexpressionundersaltstress[J].JournalofPlantPhysiolo ̄gyꎬ2005ꎬ162(1):55-66.[20]李淼ꎬ李桂祥ꎬ刘伟.桃NHX基因家族的全基因组鉴定与表达分析[J].山东农业科学ꎬ2021ꎬ53(12):8-16. [21]SandhuDꎬPudusseryMVꎬKaundalRꎬetal.Molecularchar ̄acterizationandexpressionanalysisoftheNa+/H+exchangergenefamilyinMedicagotruncatula[J].Functional&Integra ̄tiveGenomicsꎬ2018ꎬ18:141-153.[22]YokoiSꎬQuinteroFJꎬCuberoBꎬetal.DifferentialexpressionandfunctionofArabidopsisthalianaNHXNa+/H+antiportersinthesaltstressresponse[J].ThePlantJournalꎬ2002ꎬ30(5):529-539.[23]WangLGꎬWuXXꎬLiuYFꎬetal.AtNHX5andAtNHX6controlcellularK+andpHhomeostasisinArabidopsis:threeconservedacidicresiduesareessentialforK+transport[J].PLoSONEꎬ2015ꎬ10(12):e0144716.32㊀第1期㊀㊀㊀㊀肖莎莎ꎬ等:中国沙棘HrNHX6基因的生物信息学分析及其在盐胁迫下的表达模式。
基于深度学习的内容推荐算法研究综述

协同过滤推荐可以分为基于近邻和基于模型的推荐
方 法 ,是 利 用 用 户 物 品 交 互 数 据(比 如 评 分 数 据)实
现 推 荐(预 测)的 推 荐 方 法 ;混 合 推 荐 是 通 过 组 合 不
同的模型弥补模型之间的缺点来提高整个系统推荐
性 能 的 一 种 方 法[16]。 本 节 主 要 从 推 荐 的 算 法 思 路 、
0 概述
信息网络的迅速发展使得数据规模呈现爆发式
增 长 ,各 种 应 用 软 件(如 电 子 商 务 平 台 等)对 数 据 的 依 赖 性 越 来 越 强[1]。 但 是 ,数 据 的 增 量 超 过 了 平 台 或 系 统 的 承 受 范 围 ,这 种 现 象 被 称 为“ 信 息 过 载 ”问
近 年 来 ,应 用 软 件 的 数 量 呈 现 爆 发 式 增 长 ,滋 生 了越来越多的多源异构数据。这些数据来源多样、 结 构 复 杂 ,传 统 机 器 学 习 方 法 无 法 有 效 地 提 取 其 中 的 表 征 。 面 临 数 据 带 来 的 一 系 列 问 题 ,深 度 学 习 对 多源异构数据表征的充分学习获得了广泛的关注, 迅速成为研究热点之一。深度学习作为前沿技术, 泛指端到端的非线性可微模型。其优势主要体现在 非 线 性 转 换 、深 层 特 征 学 习 、高 弹 性 及 可 用 性 等 3 个 方 面 。 在 学 术 研 究 层 面 ,深 度 学 习 的 快 速 发 展 激 发 了 学 者 对 推 荐 领 域 学 术 研 究 的 兴 趣 ,逐 渐 出 现 了 Wide&Deep[9]、CDL[10]、AutoRec[11]等 经 典 的 深 度 推 荐 模 型 。 在 工 业 应 用 层 面 ,基 于 深 度 学 习 应 用 的 推 荐 算 法 不 断 改 进 与 创 新[12-14],为 各 领 域 的 产 品 提 供 商业化服务。
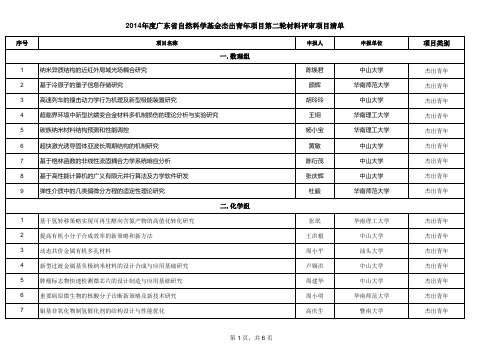
2014年度广东省自然科学基金第二轮杰出青年项目清单
第 3 页,共 6 页
20 21 22 23
