植物的性别决定
植物的性别与繁殖方式研究

CHAPTER 06
研究前景与展望
植物性别与繁殖方式研究意义
揭示植物生殖机制
通过研究植物性别与繁殖方式,可以深入了解植物生殖系统的运 作机制,为植物生殖生物学提供理论基础。
指导农业生产实践
掌握植物性别与繁殖方式的知识,有助于指导农业生产实践,提 高农作物产量和品质。
保护生物多样性
了解不同植物的性别与繁殖特性,有助于制定针对性的保护措施 ,保护生物多样性。
植物繁殖方式简介
有性繁殖特点及过程
特点
有性繁殖是通过雌、雄配子结合形成 受精卵,进而发育成新个体的繁殖方 式。这种方式可以产生基因重组,增 加后代遗传多样性。
过程
包括花粉的形成和传播、花粉在柱头 上的萌发、花粉管伸长和精子释放、 受精作用以及种子和果实的形成等阶 段。
无性繁殖特点及过程
特点
无性繁殖是不通过生殖细胞结合,而是由母体直接产生新个体的繁殖方式。这 种方式可以保持母本的遗传特性,繁殖速度快。
过程
包括分株、扦插、压条、嫁接等。这些过程都是利用植物的营养器官(根、茎 、叶)来产生新的个体。
繁殖方式对植物生长的影响
有性繁殖的影响
有性繁殖可以增加后代的遗传多样性,提高植物适应环境变 化的能力。但是,由于基因重组的不确定性,有性繁殖也可 能导致后代生长表现不稳定。
无性繁殖的影响
无性繁殖可以保持母本的优良性状,快速繁殖大量新个体。 但是,长期进行无性繁殖可能导致植物遗传多样性降低,增 加病虫害的风险。同时,无性繁殖也可能导致植物体内积累 大量有害物质,影响植物的生长和健康。
未来研究方向及领域,关 注更多非模式植物的性别与繁殖方式 ,揭示它们的生殖机制和适应环境的 策略。
要点二
强化技术应用研究
雌雄异株植物性别鉴定方法概述

性状是由基因决定的,对于性别这个特殊的性状而言,植物雌雄株间表现的差异,无论是性染色体的差异、生理代谢方面的差异、还是同工酶表现的差异,都能从植物基因组DNA上反映该物种性别的本质特征。这种研究方法可在植物的任何发育阶段(只要能提取DNA的植株材料)进行,且鉴定准确、结果重复性好。此方法从分子水平入手,利用DNA分子标记技术,通过分析已知性别植物基因组DNA间的多态性,筛选与其性别相关的分子标记,寻找雌雄植株间的本质差异,为植物的性别鉴定提供理论依据。
关键字:雌雄异株,性别鉴定,评价
1绪论
自然界被子植物大多是雌雄同株,雌雄异株植物仅占6% (Renner等,1995)。由于雌雄异株植物,在植物性别决定尤其是性染色体进化研究中具重要地位(Ioan,2001;Ainsworth,1998),且由于同一树种、不同性别的植物在形态学及生物学特性、经济价值等方面存在一定的差距,林业生产、园林事业以及蔬菜种植中常常希望根据经营目的与利用方向,从中选择栽培某一种性别的植物以满足需要,对植物的早期性别鉴定就显得尤为重要。早在上世纪50年代初波兰学者Bugala首先通过叶色鉴定了欧洲山杨(Populus tremula)的性别(Bugala,1951)。但通过外部形态来鉴定植物性别的方法不够准确,且对某些植物而言,在性器官分化和发育成熟前,其雌雄株的形态差异往往不明显,所以不能作为植物性别早期鉴定的可靠依据。近年来发展了从细胞、生化和分子等不同水平上对雌雄异株植物的性别鉴定进行研究的方法。
一堂“探究植物性别决定方式”的习题课
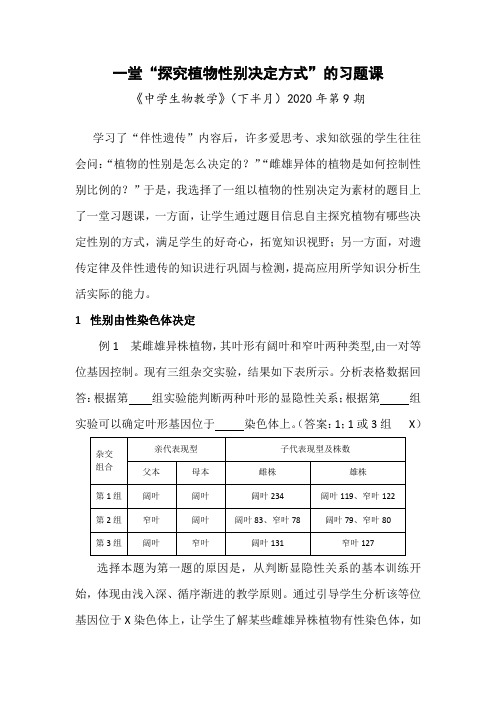
