橡胶树的基因组学研究
橡胶树种质资源的倍性鉴定

橡胶树种质资源的倍性鉴定橡胶树是一种重要的经济作物,广泛栽种于亚热带和热带地区。
与其他植物一样,橡胶树存在着自然杂交、人工杂交等多种途径形成的多倍体现象,而倍性对植物的生长发育、生殖过程、细胞特性等均有重要影响。
因此,橡胶树种质资源的倍性鉴定十分关键。
倍性鉴定是指对植物的染色体组进行分析,确定其植物体细胞包含的染色体组数。
通常采用细胞学方法,如根尖染色体观察、荧光原位杂交等技术,也可以利用分子生物学方法,如荧光PCR、流式细胞术等技术。
在橡胶树种质资源的倍性鉴定中,常常用到的是荧光原位杂交技术和荧光PCR技术。
荧光原位杂交技术是一种基于DNA同源性的高级细胞生物学技术,可以将目标DNA序列与荧光素染色剂结合,经过融合反应,产生出发光信号。
在橡胶树的倍性鉴定中,可用较短的DNA序列或人造引物作为探针,将其连接至荧光素染色剂,结合至橡胶树细胞中的目标基因或染色体上,通过荧光显微镜观察发光程度,从而确定细胞核中染色体数目。
荧光PCR技术是一种PCR技术的改良版,其在PCR反应中加入荧光素酶和荧光探针,做到增加PCR产物的同时,即时检测反应结果的同时还可以确定荧光标记的目标物的基因信息和数量。
在橡胶树倍性鉴定中,荧光PCR技术通常选择利用基因多样性差异较大的引物组合,通过PCR扩增橡胶树DNA片段,利用荧光标记旁的引物缩短片段,从而对产物分析,确定细胞核染色体数目。
橡胶树倍性鉴定中应当注意的问题1、样本选择:橡胶树种间间存在着明显的品种差异和种间杂交现象,对于不同品种或外来种而言,倍性情况往往存在较大的差异,因此,选择的样本要有代表性和典型性,同时要注意避免杂交现象所带来的干扰。
2、适当调整染色体观察区域:在染色体观察中,要特别注意细胞取材的大小、染色体的展开程度等,避免单细胞或碎片状的观察样本,检验过程中必须着重保障对橡胶树核型分析的准确性。
3、技术选择:不同的技术具备不同的优劣,应根据实际需求和技术难度合理选择。
橡胶草apx基因家族的全基因组鉴定及表达分析

西北植物学报,2019,39(11):1935-1942A c t aB o t .B o r e a l .GO c c i d e n t .S i n.㊀㊀d o i :10.7606/j .i s s n .1000G4025.2019.11.1935㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀h t t p ://x b z w x b .a l l jo u r n a l .n e t 收稿日期:2019G07G18;修改稿收到日期:2019G10G15基金项目:海南省自然科学基金创新研究团队项目(2018C X T D 341);国家自然科学基金面上项目(31570301)作者简介:高㊀璇(1995-),女,在读硕士研究生,主要从事橡胶草基因功能分析和生物技术研究.E Gm a i l :1577384808@q q.c o m ∗通信作者:张雪妍,副研究员,硕士生导师,主要从事天然橡胶生物合成调控机制研究.E Gm a i l :z h a n g x u e ya n _c a a s @126.c o m 橡胶草A P X 基因家族的全基因组鉴定及表达分析高㊀璇1,2,刘进平1,于㊀莉2,袁博轩2,王旭初2,张雪妍2∗(1海南大学热带作物学院,海口570228;2海南师范大学生命科学学院,海口571158)摘㊀要:该研究利用生物信息学方法,在橡胶草(T a r a x a c u mk o k Gs a g h y z )全基因组中鉴定出7个T k A P X s 基因家族成员,进一步分析发现7个成员中有1对是复制基因(T k A P X 4/T k A P X 6).进化分析结果显示,7个基因可分为4个亚组;染色体定位分析表明,T k A P X 基因家族成员分布广泛,7个T k A P X s 基因位于7条不同的染色体上;亚细胞定位结果表明,T k A P X 1㊁T k A P X 3㊁T k A P X 5和T k A P X 7定位于细胞质,T k A P X 4和T k A P X 6定位于质膜,T k A P X 2定位于叶绿体.启动子区域顺式作用元件分析结果显示,橡胶草A P X 基因含有大量的应激反应元件,推测A P X 基因可能对各种外界刺激和胁迫存在灵敏的应激反应机制.实时荧光定量P C R 分析表明,橡胶草7个T k A P X s 基因家族成员在花萼㊁花瓣㊁花梗中表达量均较低,大多数T k A P X s 在根茎叶中的表达水平大于花及胶乳,其中T k A P X 1在根和茎中的表达水平最高,推测T k A P X 1基因可能在橡胶草生长发育过程中起重要作用.进一步对T k A P X s 基因家族在逆境胁迫(冷㊁热㊁盐㊁旱)和激素(乙烯㊁茉莉酸甲酯)处理下的表达分析显示,T k A P X 3在根及叶中的表达水平均较对照有大幅度升高,推测T k A P X 3基因可能在橡胶草应答逆境胁迫和激素处理反应过程中起重要作用.关键词:橡胶草;抗坏血酸过氧化物酶;系统进化;基因表达分析中图分类号:Q 786;Q 789文献标志码:AG e n o m e Gw i d e I d e n t i f i c a t i o na n dE x p r e s s i o nA n a l y s i s o f A P X G e n eF a m i l yi n T a r a x a c u mk o k Gs a g h y z G A O X u a n 1,2,L I UJ i n p i n g 1,Y U L i 2,Y U A NB o x u a n 2,WA N G X u c h u 2,Z H A N G X u e ya n 2∗(1C o l l e g e o f T r o p i c a l C r o p s ,H a i n a nU n i v e r s i t y ,H a i k o u ,570228,C h i n a ;2C o l l e g e o f L i f e S c i e n c e ,H a i n a nN o r m a l U n i v e r s i t y,H a i k o u ,571158,C h i n a)A b s t r a c t :I n t h i s s t u d y ,7T k A P X s g e n e sw e r e i d e n t i f i e d i n T a r a x a c u mk o k Gs a g h y z w h o l e g e n o m eu s i n g b i o i n f o r m a t i cm e t h o d s .F u r t h e ra n a l y s i sr e v e a l e dt h a t2g e n e s (T k A P X 4/T k A P X 6)w e r er e pl i c a t i o n g e n e s .A c c o r d i n g t ot h er e s u l t so f p h y l o g e n e t i cr e l a t i o n s h i ps ,7T k A P X g e n e s w e r ed i v i d e di n t of o u r g r o u p s .T h e c h r o m o s o m a l l o c a l i z a t i o na n a l y s i s i n d i c a t e d t h a t t h e 7T k A P X g e n e sw e r ew i d e l y d i s t r i b u t e d a n dl o c a t e d o n 7d i f f e r e n t c h r o m o s o m e s .S u b c e l l u l a r l o c a l i z a t i o n r e s u l t s i n d i c a t e d t h a t T k A P X 1,T k A P X 3,T k A P X 5a n d T k A P X 7w e r e l o c a l i z e d i n t h e c y t o pl a s m ,T k A P X 4a n d T k A P X 6w e r e l o c a l i z e d o n t h e p l a s m am e m b r a n e ,a n d T k A P X 2w a s l o c a l i z e d i n t h e c h l o r o p l a s t .T h e a n a l y s i s o