新型高级氧化体系的构建及对水中毒性污染物的控制 原位研究SOFC纳米钴铁合金-陶瓷复合阳极的耐硫抗积碳机理 高效超分子水凝胶基因载体的研究 高容量锂离子电池用硅/碳复合负极材料的研发
方晶云 杨成浩 马栋 唐永炳
中山大学 华南理工大学 暨南大学 中国科学院深圳先进技术 研究院
第 4 页,共 6 页
17 18 19
基于对象典型性的评论推荐技术研究 对最优中短长度信道编码的研究 新一代广播电视传输技术研究
蔡毅 陈立 杨昉
华南理工大学 中山大学 深圳清华大学研究院
杰出青年 杰出青年 杰出青年
七.管理组
1 2 3 4 面向新服务经济的服务能力与策略设计研究 多尺度碳市场周期辨识与风险管理方法及应用研究 基础设施投资质量:决定机制、企业影响及最优配置 知识溢出与创新地理:基于新新经济地理学视角 冯博 朱帮助 张光南 陶锋 华南理工大学 五邑大学 中山大学 暨南大学 杰出青年 杰出青年 杰出青年 杰出青年
杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年
第 6 页,共 6 页
第 1 页,共 6 页
8 9
孕妇对邻苯二甲酸酯类增塑剂的暴露及室内空气对其暴露的影响 呼气直接质谱分析新技术在石化苯系物暴露的应用研究
郭英 李雪
暨南大学 暨南大学
杰出青年 杰Leabharlann 青年三.农学组1 2 3 4 5 6 7 8 9 10 华南地区主要野生动物携带病毒谱调查研究 一种新型微生物群体感应通讯机制与病原防控基础研究 整合多维高通量测序网络数据挖掘端粒维持机制的相关基因 间歇性低压低氧的长效抗抑郁作用和机制研究 植物逆境胁迫的代谢应答机制及其在经济作物高质化的应用研究 脊椎动物抗病毒机制的起源与演化研究 基于zein胶体颗粒稳定Pickering乳液结构单元的有序组装构建功能性膜材料及构效关系研究 离子通道对神经元兴奋-转录偶联的调控机制及其在自闭症中的作用 水稻高产优质新基因的挖掘与应用探讨 信号转导及疾病相关重要蛋白复合体的结构与功能研究 沈永义 邓音乐 熊远妍 曹雄 杨子银 元少春 尹寿伟 李勃兴 王少奎 刘伟 汕头大学 华南农业大学 中山大学 南方医科大学 中国科学院华南植物园 中山大学 华南理工大学 南方医科大学 华南农业大学 深圳北京大学香港科技大 学医学中心 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年 杰出青年
棉花蔗糖合成酶3基因第1个内含子在基因表达中的作用

入到 G S基 因编码 区中构建载体 s U
:1 1 将载体 S sg:11通过农杆菌介导转 入拟南芥 。对转基 因拟 :2 , u i f :2
南芥进行 G S活性分析 , U 发现该内含子对 G S基 因在拟南芥 中的表达具有 负调控作用 , U 并且 特异抑制报告基
因在花粉 中表达 。其结果显示 , 棉花 S S 因编码区的第 1 内含子对基 因的表达具有一定的调控作用。 U 3基 个 关键词 : 棉花 ; U 3 S S ;内含子 ; 因表 达调控 基
为对 照 。
14 GU . S活性 分析
G S组 织化 学染 色 及荧 光定 量 分 析 按 照 Jf r n等 _ U ees f o 1 的方 法进 行 。将 材 料 浸 泡
在染 色液 (0m o ・ 磷 酸 钠 缓 冲溶 液 p 7 0,. o ・ K [ e C ,. o L K [ e 5 m l L H . 0 1mm l L F ( N)] 0 1mm l・ F
S S 基因的 m N U3 R A丰度远远高于无毛突变体的胚珠 J 。通过 R A干扰实验抑制 S S N U 3的转录后 , 出现
了破 坏棉 花纤 维 细胞起 始 、 抑制 纤维 伸 长和影 响 种子发 育 , 这几 个对 棉花 生长 发育 不利 的性状 』 。
因为决定细胞命运 的基因功能与基因的表达模式密切相关 , 以基 因的表达调控显得尤为重要。研 所