一堂“探究植物性别决定方式”的习题课《中学生物教学》(下半月)2020年第9期学习了“伴性遗传”内容后,许多爱思考、求知欲强的学生往往会问:“植物的性别是怎么决定的?”“雌雄异体的植物是如何控制性别比例的?”于是,我选择了一组以植物的性别决定为素材的题目上了一堂习题课,一方面,让学生通过题目信息自主探究植物有哪些决定性别的方式,满足学生的好奇心,拓宽知识视野;另一方面,对遗传定律及伴性遗传的知识进行巩固与检测,提高应用所学知识分析生活实际的能力。
1性别由性染色体决定例1 某雌雄异株植物,其叶形有阔叶和窄叶两种类型,由一对等位基因控制。
现有三组杂交实验,结果如下表所示。
分析表格数据回答:根据第组实验能判断两种叶形的显隐性关系;根据第组实验可以确定叶形基因位于染色体上。
(答案:1;1或3组X)选择本题为第一题的原因是,从判断显隐性关系的基本训练开始,体现由浅入深、循序渐进的教学原则。
通过引导学生分析该等位基因位于X染色体上,让学生了解某些雌雄异株植物有性染色体,如本题中的植物即为XY型性别决定。
2 性别由单基因决定例2 芦笋是雌雄异株的多年生植物,一次种植可多年采摘其嫩茎作为蔬菜。
芦笋的性别由一对等位基因决定,雄株基因型为AA或Aa、雌株基因型为aa;芦笋自然种群中性别比例为1∶1。
研究人员在芦笋的某些雄株(Aa)上发现了罕见的两性花,让其自花传粉,F1全为雌雄异株、只开单性花(只有雄蕊或雌蕊),且雄株与雌株的比例为3∶1。
请补充从F1雄株中筛选雄性纯合子(AA)的方案:①分别收集不同雄株的花粉给基因型为的雌株授粉,分别得到大量种子。
②播种种子,待其生长发育至开花时,观察植株性别。
若,则提供花粉的雄株为所需的雄性纯合子(AA)。
(答案:①aa ②全为雄株)本题的已知条件为植物性别由常染色体上的一对等位基因控制,雄性为显性性状,雌性为隐性性状。
从F1雄株中筛选纯合子就是验证F1雄株基因型是AA还是Aa,目的是考查学生对基因的分离定律中测交实验法的掌握情况。
植物的性别决定机制

植物的性别决定机制植物的性别决定机制是指植物如何决定自身的性别,即雌雄植株的形成过程。
与动物不同,植物的性别决定并不是由遗传因素所决定,而是受到一系列环境和生理因素的影响。
本文将深入探讨植物的性别决定机制,揭示雌雄植株的形成过程。
一、植物的性别特征在植物中,性别特征主要表现为花部的形态以及生殖器官的结构。
雄性植株的花部通常包含花蕊和雄蕊,而雌性植株的花部则具有花药和子房。
除了这些显性的性别特征外,还有一些植物的性别表现较为隐蔽,需要通过细微的形态差异或分子水平的遗传分析才能确定性别。
二、雌雄异株植物雌雄异株植物是指具有明显的雄性植株和雌性植株的植物。
这些植物通常在不同的植株上发育出雄性和雌性的花部。
这种性别分化主要受到植物激素的调控。
在雄性植株上,大量的雄性激素促使花部发育为雄蕊和花蕊;而在雌性植株上,雄性激素水平较低,使得花部发育为子房和花药。
三、雌雄同株植物雌雄同株植物是指同一株植物上同时存在雄性和雌性的花部。
这种性别决定机制受到复杂的遗传因素和环境因素的共同影响。
在雌雄同株植物中,有些植物呈现两性花,即具有既有雄蕊又有子房的花部。
这种花部结构的形成是由于某些基因对花蕊和子房的发育同时发挥作用。
四、环境因素对性别决定的影响除了遗传因素外,植物的性别决定还受到环境因素的调控。
光照、温度、水分等环境条件的变化都会对植物的性别决定机制产生影响。
例如,一些植物在高温条件下容易形成雌性植株,而在低温条件下则更容易形成雄性植株。
这种环境因素对性别决定的影响使得植物具有性别的可塑性,能够适应不同的环境条件。
五、植物人工性别控制的应用对植物性别决定机制的深入了解,为植物人工性别控制提供了理论基础。