f t h e c i s Ga c t i n g el e Gm e n t i n t h e p r o m o t e r r e g i o ns h o w s t h a t t h e r e a r e a l a r g en u m b e r o f s t r e s s r e s po n s e e l e m e n t s i n T k A P X s s e q u e n c e ,s u g g e s t i n g t h a t t h e r em i g h t b e s e n s i t i v e s t r e s s r e s po n s eo f T k A P X s t ov a r i o u s e x t e r n a l s t r e s s .R e a l Gt i m e q u a n t i t a t i v eP C Ra n a l y s i s s h o w e d t h a t t h e e x p r e s s i o n l e v e l s o f 7T k A P X g e n e s f a m i l y me m b e r s i n c a l y x e s ,p e t a l s a n d p e d i c e l sw e r e l o w.T h ee x pr e s s i o n l e v e l o fm o s t T k A P X s i nr o o t sa n d l e a v e sw a s g r e a t e r t h a n t h a t o f f l o w e r s a n d l a t e x .T k A P X 1h a s t h e h i g h e s t e x p r e s s i o n l e v e l i n r o o t s a n d s t e m s ,s u gGg e s t i n g t h a t i tm a y p l a y a n i m p o r t a n t r o l e i n t h e g r o w t h a n d d e v e l o p m e n t o f T.k o kGs a g h y z.F u r t h e r m o r e, w ed e t e c t e dt h ee x p r e s s i o nl e v e lo f T k A P X g e n e sa f t e rs t r e s s(c o l d,h o t,s a l t,d r o u g h t)a n dh o r m o n e (e t h y l e n e,m e t h y l j a s m o n a t e)t r e a t m e n t s.T h e e x p r e s s i o n l e v e l s o f T k A P X3i n r o o t s a n d l e a v e s i n c r e a s e d s i g n i f i c a n t l y c o m p a r e dw i t h t h e c o n t r o l i n d i c a t e d t h a t T k A P X3m a y b e i n v o l v e d i n t h e s t r e s s r e s p o n s e a n d h o r m o n e r e s p o n s e i n T.k o kGs a g h y z.K e y w o r d s:T a r a x a c u mk o kGs a g h y z;a s c o r b a t e p e r o x i d a s e;p h y l o g e n y a n a l y s i s;g e n e e x p r e s s i o na n a l y s i s㊀㊀橡胶草(T a r a x a c u mk o kGs a g h y z R o d i n,简称T K S),属菊科(A s t e r a c e a e)蒲公英属(T a r a x a c u m)大角蒲公英组(S e c t.M a c r o c o r n u t a V.S o e s t),与巴西橡胶树㊁银胶菊并称世界三大产胶植物,又俗称俄罗斯蒲公英(R u s s i a nd a n d e l i o n)[1],或者新疆橡胶草(X i n j i a n g R u b b e rG r a s s)[2G3],原产于哈萨克斯坦和中国的天山区域[4],在中国东北㊁华北及西北等地均有小规模野生种群分布,是一种多年生草本植物.其根部天然橡胶含量高达20%以上,个别植株可达20%~27.87%[5].橡胶草具有生长周期短,产胶量较大,适应性广,遗传转化相对容易,当年播种即可收获,并且具有适合机械规模化生产等优点,是最有发展前景的天然橡胶产胶潜在替代植物之一.随着中国天然橡胶自给率逐年下降以及对进口橡胶的依赖程度日益增强,新型天然橡胶资源的开发和高效利用是保障天然橡胶战略安全的有效途径[6].因此,在世界经济快速发展㊁中国天然橡胶刚需巨大的背景下,橡胶草将有可能成为新世纪重要的新型天然橡胶产胶替代作物,有着巨大的发展前景.生物在有氧代谢的过程中会产生活性氧(R O S),其在植物细胞应答环境因素中起着重要作用[7].H2O2作为主要的活性氧类型的信号分子,广泛参与植物细胞的发育及抗逆的调节,比如根毛的伸长㊁干旱及盐胁迫响应等[8].在生长发育及应激反应中,活性氧可通过与其他信号或植物激素的相互作用协同或拮抗许多细胞回路[9G10].生物或非生物胁迫(干旱㊁盐㊁高温㊁低温等)会使植物产生过多的活性氧物质,对细胞膜和细胞器造成不同程度的损伤[11].在天然橡胶合成过程中,乳管细胞内活性氧代谢失调会引起黄色体膜破裂,最终导致胶乳原位凝固和产胶衰竭[12].高温的胁迫还会提高氧扩散速度并且活化氧化反应,从而加速橡胶氧化反应速度,影响天然橡胶的质量[13].因此在橡胶草中的活性氧清除和相关酶的进一步利用,对天然橡胶的质量极为重要.植物体中存在酶类和非酶类两种清除细胞内活性氧的抗氧化系统,其中抗氧化物酶类系统中包括了超氧化物歧化酶(S O D)㊁过氧化氢酶(C A T)㊁谷胱甘肽还原酶(G R)㊁抗坏血酸过氧化物酶(A P X)等[14].抗坏血酸过氧化物酶(A P X;E C1.11.1.11)是含有血红素的过氧化物酶家族,它以抗坏血酸作为特异性电子供体,催化H2O2生成水,通过清除活性氧维持细胞氧化还原稳态[15].在高等植物中, A P X基因多以家族的形式存在,并根据不同的亚细胞定位分为4种类型,分别为细胞质型㊁叶绿体型㊁线粒体型和微体型[16].拟南芥A P X有8个家族成员,其中3个位于细胞质,2个位于叶绿体,另外3个位于过氧化物酶体.目前,棉花(G o s s y p i u m h i r s u t u m)[11]㊁水稻(O r y z as a t i v a)[17]㊁菠菜(S p iGn a c i a o l e r a c e a)[14]等物种的A P X基因家族成员已经被鉴定分析.优质橡胶草品系的培育及推广将成为天然橡胶商业化应用的重要原料来源,是应对当前中国大量进口天然橡胶问题的途径之一.橡胶草较易进行组织培养和遗传转化,可以利用分子改良的方法创制新材料,用于橡胶草的品系培育,但前提是需要对其生长发育及各种抗逆境的分子机制开展相关研究.本研究从橡胶草基因组中鉴定出7个T k A P X s家族成员,利用生物信息学技术全面分析橡胶草A P X 基因家族成员的染色体定位㊁基因结构㊁进化关系㊁启动子区域的顺式作用元件及表达模式等,为深入探索T k A P X在橡胶草生长发育及响应非生物胁迫过程中的分子机制提供一定的理论基础.1㊀材料和方法1.1㊀材㊀料实验于2018~2019年进行,选取生长良好㊁大小一致的播种后20d橡胶草无菌幼苗为材料,分别进行以下处理:1)低温处理:将环境温度(24ʃ2)ħ下生长的植株转移到4ħ的生长室.2)高温处理:将环境温度(24ʃ2)ħ下生长的植株转移到37ħ的生长室.6391西㊀北㊀植㊀物㊀学㊀报㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀39卷3)干旱处理:将植株从培养基中取出,用自来水洗净根部后,迅速浸泡于10%P E G6000溶液中.4)盐分处理:将植株从培养基中取出,自来水洗净根部后迅速浸泡于200m m o l/LN a C l溶液中.5)激素处理:叶面分别喷施100m g/L乙烯利和0.8m m o l/L茉莉酸甲酯,以喷施含2.2%无水乙醇的蒸馏水处理的橡胶草为对照.