3-RNA转录后加工-
RNA转录与转录后加工

在大多数真核生物中,RNA聚合酶在转录终止后,会在3’端加上一段多聚腺苷酸尾巴。这个过程称为加尾。加 尾的主要作用是促进RNA从核内向细胞质转运,并保护RNA免受3’核酸外切酶的降解。此外,多聚腺苷酸尾巴 也是一些RNA结合蛋白的识别位点,参与mRNA的稳定性、定位和翻译调控。
剪接
总结词
剪接是指将转录的RNA中的内含子序列 去除,并将外显子序列连接起来的加工 过程。
详细描述
C-to-U编辑由胞嘧啶脱氨酶催化,将RNA 中的胞嘧啶转变为尿嘧啶,导致RNA序列 发生变化。这种编辑可以影响RNA的翻译 和功能。
其他编辑类型
总结词
除了A-to-I和C-to-U编辑外,还存在其他类型的RNA编辑,如C-to-A编辑、C-to-G编 辑等。
详细描述
这些编辑类型在特定的生物或组织中发生,由不同的酶催化,导致RNA序列发生不同 的变化。这些编辑可以影响RNA的稳定性、翻译和功能。
肽链终止
终止密码子出现时,核糖体 释放合成的多肽链,并回收 mRNA。
蛋白质合成的起始
起始氨基酸的识别
起始密码子(AUG)被识别并结合甲酰蛋氨酸,形成甲酰蛋氨酸-tRNA。
甲酰蛋氨酸-tRNA在核糖体上的定位
甲酰蛋氨酸-tRNA与起始因子结合,定位到核糖体的P位点。
起始复合物的形成
甲酰蛋氨酸-tRNA与mRNA结合,形成起始复合物。
02
翻译水平调控
03
细胞内环境调控
翻译过程中蛋白质的表达水平可 以影响RNA的稳定性。
细胞内的pH值、离子浓度等环境 因素也可以影响RNA的稳定性。
05
RNA的翻译和蛋白质合成
mRNA的翻译
翻译起始
mRNA在核糖体上定位并结 合翻译起始因子,形成起始 复合物。
简述rna转录后加工过程

简述rna转录后加工过程摘要:1.RNA转录后加工过程的概述2.RNA转录后加工的主要步骤a.剪接b.剪切c.RNA编辑d.RNA降解3.各步骤的功能和意义4.实例分析5.RNA转录后加工在生物体中的作用6.研究RNA转录后加工的意义和前景正文:在我们生物体内,基因通过转录过程将DNA信息转化为RNA,但这只是RNA生命历程中的第一步。
接下来,RNA要经历一系列复杂的加工过程,才能最终发挥其生物学功能。
这个过程被称为RNA转录后加工。
RNA转录后加工的主要步骤包括剪接、剪切、RNA编辑和RNA降解。
剪接是指将RNA前体分子中的内含子去除,并将外显子连接成成熟的RNA分子。
这一过程通过特定的酶家族,如剪接酶,来实现。
剪切是指在RNA分子的3"端添加poly(A)尾巴,这是几乎所有真核生物RNA的共同特征。
RNA编辑则是指在RNA分子上发生碱基改变,这一过程依赖于特定的编辑酶和相应的底物。
最后,RNA降解是指RNA分子在细胞内的分解过程,这对于调控RNA水平和维持细胞内稳态至关重要。
这些加工过程对于RNA最终的生物学功能具有重要意义。
以剪接为例,它能消除RNA前体中无功能的RNA片段,使成熟的RNA更具特异性和高效性。
同时,RNA编辑能够改变RNA的序列,从而影响其翻译效率和稳定性。
在生物体中,RNA转录后加工涉及多种生物过程,如基因表达调控、病毒复制和免疫反应等。
对RNA转录后加工的研究,有助于我们深入了解生命过程中的基因表达调控机制,为治疗疾病和开发新型药物提供理论依据。
随着生物科学技术的不断发展,对RNA转录后加工的研究将越来越深入。
第8章 转录后加工

4、拼接(splicing)
Ø 大多数的真核生物基因是断裂基因;
Ø 其中编码序列称为外显子(exon),外显子之间的 介入序列称为内含子(intron);
Ø 少数真核生物基因(如组蛋白、干扰素)是连续的;
Ø 高等真核生物的基因中多数内含子比外显子长得多, 而低等真核生物(如酵母)的基因中内含子比较短而 且少见;
Ø 有些生物的rRNA前体含有内含子,需要拼接;
p.205
Ø 哺乳动物的18S, 28S, 5.8S rRNA gene 组成一个 转录单位,由RNA pol I 转录产生45S的前体分子;
Ø 5S rRNA gene 与不转录区域组成转录单位, 由RNA pol III转录;
small nucleolar RNA(snoRNA)
Ø 高度精确; Ø 依赖于多种顺式作用元件和反式作用因子; Ø 共转录事件;
顺式元件1
Ø 内含子具有一致的保守序列,即5’拼接点为 GU,3’拼接点为AG,称为BreathnathChambon规则,也称GU/GT-AG规则。
顺式元件2
为什么只有mRNA被加帽?