目前,人们常常利用这些性别决定机制来控制植物的性别。
例如,在果树种植过程中,为了提高果实的产量和品质,常常需要控制雌雄植株的比例。
通过合理的栽培管理、灌溉技术以及植物激素的应用,人们可以有效地控制植物的性别。
六、未来的研究方向尽管对于植物的性别决定机制已经有了一定的了解,但仍然存在很多未被揭示的谜团。
生物学中的性别和性别决定机制研究

生物学中的性别和性别决定机制研究性别是生物学中一个重要的概念,它决定了一个个体在进化、繁殖和行为上的差异。
性别的产生和决定机制一直是生物学家们关注和研究的焦点之一。
本文将从植物和动物两个方面介绍性别和性别决定机制的研究进展,并探讨未来的研究方向。
一、植物中的性别和性别决定机制研究在植物中,性别决定机制的研究主要集中在两个方面:雌雄同体植物和雌雄异体植物。
1. 雌雄同体植物的性别决定机制雌雄同体植物是指同一个个体上既有雄蕊又有雌蕊。
性别决定机制的研究发现,这类植物的性别决定主要受到基因和环境的调控。
具体来说,某些基因在植物发育过程中的表达和调控可以决定表达雄性器官还是雌性器官,而外界环境因素如温度和光照等也会对性别的表达产生影响。
2. 雌雄异体植物的性别决定机制雌雄异体植物是指同一物种的雄性个体和雌性个体分别发育成两种不同的形态。
关于雌雄异体植物的性别决定机制,研究发现植物的性染色体在这过程中起到了重要的作用。
比如,一些物种的雄性个体含有XY性染色体,而雌性个体则是XX性染色体。
性染色体决定了植物的性别。
二、动物中的性别和性别决定机制研究在动物中,性别决定机制的研究更加复杂和多样化。
以下将以两个经典的案例来介绍动物中的性别和性别决定机制。
1. 爬行动物中的性别决定机制对于一些爬行动物,如鳄鱼和龟类,性别是由环境温度决定的。
具体而言,鳄鱼和龟类的卵在孵化过程中受到温度的影响,高温下孵化出的是雌性个体,低温则孵化出雄性个体。
这说明环境温度是影响性别决定的重要因素。
2. 哺乳动物中的性别决定机制哺乳动物中,性别决定机制的研究主要聚焦在性染色体和性别基因上。
人类和大多数哺乳动物都拥有两种性染色体,即XX和XY。
在此基础上,性别基因的表达和作用决定了个体的性别发育。
例如,Y染色体上的关键基因SRY编码了性决定区域的蛋白质,它在雄性个体的生殖器官发育过程中起到了关键作用。
三、未来的研究方向尽管对于性别和性别决定机制的研究已经取得了很大进展,但仍然有许多问题有待解决。
附加2-植物的性别

四、雌雄异株植物性别的鉴定
2、通过生理代谢差异鉴别 千年桐(Vernicia mon-tanaLiour)的雌株叶部组织的还原能力大于雄 株。 大麻雄株组织的呼吸速度大于其雌株,而过氧化氢酶的活性也比雌株 高50%~70%。
在银杏、石刁柏中都发现过氧化物酶的活性与植株的性别有关。
四、雌雄异株植物性别的鉴定
5、环境条件与植物性别分化
①、日照长度
长日 雄株 雌株 雄株
大麻
短日 雌株
②、温度
≤10℃
南瓜 (晚上) >10℃
雌花
雄花
二、植物性别决定
5、环境条件与植物性别分化
③、营养
钾肥 雄花 雌花
黄瓜
氮肥
④、外源激素
瓠[hù] 瓜
乙烯利
雌花
三、植物雌雄异株的意义
(许多雌雄异株的植物,其经济价值是不同的)
3、通过化学物质分析鉴别
酚类、玉米素、黄酮、单宁、萜类内酯、内源激素等在不同雌雄异株植物中 含量会有相应的差异。
4、通过同工酶差异鉴别 过氧化物酶及多酚氧化酶、黄瓜的吲哚乙酸氧化酶、多酚氧化酶以及 超氧 化物歧化酶同工酶都可以作为其雌性早期鉴定的生化标记。 分子酶学“一个基因一个同工酶亚基”的理论。 张立平等,和晁无疾在对葡萄雌雄株的超氧化物歧化酶同工酶进行分析,对 葡萄植株进行性别鉴定的结果不一致。