杂交 时 , 发现其 只 在棉 花胚 珠表 皮层 表达 , mR A 的丰 度 非 常高 , 些 特 点说 明 S S 且 N 这 U 3基 因 与棉 纤 维细 胞 的分 化 有非 常密 切 的关 系 J 。将 一个 无 毛突 变体 与野 生型 的棉 花 进行 对 比分析 发 现 , 野生 型 的胚珠 中
ORCA3转录因子对红豆杉细胞中紫杉醇生物合成的影响

收稿日期:2012-04-23;修回日期:2012-05-13基金项目:国家自然科学基金青年基金项目(200906036);国家自然科学基金面上项目(20776058)作者简介:于小青(1982-),女,山东潍坊人,硕士研究生。
研究方向:植物次生代谢与调控。
△该作者目前就读于华中科技大学生命科学与技术学院,系2009级本科生。
*通讯作者:付春华。
E-mail:fuch2003@126.com ·研究报告·ORCA3转录因子对红豆杉细胞中紫杉醇生物合成的影响*于小青1,杨海燕△,张蒙1,栗茂腾1,2,付春华1,2*,余龙江1,2(1.华中科技大学生命科学与技术学院,资源生物学与生物技术研究所,湖北武汉430074;2.华中科技大学分子生物物理教育部重点实验室,湖北武汉430074)摘要:ORCA3是一个受MeJA诱导的调控植物基础和次生代谢的转录调控因子。
本文拟探讨ORCA3转录因子对红豆杉细胞中紫杉醇生物合成的影响,旨在为创制紫杉醇稳定高产的红豆杉细胞系,解决细胞大规模培养中紫杉醇含量低的瓶颈问题提供参考。
从长春花中克隆ORCA3基因后,采用农杆菌介导法转入红豆杉细胞中,通过PCR、组织化学活性染色等检测法,筛选得到20余株转基因细胞系。
HPLC检测转基因细胞系内紫杉醇含量表明:与阴性对照相比,转基因细胞系的紫杉醇含量增加了0.5 2.5倍,最高可达159.315μg/g。
因此,ORCA3转录因子可以调控红豆杉细胞中紫杉醇合成。
关键词:红豆杉;遗传转化;ORCA3;紫杉醇中图分类号:Q789文献标识码:A文章编号:1008-0457(2012)03-0189-05紫杉醇(Taxol)是一种重要的抗肿瘤药物,由于它在红豆杉植物中含量极低(约为0.01%),市场供求矛盾突出[1]。
目前,紫杉醇主要靠天然提取和化学半合成法获得,这两种方式均需要依赖有限的红豆杉资源。
因此,细胞大规模培养方法被认为是生产紫杉醇最有希望的替代途径之一[2]。
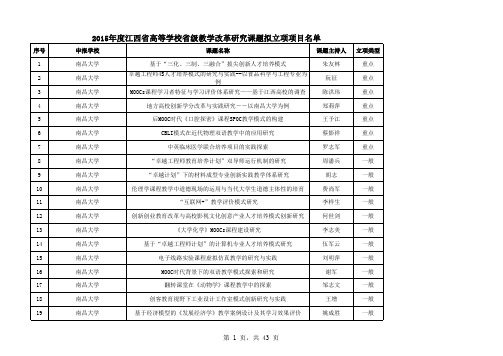
2015年江西省高等学校教学改革立项
化志章 王昌晶 刘清华 黄小勇 郑蝉金 罗海梅 程晏蓓 熊小勇 杨丽华 叶继华 黄福生 聂晶 刘忠民 蒋梅鑫 柳敏 刘碧珍 曾广洪 汤舒俊 盛寿日
重点 重点 重点 重点 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般
第 4 页,共 43 页
82 83 84 85 86 87 88 89 90 91 92 93 94 95 96
41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61
南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学
段卫东 吴伟伟 贾书磊 魏丹 袁新春 李春泉 黄伟 陈加祥 李小鹏 时影 汪艳璐 刘捷 王秋伟 sujunhua 刘敏 廖一静 孙翔 蔡丹 王振 徐晓芳 李静
一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般 一般
第 2 页,共 43 页
2015年度江西省高等学校省级教学改革研究课题拟立项项目名单