收集对照(未处理)及以上处理3㊁6㊁12和24h 后植物材料的叶片及根,用于分析T k A P X响应胁迫及激素的表达情况,每个处理3次重复.选取生长于人工气候室的橡胶草植株,采集不同生长阶段不同组织(花萼㊁花瓣㊁花梗㊁主根㊁侧根㊁茎㊁幼叶㊁老叶㊁一年胶乳㊁两年胶乳),用于T k A P X 组织特异性表达分析.1.2㊀方㊀法1.2.1㊀橡胶草A P X基因家族成员的鉴定㊀从N CGB I上获得橡胶草基因组数据库,利用已经鉴定的拟南芥(A r a b i d o p s i s t h a l i a n a)及水稻(O r y z a s a t i v a) A P X基因家族成员的蛋白序列作为查询序列,通过本地B l a s t p得到橡胶草A P X候选基因(e v a l u e 小于1e-10)[18].对所获得的序列利用I n t e r P r o S c a n (h t t p://w w w.e b i.a c.u k/i n t e r p r o/)检测是否有A P X基因保守结构域(I P R002016),得到7个最终确定的橡胶草A P X基因家族成员.使用E x p a s y数据库的在线分析工具C a l c u l a t e(h t t p://c a.e x p a s y.o r g/t o o l s/p i_t o o l.h t m l)预测蛋白相对分子质量和等电点[19].利用在线软件S o f t b e r r y(h t t p://l i n u x1.s o f t b e r r y.c o m/)分析其亚细胞定位[20].1.2.2㊀染色体定位和复制分析㊀用M a p I n s p e c t软件[18]分析橡胶草A P X基因的染色体分布情况.用D N AMA N软件分析橡胶草A P X基因间的亲缘关系,核酸序列相似性大于90%定义为复制基因[21].1.2.3㊀M o t i f分析㊀利用在线软件M E M E(h tGt p://m e m eGs u i t e.o r g/t o o l s/m e m e)分析氨基酸序列,得到橡胶草A P X基因的保守基序[22].1.2.4㊀系统进化树构建及基因结构分析㊀使用分子进化分析软件M E G A6.0的C l u s t a lW程序[23]对橡胶草㊁拟南芥㊁水稻及向日葵(H e l i a n t h u sa n nGu u s)A P X蛋白序列进行比对,采用相邻连接法(n e i g h b o rGj o i n i n g,N J)构建系统发育树,对构建的发育树进行评估.使用基因结构在线分析软件G SGD S(h t t p://g s d s.c b i.p k u.e d u.c n/i n d e x.p h p),通过比较橡胶草A P X基因的C D S与其相应的基因组序列来分析内含子G外显子结构[24].1.2.5㊀启动子区域的顺式作用元件分布㊀为了研究橡胶草A P X基因启动子区域中的顺式作用元件,从橡胶草基因组数据库中检索每个A P X基因起始密码子上游1500b p区域的序列.然后将这些序列提交到在线分析软件P l a n t C A R E(h t t p:// b i o i n f o r m a t i c s.p s b.u g e n t.b e/w e b t o o l s/p l a n t c a r e/ h t m l/)以预测其顺式作用元件[25].1.2.6㊀基因特异性表达分析㊀使用天根植物R N A 提取试剂盒(D P441GH)提取橡胶草的总R N A, T h e r m o s c i e n t i f i c R e v e r t A i d F i r s t S t r a n d c D N A S y n t h e s i sK i t用于将总R N A反转为c D N A.利用P r i m e r软件设计T k A P X1G7的特异性引物.使用C h a m Q T M U n i v e r s a lS Y B R Q p c r M a s t e r M i x在A B I S t e p O n e P l u s上进行实时荧光定量P C R.采用2-әәC T法计算基因的相对表达水平,T k A c t i n作为内参基因,每个基因3个独立的生物学重复.热图使用M u l t i E x p e r i m e n tV i e w e r(M e V)软件绘制[20].2㊀结果与分析2.1㊀橡胶草A P X基因家族成员鉴定在橡胶草基因组数据库中,利用拟南芥和水稻A P X基因家族蛋白序列进行B l a s t p分析,经过保守结构域确认,得到7个橡胶草T k A P X s基因,分布于7个独立的基因组组装脚手架片段(s c a f f o l d)上.根据其所在s c a f f o l d的大小分别命名为T k A P X1㊁T k A P X2㊁T k A P X3㊁T k A P X4㊁T k A P X5㊁T k A P X6和T k A P X7(图1).橡胶草A P X基因家族成员的开放阅读框长度分布于753b p(T k A P X3㊁T k A P X5)至1341b p(T k A P X1)之间,由其编码的蛋白质氨基酸长度为250~446a a.蛋白质相对分子质量在27.36~50.18k D之间,等电点最小为5.08(T k A P X1),最大为8.1(T k A P X4㊁T k A P X6).亚细图1㊀A P X基因在橡胶草染色体上的定位F i g.1㊀L o c a t i o no f A P X g e n e s o n c h r o m o s o m e s o fT a r a x a c u mk o kGs a g h y z739111期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀高㊀璇,等:橡胶草A P X基因家族的全基因组鉴定及表达分析表1㊀橡胶草A P X 基因家族信息T a b l e 1㊀I n f o r m a t i o no f A P X g e n e f a m i l y i n T a r a x a c u mk o k Gs a g h yz 基因G e n e 基因编号G e n e I D 开放阅读框O R F /b p P r o t e i n 蛋白长度S i z e /a a 分子量MW /k D等电点p I 亚细胞定位S u b c e l l u l a r l o c a l i z a t i o nT k A P X 1e v m.T U.u t g 5421.9134144650.185.08细胞质C y t o p l a s m i c T k A P X 2e v m.m o d e l .u t g 8943.10105335038.036.77叶绿体C h l o r o pl a s t T k A P X 3e v m.m o d e l .u t g 5260.675325027.525.23细胞质C y t o p l a s m i c T k A P X 4e v m.m o d e l .u t g 9528.10129343046.498.10质膜P l a s m am e m b r a n e T k A P X 5e v m.m o d e l .u t g26446.1275325027.365.54细胞质C y t o p l a s m i c T k A P X 6e v m.m o d e l .u t g 448.5129343046.468.1质膜P l a s m am e m b r a n eT k A P X 7e v m.T U.u t g18379.585528431.285.85细胞质C y t o p l a s m ic 图2㊀橡胶草A P X 家族成员M o t i f 分析F i g .2㊀M o t i f a n a l y s i s o f t h eT k A P Xf a m i l y me m b e r s 胞定位结果表明,T k A P X 1㊁T k A P X 3㊁T k A P X 5和T k A P X 7定位于细胞质,T k A P X 4和T k A P X 6定位于质膜,T k A P X 2定位于叶绿体(表1).2.2㊀橡胶草A P X 基因家族蛋白保守基序分布分析为了研究橡胶草A P X 蛋白序列的保守性,进行了保守基序分布分析,利用M E M E 对橡胶草A P X蛋白序列进行分析,预测到10个保守基序.发现亲缘关系相近的成员具有相似的保守基序分布(图2).