Ø 只有RNA聚合酶 II 合成的转录产物(mRNA、 部分snRNA)才有帽子结构;
Ø 因为加帽酶只能与RNA聚合酶 II 的CTD结构 域结合;而CTD是RNA聚合酶II 特有的。
Ø 加帽酶与CTD的磷酸化形式(延伸型)结合。 Ø 转录产物一旦从RNA聚合酶II中显露出来,就
可以与加帽酶接触。
2、3’端加尾
Ø 真核生物的大多数mRNA及其前体在3’端有约 250 nt 的连续的AMP。 Ø poly(A) 由poly(A) polymerase(PAP)添加; Ø mRNA进入细胞质后,其poly(A)可以被更新 : 不断地被RNase降解,再由细胞质中的PAP重新 合成。
分子生物学-07-3-生物信息的传递-3转录后加工-小RNA-RNA拼接1
能够识别发夹 结构的内切酶
1
RNAase P 3
2 RNA酶D
4 4
4
4
2020/10/28
多种酶的处理
2020/10/28
23
tRNA的内含子去除
2020/10/28
2020/10/28
22
rRNA的转录后处理
1、原核生物和真核生物是较为相似的,分别由多种
rRNA组成它的核糖体,包括5S rRNA 、5.8S S rRNA、 18 S rRNA、 28 S rRNA
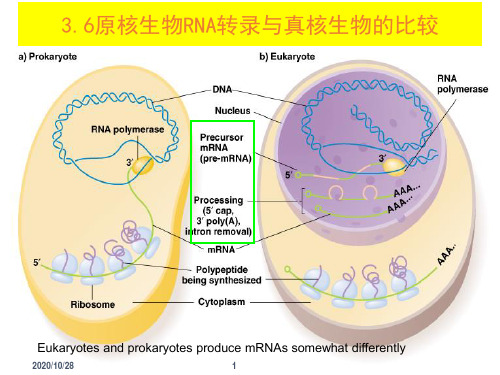
3.6原核生物RNA转录与真核生物的比较
Eukaryotes and prokaryotes produce mRNAs somewhat differently
2020/10/28
2020/10/28
1
3.6.1 原核生物mRNA的特征
1. 多顺反子,有共同的起始子和终止信号 2. 转录和翻译的时空性 3. 5′端无帽子结构, 3′端没有或只有较短的poly(A)结构 4. 起始密码子常为AUG,有时也为GUG,甚至UUG 5. 半衰期短
14
Poly(A)尾巴的功能:
(1)尾巴可能与核-质转运有关;但是无尾巴的 mRNA,如组蛋白的mRNA同样可以通过核膜 进入细胞质
(2)尾巴的长短对mRNA的翻译也无影响,因 此其功能无定论。
(3)细胞质中mRNA 的Poly(A)尾巴与蛋白质相 结合构成mRNP(一种蛋白质分子)。其功能 不清楚。
2020/10/28
帽子结构功能:
1.能被核糖体小亚基识别,促使mRNA和 核糖体的结合;
2.m7Gppp结构能有效地封闭mRNA 5’末 端,以保护mRNA免受5’核酸外切酶的降解 ,增强mRNA的稳定。
rna转录后加工方式

rna转录后加工方式
RNA转录后加工(RNA post-transcriptional processing)是指在RNA分子合成之后,在细胞中对其进行修饰和修剪的过程。
这些加工方式可以使原始RNA分子成熟,并使其具有功能性。
以下是几种常见的RNA转录后加工方式:
剪接(Splicing):在真核生物中,基因的转录产物(前体mRNA)经过剪接过程,去除其中的内含子(intron),保留外显子(exon),从而形成成熟的mRNA分子。
剪接是通过剪接体(spliceosome)来完成的,其中包括snRNPs等辅助因子。
5'端修饰:RNA的5'端通常经过加上7-甲基鸟苷(7-methylguanosine)和三磷酸核苷酸链(PPP 链)的修饰,形成5'甲基鸟苷帽(5' cap)。
这个帽子在RNA稳定性、转运和翻译起重要作用。
3'端修饰:RNA的3'端通常经过加上聚腺苷酸(polyadenylation)的修饰。
这个poly(A)尾巴有助于RNA的稳定性、转运和翻译,并参与转录终止的过程。
RNA编辑:在一些生物体中,RNA的序列可以通过RNA编辑(RNA editing)进行改变。
这种编辑通常涉及碱基的替换、插入或删除,从而改变RNA的编码能力和功能。
RNA修饰:RNA分子可能会经历各种修饰,如甲基化、脱氨基、糖基化等。
这些修饰可以增强RNA的稳定性、调节翻译和识别,以及影响RNA的功能。
RNA转录后加工是一个复杂而精确的过程,它可以使原始的转录产物转化为功能性的RNA 分子。
这些加工方式对于基因表达调控和细胞功能起着重要的作用。
RNA转录后的加工

二、真核生物RNA修饰加工的主要方式:
pre-RNA
capping tailing splicing methylation editing
mature RNA
生物学意义;
l interrupted gene (interrupted RNA)
move introns as template (stop codon) (protein translation)
Man β- globin mRNA 5’-------UGCCUAAUAAA------------poly(A) 3’ -20