序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 申报学校 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 南昌大学 课题名称 基于“三化、三制、三融合”拔尖创新人才培养模式 卓越工程师4S人才培养模式的研究与实践--以食品科学与工程专业为 例 MOOCs课程学习者特征与学习评价体系研究——基于江西高校的调查 地方高校创新学分改革与实践研究――以南昌大学为例 后MOOC时代《口腔探密》课程SPOC教学模式的构建 CBLI模式在近代物理双语教学中的应用研究 中英临床医学联合培养项目的实践探索 “卓越工程师教育培养计划”双导师运行机制的研究 “卓越计划”下的材料成型专业创新实践教学体系研究 伦理学课程教学中道德现场的运用与当代大学生道德主体性的培育 “互联网+”教学评价模式研究 创新创业教育改革与高校影视文化创意产业人才培养模式创新研究 《大学化学》MOOCs课程建设研究 基于“卓越工程师计划”的计算机专业人才培养模式研究 电子线路实验课程虚拟仿真教学的研究与实践 MOOC时代背景下的双语教学模式探索和研究 翻转课堂在《动物学》课程教学中的探索 创客教育视野下工业设计工作室模式创新研究与实践 基于经济模型的《发展经济学》教学案例设计及其学习效果评价 课题主持人 朱友林 阮征 陈洪玮 郑莉萍 王予江 蔡影祥 罗志军 周潘兵 胡志 费尚军 李样生 何世剑 李志美 伍军云 刘明萍 谢军 邹志文 王增 姚成胜 立项类型 重点 重点 重点 重点 重点 重点 重点 一般 一般 一般 一般 一般 一般 一般 一般 一般 组课程报告为基础的《医学遗传学》考试模式改革的研究
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
提出了改进的三链 DNA 模型求解 0-1 规划的设计。该方法编码 n 个变量的每种组合的所有排列情况。 此编码
n 并使对可行解的筛选更加有效。 利用寡聚脱氧 方式不仅使计算所需有效分子量从 O( ( 2 n) ! ) 下降到 O( 2 n! ) ,
核苷酸( ODN) 在 RecA 蛋白介导下与同源的双链 DNA 匹配成三螺旋 DNA 的特点, 可推广到更多以双链 DNA 分 子为计算模型的解的检测中。 关键词: 三链 DNA; 0-1 整数规划; DNA 计算 中图分类号: TP301. 6 文献标志码: A 文章编号: 1001-3695 ( 2013 ) 01-0056-04 doi: 10. 3969 / j. issn. 1001-3695. 2013. 01. 013
Improvement of 0-1 integer programming based on triplestranded DNA structure
REN Xiaoling,BAI Xue,LIU Xiyu
( School of Management Science & Engineering,Shandong Normal University,Jinan 250014 ,China)
0
引言
1957 年, Feisenfeld 等人提出了三链核酸的概念 。 由于双
链 DNA 中两条相互缠绕且方向相反的链是依靠互补碱基对之 间的氢键相连, 碱基对又偏向螺旋轴一侧, 所以在两条链之间 形成了大小两沟。三链 DNA 正是在双链 DNA 的基础上, 通过 含有多聚嘌呤的那条链, 以 Hoogsteen 和反式 Hoogsteen 型氢键 且第三条链位于双链 与新 加 入 的 第 三 条 链 相 连 接 形 成 的, DNA 的大沟中, 主要有嘧啶—嘌呤—嘧啶和嘌呤—嘌呤—嘧 啶两种类型。 2004 年,Shigemori 等人发现寡聚脱氧核苷酸能在 RecA 蛋白及 ATP γ S 的存在下与双链 DNA 形成稳定的三链结构。由 于三链 DNA 在形成过程中不会发生错配和形成发夹结构, 而 且反应后得到的解为双链形式, 比较稳定, 不会像单链 DNA 因 为过长而形成发夹结构或断裂 。 因此, 基于三链结构 DNA 计 算的错误率和编码复杂度会降低 。 目前, 关于三链 DNA 的研究侧重于生物、 医学领域, 利用