四、雌雄异株植物性别的鉴定
8、化学药剂处理鉴别
根据叶片所含酚类物质与不同化学试剂反应所产生的稳定颜色变化,
鉴别了银杏、黑杨、旱柳、沙棘、杜仲及猕猴桃的雌雄株。
优点:快捷、廉价和实用 缺点:易受环境因素及植物生长发育不同时期的影响,对鉴别结果 的可靠性有一定的干扰。
植物的性别分化

3. 相关技术
(1)矮壮素(CCC)是GA合成的抑制剂。以101MCCC溶液喷洒或浇灌黄瓜幼苗,可使植株 完全雌性化。 (2)一些无机离子,如Ag+(常用AgNO3)和 Co2+(常用CoCl2)能在一些植物的雌株中 诱导出雄花。
Thank you !
二. 环境条件的影响
基本 条件
环境 条件
日照
植物 激素
1. 一般说来,充足的氮素营养,较高的空气和土壤
温度,较低的气温(特别是夜间低温),蓝光, 种子播前冷处理等,有利于雌性分化;高温、红 光等因子则促进雄花分化。
2. 日照长度的影响因植物光周期类型而异;一般短
日照促进短日植物(SDP)多开雌花,使长日植 物(LDP)多开雄花;长日照的作用则相反。
13级生科二班
梁月圆
植物性别的分化
一. 性别分化的简要机理
二. 环境条件的影响雄同 花植物 雌雄同株 异花植物 雌雄异 株植物 桃、苹果、 草莓...... 玉米、瓜 类...... 大麻、银 杏......
2.性别分化的机理:
一般认为雌、雄同株植物形成雌、雄器官 的基因均已在全能的分化细胞中预先编码,环 境因子或化学药剂只起着一个阻抑或脱阻抑的 作用。
数大大下降,雌花数显著提高。一氧化碳处
理不仅可改变雌雄花的比例,而且可改变雌
雄花出现的顺序,降低了雌花着生的节位,
可使黄瓜提前长成上市。
三. 鉴定及实践意义
1. 性别鉴定
(1) 开花顺序
(2) 呼吸速率
2. 实践意义
以种子和果实为收获对象则需要大量的
雌花或雌株,而有时为了其他目的,就更
欢迎雄株,例如以纤维为收获物的大麻, 以雄株为优,因其纤维拉力较强,为了得 到银杏种子,宜多种雌株,而如用银杏作 行道树,则又以雄株为佳。
植物和动物的性别表达和遗传方面的研究

植物和动物的性别表达和遗传方面的研究植物和动物作为自然界中不可或缺的组成部分,其性别表达和遗传方面的研究也一直备受科学家的关注。
通过对植物和动物性别表达和遗传的深入研究,可以更好地了解性别演化的规律和机制,为保育和利用生物资源提供重要的理论基础。
一、植物的性别表达和遗传方面的研究1.性别决定系统植物的性别决定系统可以分为两类:单倍体和二倍体。
单倍体的性别决定系统主要是通过雌雄配子体现,如大多数苔藓类植物。
而二倍体植物的性别决定系统则主要是通过染色体和基因的调控来实现。
比如草本植物中,雌雄性和单性花的形成都是由基因调控的。
2.雌雄同体植物的性别转换雌雄同体植物是指植物的雌雄生殖器官同时存在于同一个个体中。
而这些个体的性别通常在发育过程中由外界环境和内源性调节因素来决定。
比如,一些土豆、黄瓜等作物种类中,灰姑娘因素可以影响植物体内雄性激素合成的过程,从而调节植物的性别转换。
3.基因调控植物的性别遗传也是通过基因调控来实现。
其中,一些基因编码了特定的转录因子,可以影响植物的雄蕊、雌蕊和萼片等性状的发育。
比如,阿拉伯芥中的MS1基因编码转录因子,可以影响花的雄蕊的形成和发育。
二、动物的性别表达和遗传方面的研究1.性别决定系统动物的性别决定系统也可以分为两类:单倍体和二倍体。