每个T k A P X m o t i f 数目在2~10之间,m o t i f 1和m o t i f 7存在于所有T k A P X 中.T k A P X 1㊁T k A P X 3㊁T k A P X 5和T k A P X 7具有相同的m o t i f.m o t i f 8和m o t i f 9为T k A P X 4和T k A P X 6特有.2.3㊀橡胶草A P X 基因家族进化分析为了明确橡胶草A P X 基因家族与其他物种间的进化关系,利用拟南芥(8个A P X 基因)㊁水稻(8个A P X 基因)㊁向日葵(9个A P X 基因)及橡胶草A P X 基因家族的蛋白序列构建系统进化树(图3),根据进化分析,将其分为5个亚组.T k A P X 3和T k .橡胶草;A t .拟南芥;O s .水稻;H a .向日葵图3㊀橡胶草和其他植物A P X 基因家族进化树T k .T a r a x a c u mk o k Gs a g h y z ;A t .A r a b i d o ps i s t h a l i a n a ;O s .O r yz a s a t i v a ;H a .H e l i a n t h u s a n n u u s F i g .3㊀P h y l o ge n e t i c t r e e o fA P Xf a m i l i e s i n T a r a x a c u mk o k Gs agh yz a n do t h e r p l a n t s 8391西㊀北㊀植㊀物㊀学㊀报㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀39卷T k A P X5聚类到A亚组,T k A P X1和T k A P X7聚类到B亚组,T k A P X4和T k A P X6属于C亚组, T k A P X2属于E亚组,而D亚组没有橡胶草A P X 基因家族的成员,可能是因为在长期进化过程中T k A P X的功能发生了分化.根据以上分析可以看出,橡胶草不同的A P X家族成员之间存在着一定的进化差异,推测可能通过多重机制来清除细胞内的活性氧,达到抗氧化的效果.由图4可以看出,T k A P X的外显子数目为7~12个,且亲缘关系相近的成员外显子数目保持一致,如T k A P X1和T k A P X7,鉴定的复制基因(T k A P X4/T k A P X6)具有完全一致的内含子(i nGt r o n)G外显子(e x o n)结构.2.4㊀橡胶草A P X基因家族启动子区域的顺式作用元件分布㊀㊀翻译起始位点上游的1500b p序列用来分析顺T k.橡胶草;A t.拟南芥图4㊀橡胶草A P X基因家族进化及基因结构T k.T a r a x a c u mk o kGs a g h y z;A t.A r a b i d o p s i s t h a l i a n aF i g.4㊀P h y l o g e n e t i c a n de x o nGi n t r o n s t r u c t u r e a n a l y s e s o f A P X g e n e s i n T a r a x a c u mk o kGs a g h yz图5㊀T k A P X启动子区域的顺式作用元件分布F i g.5㊀D i s t r i b u t i o no f c i sGe l e m e n t s i n t h e p r o m o t e r r e g i o n s o f T k A P X g e n e s939111期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀高㊀璇,等:橡胶草A P X基因家族的全基因组鉴定及表达分析式作用元件的分布,根据系统发育关系将T k A P X 分为4组.不同的顺式作用元件用不同颜色的大写字母表示(图5),其中包括2个转录起始(红色)㊁6个响应植物激素反应(绿色)和17个应激反应(紫色)的顺式作用元件,这意味着应激反应和激素共同调控橡胶草A P X的表达.从图5可以看出,复制基因T k A P X4/T k A P X6启动子具有相似的顺式作用元件分布.2.5㊀橡胶草A P X基因家族成员表达模式分析图6显示,7个T k A P X s家族成员在花萼㊁花瓣㊁花梗中表达量均较低;复制基因T k A P X4/ T k A P X6在8个不同组织中的表达水平相似; T k A P X5在8个组织中均低丰度表达;随着橡胶草的生长发育,叶和胶乳中的T k A P X1㊁T k A P X2㊁T k A P X7表达水平有不同程度的上升,且T k A P X1在根和茎中的表达水平最高.大多数T k A P X在根茎叶中的表达水平大于花及胶乳.分析乙烯及茉莉酸甲酯处理后A P X基因家族成员的表达规律发现:在叶中除了T k A P X3,其他均低于对照材料叶中对应的T k A P X的表达水平,推测T k A P X3在橡胶草响应乙烯及茉莉酸信号的过程中起重要作用,其他家族成员反向调节,尤其是T k A P X5的表达水平显著低于对照,对激素处理表现负反馈;茉莉酸甲酯处理后的橡胶草根部中,7个T k A P X s表达量与叶中规律完全不同,除了T k A P X5的表达水平显著低于对照,其余6个家族成员表达水平均有不同程度的升高,其中T k A P X1及T K A P X3提高的程度最为明显,且均为6h达到最高;乙烯处理后的橡胶草根中,却是只有T K A P X3表达量升高,其他成员表达量均降低.低温㊁高温和盐分处理后,橡胶草的根及叶中T K A P X3表达水平也均大幅度升高,说明其在响应冷㊁热㊁盐胁迫中起着重要作用.其中,P E G处理后叶及根中T K A P X3出现先升高后降低的趋势;图6㊀T k A P X表达模式分析F i g.6㊀E x p r e s s i o n p a t t e r na n a l y s i s o f T k A P X0491西㊀北㊀植㊀物㊀学㊀报㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀39卷T k A P X1表达水平在高低温和盐分处理后叶中均变化不大,只有干旱胁迫后有降低;T K A P X5的表达在各种胁迫后均大幅降低.3㊀讨㊀论A P X是重要的抗氧化酶类之一,对于维持植物体内活性氧代谢平衡有着重要作用[7,26].当高等植物受到生物或者非生物胁迫时,A P X会调控一系列生理生化反应,快速清除植物细胞中过量的H2O2,使细胞免受活性氧的毒害.目前对A P X基因克隆及功能的研究主要集中在拟南芥㊁棉花等植物,而在橡胶草中却罕见报道.橡胶草基因组数据的获得为其基因家族的分析提供了基础,便于进一步了解基因功能及调控机制.本研究从橡胶草的全基因组序列中鉴定出7个A P X家族成员,与拟南芥(8个)㊁水稻(8个)㊁玉米(9个)相近.通过进化分析,可以看出亲缘关系近的成员具有相同的亚细胞定位,比如1对复制基因T k A P X4和T k A P X6均定位于细胞质膜.基于不同成员的亚细胞定位分析结果,可以推测他们的功能具有多样性,通过多细胞器配合,多重机制清除细胞内多余的活性氧.基因结构的差异在基因家族的进化过程中起着重要的作用[27].对拟南芥及橡胶草的A P X基因家族成员基因结构分析,发现同一个亚组的成员具有相似但不完全相同的内含子㊁外显子结构,表明在长期的进化过程中T k A P X的功能发生了分化.错义突变K a和同义突变K s比值可用来估算复制基因在进化过程中受到的自然选择类型.如K a/K s大于1,则认为有正选择效应;反之则认为有纯化选择作用;如K a/K s等于1,则为中性选择.本研究对具有复制关系的T k A P X4/T k A P X6进行K a/K s 计算,结果小于1,处于纯化选择.从启动子区域的顺式作用元件分析可看出,橡胶草A P X基因的启动子均含有大量的应激反应元件,由此可推测其表达可能受应激反应严格控制.实时荧光定量P C R结果表明,T k A P X1在未做处理的不同组织中均表达且表达量高于其他家族成员,推断其在橡胶草生长发育中起着更重要作用. T k A P X5在不同组织中均低丰度表达.复制基因T k A P X4/T k A P X6表达模式基本一致,由此推测其在橡胶草生长发育中发挥着相似的作用.在4ħ㊁37ħ㊁N a C l㊁P E G处理橡胶草后,根和叶中的T k A P X3表达水平均上升,说明其可能在橡胶草响应胁迫中起着重要的作用.T K A P X5的表达在各种胁迫后表现了负反馈,均大幅降低,推测其有可能在相关的分子机制中有重要的作用.4ħ冷处理橡胶草叶后,T k A P X4/T k A P X6表达水平先升高后下降到处理前水平,T k A P X1的表达量变化不大.茉莉酸甲酯是一种重要的植物激素,其与植物的生长发育及抗逆反应息息相关.