2、mRNA 3’端的加尾时间:
➢转录过程中暴露 出AAUAAA信号 后,核酸酶在该信 号下游约11-30个 碱基处进行切割。
➢polyA的长度一 般是50-200个碱 基左右。
•外显子较短(100~200bp),内含子较长(1 kb)。
Hale Waihona Puke ➢剪接(RNA splicing):内含子的去除和外显 子的连接过程就称为剪接或称为RNA 剪接。
➢不均一核RNA(heterogeneous nuclear, hnRNA) :mRNA 的初始转录产物比成熟的 mRNA平均长度长,非常不稳定,序列的复 杂程度也非常高,称为不均一核RNA。
Rabbit α- globin mRNA 5’-------CUUUGAAUAAA-----------poly(A) 3’ -20
Rabbit β- globin mRNA 5’-------UGGCUAAUAAA-----------poly(A) 3’ -20
Man α- globin mRNA 5’-------CUUUGAAUAAA------------poly(A) 3’ -20
RNA转录和加工

套索结构的发现使人们认识到, 套索结构的发现使人们认识到,内含子的剪接是通过 两次转酯反应完成的。在第一次转酯反应中, 两次转酯反应完成的。在第一次转酯反应中,分支位 进攻5 剪接位点, 点A的2’-OH进攻5’剪接位点,使其断裂,同时这个A -OH进攻 剪接位点 使其断裂,同时这个A 与内含子的第一个核苷酸( 形成2 与内含子的第一个核苷酸(G)形成2’ , 5’ -磷酸 二酯键,内含子自身成环,形成套索结构。 剪接位 二酯键,内含子自身成环,形成套索结构。3’剪接位 点的断裂依赖于第二次转酯反应。上游外显子的3 - 点的断裂依赖于第二次转酯反应。上游外显子的3’- OH末端攻击3 剪接位点的磷酸二酯键 促使其断裂, OH末端攻击3’剪接位点的磷酸二酯键,促使其断裂, 末端攻击 剪接位点的磷酸二酯键, 使上游外显子的5 -0H和下游外显子的 - 和下游外显子的5 使上游外显子的5’-0H和下游外显子的5’-磷酸基团 连接,并释放出内含子,完成剪接过程。 连接,并释放出内含子,完成剪接过程。被切除的内 含子随后变成线性DNA 随即被降解。 DNA, 含子随后变成线性DNA,随即被降解。
通过分析体外剪接反应中形成的中间体, 通过分析体外剪接反应中形成的中间体,发现内含子 是以一种套索结构( 是以一种套索结构(lariat structure )的形式被切除 即内含子5 端的鸟苷酸依靠 , - 端的鸟苷酸依靠2 的,即内含子5’端的鸟苷酸依靠2’,5’-磷酸二酯键与 靠近内含子3 末端的一个腺苷酸连接在一起 末端的一个腺苷酸连接在一起。 靠近内含子3’末端的一个腺苷酸连接在一起。该腺苷 酸被称作分支位点 分支位点, 酸被称作分支位点,因为在套索结构中它形成了一个 RNA分支 分支。 RNA分支。
在内含子的剪接过程中, 在内含子的剪接过程中,剪接装置必须识别正确的 剪接位点,以保证外显子在剪接的过程中不被丢失, 剪接位点,以保证外显子在剪接的过程中不被丢失, 同时荫蔽的剪接位点要被忽略。 同时荫蔽的剪接位点要被忽略。所谓隐蔽剪接位点 (cryptic splice site )是指与真正的剪接位点 相似的序列。已经知道一类被称为SR蛋白( 相似的序列。已经知道一类被称为SR蛋白(SR SR蛋白 protein)的剪接因子在剪接位点的选择中发挥重要 protein) 作用。 作用。
RNA转录后的剪切与加工

目录
• rna转录后的剪切 • rna加工 • rna剪切与加工的相互关系 • rna剪切与加工的异常表达与疾病
01
rna转录后的剪切
剪切的定义与重要性
剪切的定义
RNA转录后,通过特定的核酸酶将 RNA分子从转录起始位点至终止位点 之间的序列进行切割的过程。
剪切的重要性
剪切的过程
在剪切过程中,核酸酶首先识别RNA分子中的特定位点,然后进行切割,产生两 个新的RNA分子片段。这些片段可能进一步被加工或降解。
剪切的调控机制
剪切的调控机制包括多种因素,如基因的启动子、增强子、沉默子和miRNA等。这些因素可以影响 RNA聚合酶的活性,从而影响转录的起始和终止,进一步影响剪切过程。
高效、更灵敏的技术用于研究这些过程。
04
rna剪切与加工的异常表 达与疾病
剪切与加工异常的表达模式
异常剪切
在某些情况下,RNA剪切过程可能发生异常,导致产生异常的RNA剪切产物。这些异常的剪切产物可能导致基因 表达的异常,进一步影响细胞功能。
异常加工
RNA加工过程中,如甲基化、磷酸化等修饰过程发生异常,也可能导致RNA的功能异常。这些异常的RNA可能 无法正确地指导蛋白质的合成,或者可能产生有毒性的RNA。