第 30 卷第 1 期 2013 年 1 月
计 算 机 应 用 研 究 Application Research of Compun. 2013
* 基于三链 DNA 结构的 0-1 整数规划改进研究
任晓玲,白
摘
雪,刘希玉
( 山东师范大学 管理科学与工程学院,济南 250014 ) 要: 为实现 DNA 计算中对解的有效筛选, 防止探针与探针之间的错配、 发夹结构等, 以及便于检测最终解,
Abstract: To realize the effective screening of solutions in DNA computing and to prevent the mismatch between probes,the formation of the hairpin structure etc, this paper presented an improved triplestranded DNA model to solve 0-1 integer programming problem. This method made a full array of all combinations of n 0-1 variables. Compared with the original one,the amount of DNA strands needed dropped from O( ( 2 n) ! ) to O( 2 n n! ) and the improved one made a better selection of feasible solutions. Triplehelix structure could be constructed by the homologous doublehelix DNA with the oligodeoxyribonucleotides ( ODN) in the mediated of RecA protein. It could make use of its special structure to promote it to the selection of solutions with the doublehelix computing model. Key words: triplestranded DNA; 0-1 integer programming; DNA computing
收稿日期: 2012-05-22 ; 修回日期: 2012-06-25 J12LN65 ) 项目( J12LN22 ,
三链 DNA 结构实现 DNA 计算的实例不多, 主要解决了 0-1 整 [1 , 2 ] [3 , 4 ] [5 ] 、 、 数规划问题 可满足性问题 顶点着色问题 、 邮递员 [6 ] 问题以及工序安排问题 。2008 年, 杨静等人利用同源的存 在抗原蛋白质的脱氧核苷酸( ODN ) 在双螺旋 DNA 链中形成 的三链 DNA 结构解决了简单的 0-1 整数规划问题, 通过对三 3, 4]在阐 链 DNA 的构造和筛选完成了可行解的获取 。 文献[ 述三链 DNA 的形成过程以及可行性实验研究的基础上, 提出 将三链 DNA 计算模型应用于求解可满足性问题 。将 n 个变量 ODN 复合体探针, 的不同取值对应为 2 n 种不同的 RecA 蛋白与初始解空间在适当的反应液中哺育, 通过探针是否与库链形 成稳定的三链 DNA 结构判断并筛选满足符合每个子句的解序 列。在求解图顶点着色问题时, 利用探针与库链中某些 DNA 双链形成的三链 DNA 结构过滤掉相邻节点着同一颜色的非可 6]利 行解, 进而筛选出相邻节点着不同颜色的可行解 。 文献[ 用类似的方案求解了供需安排问题 。