马海霞等[28]研究发现,一月生的橡胶草叶面喷施M e J A促进橡胶草乳管分化,同时M e J A诱导植物活性氧的积累[29],橡胶草的主根长度㊁主根粗度㊁叶绿素含量㊁根系活力㊁P A L活性和木质素含量均高于对照处理,还有利于橡胶草植株抗性的增强.本研究发现M e J A处理橡胶草的根后,T k A P X1和T k A P X3表达水平均提高,且T k A P X1在胶乳中的表达水平相对较高,推测其可能在胶乳活性氧代谢调控中起着重要的作用.本研究首次鉴定了橡胶草A P X基因家族,基于系统进化关系将7个基因分为4个亚组,并鉴定出1对复制基因(T k A P X4/T k A P X6).进一步的表达分析表明,该家族可能在橡胶草生长发育及应激反应中起重要作用.可作为后续研究的候选基因,开展相关功能验证的研究,并用于橡胶草分子改良新材料的创制.参考文献:[1]㊀中国科学院中国植物志编辑委员会.中国植物志[M].北京:科学出版社,1999.[2]㊀谢全亮,李鸿彬,王旭初.橡胶草90年来主要研究成果及最新研究进展[J].植物科学学报,2019,37(3):404G412.X I E QL,L IH B,WA N GXC.R e c e n t p r o g r e s s i n t h e s t u d y o fr u b b e r g r a s s(T a r a x a c u m k o kGs a g h y z R o d i n)a n d m a i na c h i e v e m e n t so v e rt h e p a s tn i n e t yy e a r s[J].P l a n tS c i e n c eJ o u r n a l,2019,37(3):404G412.[3]㊀X I E QL,D I N G G H,Z HU LP,e t a l.P r o t e o m i c l a n d s c a p e o f t h e m a t u r er o o t s i nar u b b e rGp r o d u c i n gg r a s s T a r a x a c u mk o kGs a g h y z[J].I n t e r n a t i o n a l J o u r n a l o f M o l e c u l a rS c i e n c e s,2019,20(10):2596.[4]㊀K I R S C H N E RJ,ŠTĚP N E KJ,C㊅E R N T,e t a l.A v a i l a b l ee xs i t u g e r m p l a s m o ft h e p o t e n t i a lr u b b e rc r o p T a r a x a c u m k o kGs a g h y z b e l o n g s t oa p o o rr u b b e r p r o d u c e r,T.b r e v i c o rGn i c u l a t u m(C o m p o s i t a eGC r e p i d i n a e)[J].G e n e t i c R e s o u r c e s a n dC r o p E v o l u t i o n,2013,60(2):455G471.[5]㊀仇㊀键,张继川,罗世巧,等.橡胶草的研究进展[J].植物学报,2015,50(1):133G141.Q I UJ,Z HA N GJC,L U OSQ,e t a l.R e s e a r c h a d v a n c e s a n d p e r s p e c t i v e so n r u b b e rGp r o d u c i n g T a r a x a c u m[J].C h i n e s e149111期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀高㊀璇,等:橡胶草A P X基因家族的全基因组鉴定及表达分析B u l l e t i no f B o t a n y,2015,50(1):133G141.[6]㊀高玉尧,刘㊀洋,许文天,等.不同施肥处理对橡胶草生物量积累与分配变化及相关性分析[J].分子植物育种,2018,16(9):2979G2986.G A O Y Y,L I U Y,X U W T,e t a l.C h a n g e s o f b i o m a s s a c c uGm u l a t i o na n dc o r r e l a t i o na n a l y s i si nd i f f e r e n tf e r t i l i z e rt r e a tGm e n t s i n T a r a x a c u mk o kGs a g h y z[J].M o l e c u l a rP l a n tB r e e dGi n g,2018,16(9):2979G2986.[7]㊀P A N D E Y S,F A R T Y A L D,A G A RWA L A,e t a l.A b i o t i c s t r e s s t o l e r a n c e i n p l a n t s:m y r i a d r o l e s o f a s c o r b a t e p e r o x i d a s e [J].F r o n t i e r s i nP l a n t S c i e n c e,2017,8:581.[8]㊀Q U AL,D I N GYF,J I A N GQ,e t a l.M o l e c u l a rm e c h a n i s m s o f t h e p l a n th e a ts t r e s sr e s p o n s e[J].B i o c h e m i c a la n d B i oGp h y s i c a lR e s e a r c hC o m m u n i c a t i o n s,2013,432(2):203G207.[9]㊀P A T E R S O N A H,W E N D E LJF,e t a l.R e p e a t e d p o l y p l o i d i z aGt i o n o f G o s s y p i u m g e n o m e s a n d t h e e v o l u t i o n o f s p i n n a b l e c o t t o n f iGb r e s[J].N a t u r e,2012,492(7429):423G427.[10]㊀T A O C C,J I N X,Z HU L P,e t a l.T w oGd i m e n s i o n a l g e le l e c t r o p h o r e s i sGb a s e d p r o t e o m i ca n a l y s i sr e v e a l s NGt e r m i n a lt r u n c a t i o no f t h eH s c70p r o t e i n i nc o t t o n f i b e r s i n v i v o[J].S c i e n t i f i c R e p o r t s,2016,6:36961.[11]㊀刘㊀娜,吴金华,安亚茹,等.陆地棉抗坏血酸过氧化物酶基因家族全基因组生物信息学分析[J].棉花学报,2017,29(1):17G28.L I U N,WUJH,A N YR,e t a l.B i o i n f o r m a t i c s a n a l y s i s o ft h e a s c o r b a t e p e r o x i d a s e g e n e f a m i l y i n t h e G o s s y p i u mh i r s uGt u m L.g e n o m e[J].C o t t o n S c i e n c e,2017,29(1):17G28.[12]㊀郭秀丽.橡胶树H b G R X基因的克隆及其在死皮病发生过程中的功能分析[D].海口:海南大学,2017.[13]㊀吴文剑.大分子受阻酚抗氧剂的合成与表征及其对天然橡胶抗热氧老化作用的研究[D].广州:华南理工大学,2015.[14]㊀郁征宇,葛晨辉,王小丽,等.菠菜抗坏血酸过氧化物酶基因家族的鉴定及表达分析[J].