04-抗Bt棉棉铃虫幼虫Bt受体氨肽酶N_APN2_基因克隆
农大棉8号基因组特点

农大棉8号基因组特点
1. 抗虫基因的导入:农大棉8号品种中导入了Bt基因,该基因来自于一种称为苏云金芽胞杆菌(Bacillus thuringiensis,简称Bt)的细菌。
Bt基因经过转基因技术导入棉花中,使得棉花表达Bt抗虫蛋白(Cry蛋白),从而使其对一些棉花害虫如棉铃虫、斜纹夜蛾等具有显著的抗虫效果。
2.抗草生基因的导入:农大棉8号品种中还导入了抗草生基因
(JH13-2基因),该基因通过转录后基因沉默技术(RNAi)制备。
抗草生基因通过抑制棉花中的一些天然生长因子的合成及转运,使棉花在无抗草剂喷洒的情况下能够有效抵御各类草本杂草的竞争。
3.高效果蛋白转录:农大棉8号品种中的转基因功能基因组拥有高效的转录和蛋白表达能力。
这使得该品种能够在相对较短的时间内产生足够的抗虫蛋白和抗草生蛋白,从而保证了抗虫抗草生特性的高效性。
4.优异品质表现:农大棉8号品种在基因组特点上的改良并未影响其优异的品质表现。
该品种依然保持了传统棉花的优质纤维特性,如纤维强度和长度的稳定性,同时还具有较高的纺织性能和良好的纤维度等。
5.遗传稳定性:农大棉8号品种的基因组特点的改良通过遗传工程技术进行,并经过严格的鉴定和安全性评估。
研究表明,农大棉8号品种的转基因特性在连续繁殖后会保持稳定,不会对环境和人类健康产生负面影响。
总结起来,农大棉8号品种的基因组特点主要包括抗虫特性、抗草生特性、高效果蛋白转录、优异品质表现以及遗传稳定性。
这些特点使得农大棉8号品种成为一种具有抗虫抗草生能力、产量稳定且具备优质纤维的
转基因棉花品种。
然而,值得注意的是,转基因棉花的安全性和环境影响仍需要进一步的研究和评估。
【国家自然科学基金】_棉铃虫_基金支持热词逐年推荐_【万方软件创新助手】_20140801
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82
发生趋势 双荧光素酶报告基因系统 双向电泳 原核表达 印楝 凋亡抑制基因(iap) 克隆 信息素结合蛋白 中肠刷状缘膜囊泡 中肠 vcath基因 rt-pcr pcr-rflp p49基因 p35基因 optiprep nimbin iso-dalt ipg-dalt iap基因家族 hearnpv cyp9a17v2基因 cyp9a17v2 cry1ac chib bt菌素 bmkit基因 bac-to-bac栽体系统 acmnpv 38k基因
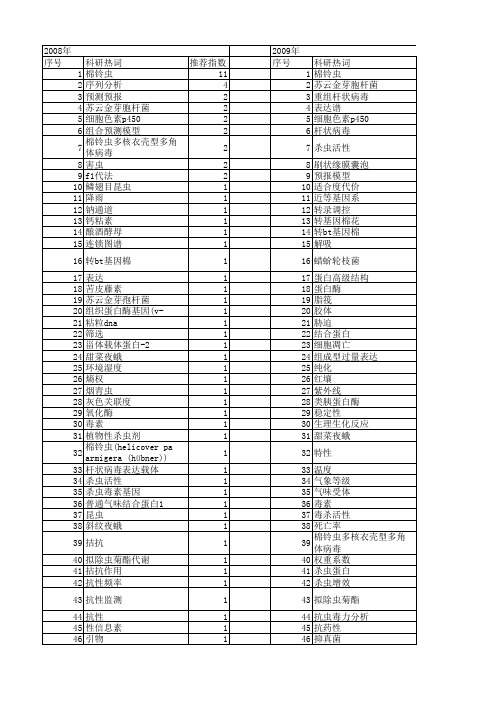
科研热词 推荐指数 棉铃虫 11 序列分析 4 预测预报 2 苏云金芽胞杆菌 2 细胞色素p450 2 组合预测模型 2 棉铃虫多核衣壳型多角体病毒 2 害虫 2 f1代法 2 鳞翅目昆虫 1 降雨 1 钠通道 1 钙粘素 1 酿酒酵母 1 连锁图谱 1 转bt基因棉 1 表达 1 苦皮藤素 1 苏云金芽孢杆菌 1 组织蛋白酶基因(v-cath) 1 粘粒dna 1 筛选 1 甾体载体蛋白-2 1 甜菜夜蛾 1 环境湿度 1 熵权 1 烟青虫 1 灰色关联度 1 氧化酶 1 毒素 1 植物性杀虫剂 1 棉铃虫(helicover pa armigera (hübner)) 1 杆状病毒表达载体 1 杀虫活性 1 杀虫毒素基因 1 普通气味结合蛋白1 1 昆虫 1 斜纹夜蛾 1 拮抗 1 拟除虫菊酯代谢 1 拈抗作用 1 抗性频率 1 抗性监测 1 抗性 1 性信息素 1 引物 1 异源表达 1 害虫综合防治 1 实验设计 1 天敌 1 多核衣壳型多角体病毒 1 基因组步移 1
Bt抗性和敏感棉铃虫钙黏蛋白及氨肽酶N1表达量的比较