单倍体动物的性别决定主要是通过雌雄配子体现,如自创动物。
而大多数哺乳动物和鸟类的性别决定则主要是通过染色体和基因的调控来实现。
2.性别基因哺乳动物中,性别基因通常分为X染色体和Y染色体。
多数情况下,XY染色体系统使雄性发育而XX染色体系统使雌性发育。
但是,也有一些哺乳动物具有相反的CBOQ基因,如野生喜马拉雅兔,它们中的雌性拥有X和Y染色体,而雄性只有X染色体。
3.遗传调控动物的性别遗传也是由基因调控来实现的。
其中,许多性别相关因子编码了雄性或雌性激素水平的调节因子,能够影响生殖器官和性腺的发育。
比如雄性胚胎中的SRY基因编码一种具有转录调节活性的蛋白质,它被认为是哺乳动物男性发育的核心基因。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
植物的性别决定张 霞 王绍明(新疆石河子大学生物科学系新疆石河子832003) 许多高等植物是雌雄同株类型,并无明显的性染色体机制存在。
同株上的雌雄性器官,显然是在发育后期通过细胞分化形成的,没有什么特殊的遗传基础,但在少数雌雄异株的植物内,也有与动物类似的性别决定机制。
性器官随发育条件而形成,因而实际上受环境影响而改变的情况较常发生。
1 基因决定性别1.1 一对等位基因决定性别 一些植物的性别差异并不是由整个性染色体所决定,而是由一对等位基因控制。
如石刁柏(A s p ar agus of f icinalis)在正常情况下是雌雄异株植物,雌株和雄株呈1∶1之比。
但有时在一个群体中可以见到雌雄同株的个体,它的雌花中有退化的雄蕊,而雄花中有退化而无功能的雌蕊柱头,甚至还可以产生成活的种子。
如将这些种子种下,会出现3/4雄株和1/4雌株,表明是一对基因分离的结果,如把分离出的雄株与正常雌株再行杂交,其中有大约1/3雄株的杂交后代完全为雄株,其余2/3杂交后代中雌株∶雄株= 1∶1,可见雄性基因对雌性基因呈完全显性作用。
在群体中,雄株大都处于杂合状态。
1.2 一组复等位基因决定性别 葫芦科的一种植物喷瓜(Ecballium elaterium),其性别是由1个基因座位的3个复等位基因所控制,即a D、a+和a d。
a D对a+和a d为显性,a+对a d为显性。
a D决定雄性,a+决定雌雄同株,a d决定雌性,它们的不同组合决定植物的性别。
a D a+和a D a d 为雄株,a+a+和a+a d为雌雄同株,a d a d为雌株。
不存在纯合的a D a D,表明a D可能有显性纯合致死作用。
1.3 两对等位基因决定性别 玉米(Zea may s)是雌雄同株植物,雄花为圆锥花序,生长在植物的顶端,雌花为穗状花序,生长在植株中部的叶腋间。
正常的雌雄同株玉米雌花序是由显性基因Ba控制,雄花序是显性基因T s控制,基因型为BaBaT sT s。
当基因Ba突变为ba时,基因型baba就会使植株不能长出雌花序而成为雄株,因此,雄株的基因型为babaT sT s或babaT sts。
当基因型T s突变为ts时,基因型t sts就会使植株的雄花序变成雌花序,可以通过受精结出种子,成为雌株,基因型为BaBatst s或Babat sts。
当ba和ts分别为隐性纯合时,即babatst s时,雄花序上长出雌穗,变成雌株。
2 染色体决定性别2.1 雄异配子型 有些被子植物(大麻属Cannabis,草属Humulus,菠菜属Spinacia,麦瓶草属Silene)和许多苔藓植物可以从细胞学上鉴别出性别染色体。
雌株为同型性染色体,雄株为异型性染色体。