上海师范大学学报(自然科学版),2018,47(6):664G672.Y UZY,G ECH,WA N GXL,e t a l.I d e n t i f i c a t i o n a n d e xGp r e s s i o na n a l y s i s o f a s c o r b a t e p e r o x i d a s e g e n e f a m i l y i n s p i nGa c h[J].J o u r n a l o f S h a n g h a iN o r m a lU n i v e r s i t y(N a t u r a lS c i e n c e s),2018,47(6):664G672.[15]㊀乔㊀枫,耿贵工,曾㊀阳,等.枸杞抗坏血酸过氧化物酶基因的克隆与表达分析[J].中国农业大学学报,2019,24(4):64G72.Q I A OF,G E N G G G,Z E N G Y,e t a l.M o l e c u l a rc l o n i n ga n de x p r e s s i o n p a t t e r n so f L c A P X f r o m L y c i u mc h i n e n s e[J].J o u r n a l o f C h i n aA g r i c u l t u r a lU n i v e r s i t y,2019,24(4):64G72.[16]㊀S H I G E O K AS.R e g u l a t i o na n d f u n c t i o no f a s c o r b a t e p e r o x iGd a s ei s oe n z y m e s[J].J o u r n a lof E x p e r i m e n t a l B o t a n y,2002,53(372):1305G1319.[17]㊀T E I X E I R AFK,M E N E Z E SGB E N A V E N T EL,G A L VÃOV C,e t a l.R i c e a s c o r b a t e p e r o x i d a s e g e n e f a m i l y e n c o d e s f u n cGt i o n a l l y d i v e r s ei s o f o r m sl o c a l i z e di n d i f f e r e n t s u b c e l l u l a rc o m p a r t m e n t s[J].P l a n t a,2006,224(2):300G314.[18]㊀T A OCC,J I N X,Z HU LP,e t a l.G e n o m eGw ide i n v e s t i g aGt i o na n d e x p r e s s i o n p r of i l i ng o f A P X g e n e f a m i l y i n G o s s y p iGu mh i r s u t u m p r o v i d e n e w i n s i g h t s i n r e d o xh o m e o s t a s i sm a i nGt e n a n c e d u r i n g d i f f e r e n t f i b e r d e v e l o p m e n t s t a g e s[J].M o l e cGu l a r G e n e t i c s a n dG e n o m i c s:M G G,2018,293(3):685G697.[19]㊀A R T I MO P,J O N N A L A G E D D A M,A R N O L D K,e t a l.E x P A S y:S I B b i o i n f o r m a t i c sr e s o u r c e p o r t a l[J].N u c l e i cA c i d sR e s e a r c h,2012,40(W1):W597GW603.[20]㊀J I NX,Z HULP,Y A OQ,e t a l.E x p r e s s i o n p r o f i l i n g o fm iGt o g e nGa c t i v a t e d p r o t e i nk i n a s e g e n e sr e v e a l s t h e i re v o l u t i o nGa r y a n df u n c t i o n a l d i v e r s i t y i nd i f f e r e n t r ub b e r t r e e(H e v e ab r a s i l i e n s i s)c u l t i v a r s[J].G e n e s,2017,8(10):261.[21]㊀T A N G K,D O N GCJ,L I UJY.G e n o m eGw ide c o m p a r a t i v ea n a l y s i s o f t h e p h o s p h o l i p a s eD g e n e f a m i l i e s a m o n g a l l o t e tGr a p l o i dc o t t o na n di t sd i p l o i d p r o g e n i t o r s[J].P L o S O n e,2016,11(5):e0156281.D O I:10.1371/j o u r n a l.p o n e.0156281.㊀[22]㊀B A I L E Y T L,W I L L I AM SN,M I S L E H C,e t a l.M E M E:d i s c o ve r i n g a n da n a l y z i n g D N Aa n d p r o t e i ns e q u e n c em o t if s[J].N u c l e i cA c i d sR e s e a r c h,2006,34(W e bS e r v e r i s s u e):369G373.[23]㊀T A M U R A K,S T E C H E R G,P E T E R S O N D,e t a l.M E G A6: m o l e c u l a r e v o l u t i o n a r y g e n e t i c s a n a l y s i s v e r s i o n6.0[J].M o l e c uGl a r B i o l o g y a n dE v o l u t i o n,2013,30(12):2725G2729.[24]㊀HU B,J I NJ,G U OA Y,e t a l.G S D S2.0:a n u p g r a d e d g e n ef e a t u r ev i s u a l i z a t i o ns e r v e r[J].B i o i n f o r m a t i c s,2014,31(8):1296.D O I:10.1093/b i o i n f o r m a t i c s.[25]㊀L E S C O T M,DÉH A I SP,T H I J S G,e t a l.P l a n t C A R E,ad a t a b a s eo f p l a n t c i sGa c t i n g re g u l a t o r y e l e m e n t s a n da p o r t a lt o t o o l s f o r i n s i l i c o a n a l y s i s o f p r o m o t e r s e q u e n c e s[J].N uGc l e i cA c id sRe s e a r c h,2002,30(1):325G327.[26]㊀王亦学,杜建中,郝曜山,等.过量表达G h A P X基因提高中林美荷杨耐盐性[J].分子植物育种,2017,15(7):2579G2583.WA N G Y X,D UJZ,H A O YS,e t a l.O v e rGe x p r e s s i o no fG h A P X g e n e i n t h e i m p r o v e m e n t o f s a l t t o l e r a n c e i nP o p u l u s s p[J].M o l e c u l a r P l a n tB r e e d i n g,2017,15(7):2579G2583.