摘要
棉铃 虫体 内的钙黏蛋 白( AD 和氨肽酶 N1 A N1是 B 的主要作用靶标 , C ) (P ) t 两种 受体蛋 白的变化也是棉铃 虫
对 B 产 生抗性的主要原 因。本文利用实时荧光定量 P R技术 比较 了 B 抗性和敏感棉铃 虫 C t C t AD和 A N1 P 的表达 量 变化 , 分析受体蛋 白表 达量变化 与抗性 的关 系。结果表 明, 与敏 感品 系相 比, 抗性 品 系棉铃 虫 C D和 A N1的 A P 表达量 明显 降低 , 、 12龄幼 虫的差异达到极显著 。在敏 感品 系中, A C D和 A N1的表达 量随龄期的增加呈现“ 字 P V”
Zh n o , W e ihe z a g Ta i z n , Zh n l J a g Li , Lu q o g , Li n me 。 i in a g Ge i , Ya g La g ’ n n 。
( .C l g 厂 r utr ,Gu nx nvri ,N n i 50 0 ,C i 1 ol eD Agi l e e c u a g i i st a nn U e y g 3 0 4 hn a; 2 tt Ke a oaoyfr .Sae yL b rtr o
Co p rs n o h a h rn a d a i o e t a e N1 e p e so m a io ft e c d e i n m n p p i s - x r s in d lv l e we n Btr ss a ta d Bts s e tb e c to e e s b t e -e it n n - u c p i l o t n - b lwo m e io e p r g r o l r H lc v r a a mi e a
高考生物《基因工程知识点》总汇