2.1.1 由X和Y染色体之间的平衡决定性别的XY 型 例如一种藓Sp haer ocar p us donellii的孢子体在减数分裂时,大的X染色体和小的Y染色体各自分离到一半的孢子中。
其中有X染色体的孢子长成雌性配子体,有Y染色体的孢子长成雄性的配子体。
在二倍体的白麦瓶草(Silene album)中,雌株有两个X染色体,雄株是1对X-Y染色体,Y明显地比X大一些。
有Y染色体时,一般是雄性;没有Y,则是雌性,并且与X染色体的数目和常染色体的套数无关。
唯一的例外是1个具有4个X染色体和1个Y染色体的四倍体植株,它通常是雌雄同株的。
用X射线使Y染色体断裂,形成Y染色体有部分缺失的植株。
对这些植株的研究表明,Y染色体的一部分含有控制花药发育的基因,而另一部分具有抑制雌性性状发育的基因,这样,麦瓶草很象人和其它哺乳动物,其性别由X和Y染色体之间的平衡所决定。
雌雄异株的女娄菜(M elandr ium album)也属于XY 型性决定,Y染色体携带决定雄性的基因,具有决定雄性的作用,因此,合子含有Y染色体时,它就将发育成雄株。
W easterg and(1948)通过染色体缺失的观察证明了女娄菜Y染色体上有4个在性别决定中具有不同作用的区段,Ⅰ为雌性抑制区,可使带有Y染色体的合子发育成雄株,缺失这个区段的植株是两性的;Ⅱ为促进雄性发育区,当这个区段缺失时,就产生雌性植株;Ⅲ为雄性可育区,携带雄性可育基因,这个区段缺失时,形成雄性不育株;Ⅳ为X染色体的同源区。
在X染色体上有与Y的非同源部分,并且决定雌株的基因大部分在X 染色体上。
2.1.2 由X染色体与常染体之间的平衡决定的XY型 酸模草(R umex acetosa)及其近缘种内,在人工诱导的三倍体和四倍体后代中,有一些具有几种染色体数目的植株,这些植株象果蝇一样性别是由X染色体与常染色体之间的平衡决定的,与Y染色体的关系很小。
具有1X∶1A(A=6个常染色体)比率的植株,无论Y染色体是否存在,都是雌性。
而具有1X∶2A或3A比率的植株是雄性。
一些中间比率(3A+2X、4A+3X、6A+ 4X,有或者没有Y都一样)的植株则是中间性别。
2.1.3 属于XO型的雄异配子型 苦草(V allisner ia sp ir alis)、秦椒(Zanthox y lum p ip eritum)和薯蓣属中的D ioscorea sinuata均属于X O型性决定方式,即雌性为同型的两个X染色体,雄性为一个X染色体。
2.2 雌异配子型 在被子植物中,已知有草莓属中的—24—生 物 学 通 报 2001年第36卷第1期Rh血型的基因、抗血清及表现型的命名高志国(河北师范大学生命科学学院河北石家庄050016)王长春(河北师范大学成人教育学院河北石家庄050016) Rh血型的名称取自恒河猴学名Rhesus macaque的前两个字母,因为在最初探索新血型物质的过程中发现,当把恒河猴的红细胞反复地注入家兔及豚鼠体内时,家兔和豚鼠的血清可使恒河猴的红细胞凝集,说明恒河猴的红细胞上含有某种抗原,遂命名为R h抗原。
以后又发现经免疫的家兔或豚鼠的血清还能使大部分人的红细胞凝集,表明这些人的红细胞上也含有R h抗原,并称之为Rh阳性血型,还有一些人的红细胞不被这种抗血清所凝集,便称为Rh阴性血型,由R h抗原所决定的这一血型系统就称为R h血型。
后来,随着对各种输血反应、新生儿溶血症以及血清学试验的研究进展,关于Rh血型的免疫学及遗传学知识也不断丰富,现在已经知道,控制Rh抗原的基因(或基因组合)主要有8种,它们均位于第1对染色体上,并已制备出5种R h血型的标准抗血清。