[27]㊀Z HU XD,WA N G M Q,L I XP,e t a l.G e n o m eGw i d e a n a l yGs i s o f t h e s u c r o s e s y n t h a s e g e n e f a m i l y i n g r a p e(V i t i s v i n i fGe r a):s t r u c t u r e,e v o l u t i o n,a n d e x p r e s s i o n p r of i l e s[J].G e n e s,2017,8(4):E111.[28]㊀马海霞,何丽娜,海春燕,等.茉莉酸甲酯对橡胶草生理生化及乳管分化的影响[J].石河子大学学报(自然科学版),2017,35(4):451G457.MA H X,H ELN,H A ICY,e t a l.E f f e c t s o fm e t h y l j a sGm o n a t e o n p h y s i o l o g y,b i o c h e m i s t r y a n d l a t i c i f e r d i f f e r e n t i aGt i o no f T a r a x a c u mk o kGs a g h y z[J].J o u r n a l o f S h i h e z iU n iGv e r s i t y(N a t u r a l S c i e n c e),2017,35(4):451G457.[29]㊀吕静祎,张㊀睿,张梦媛,等.茉莉酸甲酯处理对采后苹果果皮活性氧相关代谢的影响[J].食品工业科技,2018,39(7):271G275,285.LÜJY,Z H A N GR,Z HA N G M Y,e t a l.E f f e c t s o fm e t h y lj a s m o n a t e t r e a t m e n to nr e l a t e d m e t a b o l i s m o f r e a c t i v eo x yGg e ns p e c i e s i n t h e p e e l o f p o s t h a r v e s t a p p l e f r u i t[J].S c i e n c ea n d T e c h n o l o g y o f F o o d I n d u s t r y,2018,39(7):271G275,285.(编辑:宋亚珍)㊀㊀2491西㊀北㊀植㊀物㊀学㊀报㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀39卷。
橡胶树种质资源的倍性鉴定

橡胶树种质资源的倍性鉴定橡胶树(Hevea brasiliensis)是一种重要的经济林木品种,其橡胶乳是生产天然橡胶的主要来源。
橡胶树种质资源的倍性鉴定是对橡胶树进行遗传分析和育种选择的重要手段。
本文将对橡胶树倍性鉴定的方法、意义和应用进行介绍。
一、倍性的定义和意义倍性是指一个细胞或一个有机体中染色体的数量的倍数关系。
常见的倍性有二倍体(2n)、四倍体(4n)等。
倍性对植物的生长发育和性状表达有着重要影响。
比如二倍体植株的染色体数量较少,基因自由组合的可能性多,具有丰富的遗传多样性,对杂交育种有较大的潜力;而四倍体植株的染色体数量较多,可提高植株的耐逆性、生长速度和产量等性状。
二、橡胶树倍性鉴定的方法倍性鉴定可以通过多种方法进行,常见的方法有染色体观察法、流式细胞仪法和分子生物学方法。
1.染色体观察法:染色体观察法是最早也是最常用的倍性鉴定方法。
该方法是通过染色体特异的染色剂,如乙酰好酸铁配合苏丹黑、乙酰好酸铁配合鲜明黑等,将根尖组织进行染色处理后在显微镜下观察染色体的数量和形态。
二倍体植株一般有正常的染色体配对,染色体数目为2n;而多倍体植株的染色体数目会是正常染色体数量的整数倍。
染色体观察法简单、直观,但该方法只能适用于根尖组织,且对于小型或无根尖的植物难以应用。
2.流式细胞仪法:流式细胞仪法是利用细胞在倒相差结果中,根据细胞的DNA含量来进行倍性鉴定。
该方法是先用适当的缓冲液将植物组织细胞切碎,然后通过流式细胞仪进行测定。
流式细胞仪通过光散射和荧光信号的强度,能够准确地判断细胞的倍性。
流式细胞仪法可以高通量地进行倍性鉴定,能够在较短的时间内快速得到结果,但设备价格较高,且需要专门的操作技术。
3.分子生物学方法:分子生物学方法是近年来发展起来的倍性鉴定方法。
该方法是通过使用特异的引物,扩增不同倍性植株的基因组DNA,在凝胶电泳中观察扩增片段的数量差异。
分子生物学方法可以快速准确地进行倍性鉴定,且对于无根尖或小型植物也能适用,但需要较为复杂的实验操作和仪器设备。
优质高效栽培技术橡胶树的新品种选育

优质高效栽培技术橡胶树的新品种选育橡胶树作为世界上最重要的经济林木之一,其产胶量对于橡胶行业的发展至关重要。
然而,传统的橡胶树品种在抗病虫害、生长速度和产胶量方面存在一定的局限性。
因此,研究人员不断努力进行新品种选育,以提高橡胶树的优质高效栽培技术。
本文将介绍一些相关的研究进展和创新方法。
一、经典选育和遗传改良在橡胶树品种选育中,经典选育和遗传改良是最常见的方法之一。
通过对具有优良特性的橡胶树个体进行繁殖和杂交,可以逐步筛选出更加适应不同环境和具有高产胶能力的新品种。
这种方法经过多年的实践已经取得了一定的成功,许多具有良好品质和高产量的橡胶树品种得以开发。
二、分子标记辅助选育随着生物技术的发展,分子标记辅助选育成为现代橡胶树品种选育的重要手段之一。
通过检测和分析橡胶树的遗传物质,如DNA和RNA序列,可以快速筛选出具有目标性状的个体,并进行进一步繁殖。
这种方法在提高选育效率和准确性方面具有重要的意义,并且可以帮助研究人员解析橡胶树品种演化和遗传机制。
三、基因编辑技术近年来,基因编辑技术的迅速发展使得橡胶树品种选育进入了一个全新的阶段。
通过CRISPR/Cas9等方法,研究人员可以直接编辑橡胶树的基因组,实现特定基因的敲除或添加。
这种方法极大地拓宽了品种选育的可能性,可以有针对性地改良橡胶树的特征,提高产量和抗病虫害能力。
四、光合作用改良橡胶树的光合作用是产胶的重要过程,提高光合作用效率可以直接提高橡胶树的产量。
因此,研究人员致力于改良橡胶树的光合作用,提高光合效率。
例如,通过调节光合相关基因的表达,优化叶绿素的光利用效率,改善光合作用底物的供应等手段,可以显著提高橡胶树的产量。
五、生长调控技术橡胶树生长的调控对于产量和质量的提高至关重要。
提供适宜的养分供应和水分管理,合理掌握生长周期和修剪,都可以对橡胶树的生长进行调控。
近年来,利用植物生理学和遗传学的研究成果,研究人员开展了一系列的生长调控技术的研究,如激素调控、生长素依赖性突变体筛选等,以期提高橡胶树的生长速度和产量。
橡胶树产胶生物学研究进展
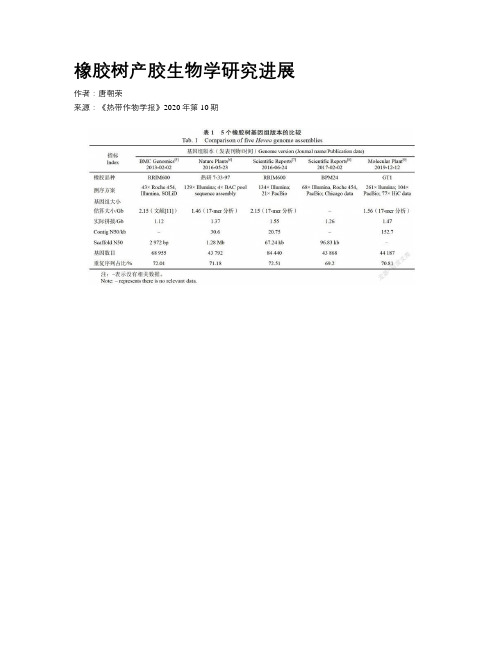
橡胶树产胶生物学研究进展作者:***来源:《热带作物学报》2020年第10期摘要:天然橡胶(顺式-1,4-聚异戊二烯)是一种重要的工业原料,主要来自橡胶树。
以天然橡胶的生物合成与产量形成为主要内容的产胶生物学研究为橡胶树高产遗传改良提供理论指导,近10年取得了重要进展。
本文从橡胶树基因组测序、橡胶转移酶、转基因、以及转录组和蛋白质组等4个方面介绍产胶生物学研究主要进展,并讨论了相关领域研究存在的问题,对未来5~10年需重点关注的研究内容提出了建议。
在介绍橡胶转移酶时,同时概述其他几种产胶植物的相关研究进展。
关键词:橡胶树;产胶生物学;基因组测序;橡胶转移酶中图分类号:S794.1 文献标识码:AAbstract: Natural rubber (cis-1, 4-polyisoprene) is an important industrial raw material,commercially harvested mainly from rubber tree (Hevea brasiliensis). Important progresses have been made in the past decade on Hevea biology of rubber production, focusing on rubber biosynthesis and latex production. The present review summarized the major progress in four research areas with relation to rubber production, i.e. Hevea genome sequencing, rubber transferase,transgenic research, and transcriptomics & proteomics. Moreover, the problems were discussed and a few of research emphasis in the next 5 to 10 years were proposed. When describing the studies of rubber transferase, the recent progress in several other rubber-producing plants were included.Keywords: Hevea brasiliensis (para rubber tree); biology of rubber production; genome sequencing; rubber transferaseDOI: 10.3969/j.issn.1000-2561.2020.10.003天然橡胶是一种重要的工业原料,在交通运输、医疗卫生、国防军工等领域应用广泛,其优异的综合性能迄今仍无法被人工合成橡胶完全替代[1]。