剪切与加工异常与疾病的关系
遗传性疾病
一些遗传性疾病的发生与RNA剪切与加工的异常有关。例 如,一些遗传性神经性疾病可能与特定基因的异常剪切有 关。
癌症
癌症的发生也常常伴随着RNA剪切与加工的异常。一些癌 症可能由于特定基因的异常剪切或加工而导致其表达水平 的上调或下调。
感染性疾病
某些感染性疾病也可能影响RNA的剪切与加工。例如,某 些病毒可能通过干扰宿主细胞的RNA剪切与加工过程来影 响基因表达,从而促进病毒的复制。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
●
●
●
●
●Байду номын сангаас
●
●
●
●
●
●
●
●
●
●
●
●
●
●
GT—AG法则: 序列分析表明,几乎每个内含子5’端起 始的两个碱基都是GT,而3’端最后两个碱基 总是AG,由于这两个碱基的高度保守性和广 泛性,有人把它称为GT—AG法则,即: 5’GT......AG 3’。 剪接信号
12
P102
● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ●
8
hnRNP
● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ●
RNA剪接真 正识别的是 hnRNP,而 不是前体 mRNA
9
1. RNA序列决定了剪接的发生位点
● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ●
比较不同基因的核苷酸序列发现, mRNA前 体中内含子的两端边界存在共同的序列,这 些序列结构可能是产生 mRNA前体剪接的 信号。
● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ●
RNA的再编码:mRNA可以改变原来的 编码信息,在某些情况下不是以固定的 方式被翻译,科学上把RNA的编码和读 码方式的改变
41
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
方式:mRNA的读码信号在tRNA、 rRNA、和其他蛋白因子作用下发生+1/1移码; 核糖体跳跃50个核苷酸; 终止子通读
30
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
31
总结
● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ●
32
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
Ⅰ类和Ⅱ类内含子及细胞核RNA这三类内含子的剪 接都是通过两次连续的转酯反应进行的。在第一次 转酯反应中,由一个游离羟基(在细胞核基因内含 子和Ⅱ类内含子中由内部2’羟基提供,Ⅰ类内含子 中由一个游离的G提供)发动对5’外显子-内含子连 接点的攻击。 第二次转酯反应中,已经释放的外显子末端的游离 3’羟基接着攻击3’内含子-外显子的连接点。在反应 中,磷酸二酯键的数目是保守的,所以反应不需要 额外能量,这是剪接进化的一个重要特征。
15
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
16
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
在剪接反应前后,磷酸二酯键的数目是不变 的,只是由原来的外显子-内含子之间的两个 5-3 磷酸二酯键变为两个外显子之间的5 -3 和内含子本身的套索结构中的5 -2 磷酸二 酯键。 转酯反应(transesterification) 酯键从一个位臵转移到另一个位臵。
23
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
24
5. I 类和II类内含子
● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ●
根据内含子的结构特点及其剪接机制把他们划 分为: I类内含子 II类内含子 细胞核RNA(GU-AG, AU-AC)
25
●
●
●
●
●
●
●
●
●
●
●
●
●
●
42
3.8.3. RNA的化学修饰
● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ●
只含有70-100个核苷酸的核仁RNA(snoRNA) 参与RNA的化学修饰 这些RNA通过碱基配对的方式,把rRNA分 子上需要修饰的位点找出来。