但是,由于在理论解释上的不同,对Rh血型的基因、抗血清及表现型的命名也不统一。
现就3种常用命名法,即“R h-Hr”命名法、“CDE”命名法和“序号”命名法叙述如下。
Rh-Hr命名法认为,Rh血型决定于单一染色体位点上的8个复等位基因,它们分别为:rh、Rh1、R h2、Rh0、rh′、r h″、Rh z、rh y。
每个等位基因控制着一种红细胞抗原(或凝集原)的产生,每种红细胞抗原可包括多个抗原决定簇或抗原因子,后者具有各自不同的抗体特异性,即能够诱导产生不同抗体。
Rh-Hr命名法将5种标准抗血清分别命名为R h0、rh′、rh″、hr′、hr″,带有每种抗原的红细胞可被至少两种抗血清所凝集。
例如,带有rh 抗原的红细胞可被hr′和hr″抗血清所凝集;带有Rh1抗原的红细胞可被Rh0、rh′和hr″抗血清所凝集;带有Rh2抗原的红细胞可被R h0、hr′和r h″抗血清所凝集;带有Rh0抗原的红细胞可被R h0、hr′和hr″抗血清所凝集;带有r h′抗原的红细胞可被r h′和hr″抗血清所凝集;带有rh″抗原的红细胞可被hr′和rh″抗血清所凝集;带有R h z 抗原的红细胞可被R h0、r h′和r h″抗血精所凝集;带有rh y抗原的红细胞可被rh′和r h″抗血精所凝集。
表1 在3种命名法中Rh基因的名称Rh-Hr命名法CDE命名法序号命名法rh cde R-1,-2,-3,4,5Rh1CDe R1,2,-3,-4,5Rh2cCE R1,-2,3,4,-5Rh0cDe R1,-2,3,4,5rh′Cde R-1,2,-3,-4,5rh″cdE R-1,-2,3,4,-5Rh z CDE R1,2,3,-4,-5rh y CdE R-1,2,3,-4,-5以上基因相互组合,可产生36种基因型 CDE命名法认为,控制Rh抗原的基因位于紧密连锁的3个染色体位点上,每个位点上各有1对等位基因,分别为Cc、Dd、Ee,每个人可具有各种组合中的任何3对基因,同1条染色本上可有8种基因组合,即cde、CDe、cD E、cDe、Cde、cdE、CDE、CdE。
按此理论,3对等位基因应控制6种抗原,但实际上仅发现C、D、E、c、e5种抗原,而未发现d抗原。
因此认为基因d是不表达的“静止基因”,所以在红细胞表面没有d抗原。
在发现的5种抗原中,以D抗原的抗原性最强,因而将红细Fr agar ia elatior和委陵菜属的金露梅(Potentilla f ruticosa)等,它们的雌株为ZW,雄株为ZZ。
3 环境条件决定性别植物中由环境条件决定性别的例子很多,这类植物也没有决定性别特殊的遗传基础,性别的分化决定于所处的内外在环境条件。
瓜类是雌雄异花而同株的植物。
已知对黄瓜早期施用多量氮肥可以促进雌花形成;缩短每日光照时间也可增加雌花;CO2有促进雌花的作用。
在南瓜中,如温度降低到10℃左右,特别在晚上,有利于雌花形成。
如果晚间低温与白天8小时光照相结合,可使雌花占绝对优势。
在雌雄异株的大麻中,由于环境条件的影响,有时可以引起性返转。
由环境决定性别的生物并不是自身没有决定性别的遗传基础或基因,恰恰相反,这类生物很可能包含了决定雌性和雄性的两套基因,因而具有向两种性别分化的可能性。
至于哪套基因能够正常地发挥作用,取决于一定的内、外在环境条件。
4 参考文献 1 李竟雄,宋同明.植物细胞遗传学.北京:科学出版社, 1997. 2 黎中明,林文君.细胞遗传学.成都:四川大学出版社, 1988.(BH)—25—2001年第36卷第1期 生 物 学 通 报。