橡胶树 基因组

橡胶树基因组橡胶树基因组:揭示橡胶生产机制的关键橡胶是一种重要的工业原料,广泛应用于汽车、建筑、航空航天等领域。
而橡胶的主要来源是橡胶树,其胶乳中含有丰富的天然橡胶。
然而,橡胶树的生产效率和品质受到许多因素的影响,如病虫害、环境因素等。
因此,研究橡胶树的基因组,揭示其生产机制,对提高橡胶生产效率和品质具有重要意义。
橡胶树基因组测序是揭示橡胶生产机制的关键。
在过去的几年中,科学家们已经完成了橡胶树的基因组测序。
通过对基因组序列的分析,人们可以了解橡胶树的基因组结构、基因数目、基因功能等信息。
此外,还可以通过比较不同树种的基因组序列,发现橡胶生产过程中所涉及的关键基因。
橡胶树基因组测序揭示了橡胶树的基因组结构和基因数目。
橡胶树基因组约有2.15亿个碱基对,包括85,446个基因。
这些基因编码了许多不同的蛋白质,这些蛋白质在橡胶生产过程中起着关键作用。
例如,其中的一些基因编码了橡胶合成酶,这些酶可以将橡胶乳中的异戊二烯聚合成天然橡胶。
除了基因数目和基因功能,橡胶树基因组测序还揭示了许多其他重要的信息。
例如,橡胶树基因组中有许多转座子元件,这些元件可以影响基因的表达和调节。
此外,基因组测序还揭示了橡胶树基因组的染色体数量和结构,这对于进一步研究橡胶生产机制具有重要意义。
橡胶树基因组测序不仅可以揭示橡胶生产机制的关键,还可以为橡胶树的育种和改良提供重要的信息。
通过对基因组序列的分析,可以找到与橡胶生产相关的基因,从而选择合适的育种材料,加速橡胶树的育种进程。
此外,基因组测序还可以帮助人们了解橡胶树的适应性和抗性机制,从而开发更加耐病、耐旱的橡胶树品种。
橡胶树基因组的测序为揭示橡胶生产机制提供了重要的信息。
未来,科学家们将继续研究橡胶树的基因组,深入了解橡胶生产的机制,推动橡胶生产的发展。
基因枪法获得GAI转基因橡胶树植株的研究

进行 抗性 筛选 。获得 了抗 性 再生 植株 , 经过 P R、 ote C S uhr n检测 结果 表 明: A 基 因 已经成 功 转入 橡 胶树 基 因 G1
(. 1 中国热带农业科学院热带生物技术研究所, 农业部热带作物生物技术重点开放实验室, 口 5 10 ; . 海 7 1 1 2 庐江中学 , 安徽 庐江 2 10 ) 3 5 0
摘要 : 将含 有 C mv5 a 3 S启 动子 、 卡那 霉 素抗 性基 因和 G S报 告基 因, U 目的基 因为 G I基 因 的转 化 载体 质 粒 A
A b t a t Th v c o I 21 p ril,w hc c n ans sr c : e e t r pB 1 a t e c ih o ti Ca v m 35S r m o e ,ka a ycn r ssa e e e G U p o tr nm i e itnc g n , S r p re e ea d GAIg ne w a r nse e o t v a b a ie ss a t e a usb a  ̄ e o trg n n e , sta f r d i heHe e r sl n i n h r c n y p r nt i t bo badm e t Th m r n. e
然橡 胶 的主要来 源 , 由于我 国橡胶 产 区受 台风影 响 较 大 , 害严 重制 约着 我 国天然 橡胶 的发展 。对 巴 风
西橡 胶树 的转 基 因研 究 已有 报 道 l Aoia等 l rk r 】 , aj
巴西橡胶树的基因组序列框架图

( NR ) 的 主 要 来 源 。NR是 一 种 具 有 高 弹 性 、 高 可 塑 性 及 高 回 弹 力 的 胶乳 聚 合 物 , 自1 8 7 6 年 以来 ,NR在 世 界 经 济 发 展 中一 直 都 起 着 至关 重 要 的 作 用 。本 文 报 道 巴西 橡 胶 树 的基 因组 序 列 框 架 图 。 已 组 装 的基 因组序列大约 1 . 1 G b , 约 占 整个 单倍 体 基 因 组 估 计 长 度 ( 2 . 1 5 G h ) 的5 1 。该 基 因 组 中大 约 有 7 8 为
( 中 国热 带 农 业 科 学 院橡 胶 研 究所 , 海南 儋州 5 7 1 7 3 7 )
摘
要: 巴西 橡 胶 树 ( He v e a b r a s i l i e n s i s ) 为 大 戟科 多 年 生 木 本 植 物 , 也 是 迄 今 为 止 全 世 界 商 品 天 然 橡 胶
Tr a n s l a t e d by TI AN La n g
Ru b b e r R e s e a r c h I n s t i t u t e , C h i n e s e Ac a d e my o f T r o p i c a l Ag r i c u l t u r a l S c i e n c e s ,
关键 词 : 巴西 橡 胶 树 ; 大戟科 ; 天然橡胶 ; 基 因组
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
橡胶树的基因组学研究
橡胶树是一种重要的经济作物,广泛栽培于东南亚、非洲和南美洲的热带和亚
热带地区。
橡胶树的橡胶是一种重要的工业原料,广泛用于轮胎、鞋底、橡胶管、橡胶制品、涂料、粘合剂等领域,其全球产量居于第一位,是非常重要的经济资源。
然而,橡胶树长期以来未被广泛的研究,其基因组学、生化和生理学等方面的研究仍然存在很多挑战。
为了更好地了解橡胶树的基因组学特征并实现优质化的种植和开发利用,橡胶树的基因组学研究变得十分重要。
一、橡胶树基因组测序工作
在过去的十年中,随着高通量测序技术的发展和应用,许多生物学领域的基因
组学研究进展较快。
这些技术的发展也促进了针对橡胶树的基因组学研究。
2008年,研究人员对橡胶树生长中期进行了转录组测序。
2013年,中科院深圳先进技
术研究院和马来西亚棕榈油研究院协同研究组对橡胶树RG1基因组进行了高质量
的测序,并于2016年公开发布了其进行基因组学研究所需的所有数据。
二、橡胶树基因组学研究的进展
1. 橡胶树基因组的组装和注释
对于任何一个生物体而言,正确的基因组测序和组装都是基础性的工作。
橡胶
树基因组的组装工作主要是通过第二代测序技术(Sanger和Illumina)和第三代测
序技术(PacBio和Oxford Nanopore)来完成的。
目前,针对橡胶树基因组的组装
已经较为完整,48K SNP的核心基因组采用Pacbio的更长读取数据完成。
在组装完成后,注释是橡胶树基因组学研究的另一项重要工作。
即对基因组中
的基因进行注释,包括确定其位置、结构和功能。
尽管橡胶树基因组组装已经完成,但仍需要进行更多的注释工作,以确定每个基因的准确位置和功能。
2020年,一
项针对橡胶树新品种基因组学的全基因组注释被完成,提出了35,751个基因,其
中2,412个是为本次新组装获得的,具有更好的全程基因组组装质量和注释质量。
2. 橡胶树的基因功能解析
基因的功能解析可以帮助人们更好地了解其在生物体内的功能和作用。
在橡胶
树基因功能的研究中,整合转录组学和蛋白质组学的方法为研究人员提供了重要的信息。
通过对橡胶树基因表达的广泛鉴定,可以更好地理解橡胶树的发育和环境适应等方面表现出的特征。
同时,对橡胶树叶片、发芽芽和根等组织的蛋白质组学分析也有所提高。
3. 橡胶树基因工程
利用基因编辑和转基因技术来改良橡胶树的产量和性质,是橡胶树基因工程领
域的前沿研究。
目前,基于CRISPR/Cas9技术的基因编辑已被广泛应用于橡胶树
的研究中。
一些研究中成功利用CRISPR/Cas9技术研发出了耐盐的橡胶树和增强
产量的橡胶树。
这些成果为橡胶树育种和种植提供了新的思路和途径。
三、展望
当前,橡胶树基因组学已经取得了一些重要的进展,但需要进一步加强对基因
组的研究。
在未来的研究中,应更加注重橡胶树的基因组学研究,特别是注重对其基因的注释和功能解析。
此外,将传统的育种方法与基因工程技术相结合,可在短期内实现橡胶树的改良和优化,从而为橡胶树的可持续发展和利用提供更多的支持。