●
●
●
●
●
低等真核生物、细菌-Ⅰ类内含子 细胞器(线粒体,叶绿体)-Ⅱ类内含子。
26
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
Ⅰ类和Ⅱ类内含子是根据它们的内部组成分 类的。每种内含子都可以折叠成典型的二级 结构。 Ⅰ类和Ⅱ类内含子含有RNA自我切割的能力, 称为自我剪接。
27
Ⅰ类内含子
● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ●
10
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
内含子两端序列不能互补; 连接区序列高度保守, GT-AG法则。序列分析表明, 几乎每个内含子5’端起始的两个碱基都是GT,而3’ 端最后两个碱基总是AG,由于这两个碱基的高度保 守性和广泛性,有人把它称为GT—AG法则,即: 5’GT.....AG 3’。 按照内含子方向,左边的称为5剪接位点(5 splice site,又叫供体位点(donor site),右边的称为3剪接位 点(3 splice site) 又叫受体位点(acceptor site).
17
3. RNA剪接大多发生在剪接体上
● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ●
18
●
●
细胞核中的小分子RNA称为细胞核小RNA (small nuclear RNA, snRNA) 细胞质中的小分子RNA称为细胞质小RNA (small cytoplasmic RNA, scRNA) 在自然状态下,它们以核糖核蛋白颗粒 (snRNP 和 scRNP)形式存在,俗称snurps 和 scyrps 在核仁中也存在着一类小的RNA,称为核仁 小RNA (small nucleolar RNA, sno RNA),它 们在核糖体RNA的加工中起作用。
●
●
●
●
●
●
●
●
●
●
●
●
由UACUAAC的保守A通过2 羟基对5剪接位点发动 亲核攻击,内含子5 端切开,内含子5 端的保守G的 5位与分支点保守的A的2位形成磷酸二酯键(5 -2 磷酸二酯键)而产生套索结构。
第一步中释放的外显子的游离3羟基攻击3剪接位点中 的键。内含子3 剪接点被切断,套索结构释放,同时, 外显子连在一起。 内含子的套索结构被切开,形成线状并很快被释放。
游离的鸟苷或鸟苷酸介导, 鸟苷或鸟苷酸的3 -OH 作 为亲核基团攻击内含子5 端的磷酸二酯键,从上游 切开RNA链。(第一个转 酯反应) 上游外显子的自由3 -OH 作为亲核基团攻击内含子3 位核苷酸上的磷酸二酯键, 使内含子被完全切开,上 下游两个外显子通过新的 磷酸二酯键连接。 (第二 个转酯反应)
28
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
29
Ⅱ类内含子
● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ●
内含子本身的靠近3端的腺苷酸2 -OH作为 亲核基团攻击内含子5端的磷酸二酯键,从上 游切开RNA链后形成套索状结构。(第一个 转酯反应) 上游外显子的自由3 -OH作为亲核基团攻击 内含子3 位核苷酸上的磷酸二酯键,使内含 子被完全切开,上下游两个外显子通过新的 磷酸二酯键连接。 (第二个转酯反应)
1. 长度和序列没有共 同性,一般16-46 个核苷酸 2. 位于反密码子下游 3. 内含子和外显子间 的边界没有保守序 列
3
真核生物tRNA的加工
● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ● ●
内含子的剪接 3‘端添加CCA 核苷酸修饰。
4
3.7.3 真核生物rRNA的转录后加工
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
19
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
snRNP:ribonucleo protein particle(许多相 对分子质量较小的核内RNA(snRNA, small nuclear RNA)以及与这些RNA结合的核蛋白, 参与mRNA的剪接。 参与剪接的snRNP包括U1,U2,U5,U4, U6,他们根据所含有的sn RNA而命名,每个 snRNP 含有一个snRNA 和几种蛋白质。
35
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●