高考生物《基因工程知识点》总汇1、基因工程的先导是?艾弗里等人的工作证明了DNA可以从一种生物个体转移到另一种生物个体2、不同生物的基因为什么可以连接在一起?因为所有生物的DNA基本结构是相同的3、真核生物的基因为什么可以在原核生物体内表达?(或者原核生物的基因为什么可以在真核生物体内表达?)所有生物共用一套密码子4、基因工程育种的原理是什么?具有什么优点?原理:基因重组优点:打破了生殖隔离,定向改造生物的性状5、与DNA有关的酶的比较6、特定的核苷酸序列,并在特定的位点上进行切割7、限制酶不切割自身DNA的原因是什么?原核生物DNA分子中不存在该酶的识别序列或识别序列已经被修饰。
8、DNA连接酶可以连接什么样的末端?①同一种限制酶切割形成的相同的黏性末端②两种不同限制酶切割后形成的相同黏性末端③任意的两个平末端9、如何防止载体或目的基因的黏性末端自己连接即所谓“环化”?可用不同的限制酶分别处理含目的基因的DNA和载体,使目的基因两侧及载体上各自具有两个不同的黏性末端。
10、载体需具备的条件及其作用11、基因工程的基本操作步骤是哪四步?目的基因的获取;基因表达载体的构建;将目的基因导入受体细胞;目的基因的检测与鉴定12、目的基因的获取方法有哪些?三种方法都需要模板吗?①从基因文库中获取目的基因②利用PCR技术扩增目的基因③通过化学方法人工合成前两种需要模板,从基因文库中寻找目的基因时需要用DNA探针利用DNA分子杂交的方法找到目的基因;化学方法人工合成不需要模板,只要知道核苷酸序列就行,这是一个纯粹的化学反应13、CDNA文库和基因组文库的区别?cDNA是指以mRNA为模板,在逆转录酶的作用下形成的互补DNA。
以细胞的全部mRNA 逆转录合成的cDNA组成的重组克隆群体成为cDNA文库。
cDNA文库只包含表达的基因,并且逆转录得来的基因缺乏内含子和启动子、终止子等调控序列基因组文库指的是将某种生物的基因组DNA切割成一定大小的片段,并与合适的载体重组后导入宿主细胞,进行克隆得到的所有重组体内的基因组DNA片段的集合,它包含了该生物的所有基因。
转基因抗虫棉

转基因抗虫棉的研究进展摘要:综述了转基因抗虫棉的研究进展,包括抗虫基因的研究、载体构建技术的研究、转化技术的研究及存在的问题等,并展望了转基因抗虫棉未来发展前景。
关键词:转基因抗虫棉花研究进展引言棉花生长周期长、虫害多,造成的损失非常严重。
据统计,在转基因抗虫棉商品化之前,全球每年用于防治棉花虫害的费用高达20亿美元,约占所有农作物防虫费用的四分之一。
[1]传统的化学农药防治棉铃虫不仅费用高,且已引发了棉虫的抗药性,同时化学杀虫剂的过量使用也带来了环境污染的问题,而转基因植物所产生的杀虫蛋白主要是通过抑制害虫消化等生理功能而达到抗虫的目的。
与施药防治棉田害虫相比,转基因技术具有较多优势:不会在土壤和地下水中造成残留;不会被雨水冲刷流失;对非靶标生物无毒性;保护作用无盲区;减少农药及用工投入[2]等。
雪花凝集素(Gulanthus nivalis agglutinin gene,GNA)是第一个转入重要作物、并对刺吸式口器害虫有抗性的基因,转GNA的水稻可降低害虫的存活率,阻止害虫的发育[3]。
另外烟草阴离子过氧化物酶[4]、昆虫几丁质酶基因[5]也被用于抗虫基因工程的研究。
迄今为止在棉花抗虫基因工程研究领域,最成功的例子是苏云金芽孢杆菌Bt杀虫基因的应用,其次是蛋白酶抑制剂基因。
另外,凝集素、α-淀粉酶抑制剂、胆固醇氧化酶等转基因抗虫植物的研究也取得了进展,所以利用基因工程技术培育转基因抗虫棉受到了各国的高度重视。
自1996年商品化种植转基因作物开始,全球转基因植物的种植面积已由1996年的170万hm2猛增到2008年的1.25亿hm2,增长了73倍,2008年全球市场价值已达75亿美元,约占全球商业种子市场的22%,其市场价值优势明显,转基因产业得到了蓬勃发展,尤其在发展中国家。
印度Bt棉2002年引入,连年种植面积快速增加,至2008年达760万hm2,产量翻番,曾经是全球棉花产量很低的国家,现已成为棉花出口国。
2023北京高三二模生物汇编:基因工程章节综合

2023北京高三二模生物汇编基因工程章节综合A.①过程的目的基因是抑制EPSP合酶的基因4.(2023·北京西城·统考二模)医生可利用分子生物学技术检测受检人是否携带HIV。
下列叙述错误的是()A.可根据HIV的RNA序列合成小段DNA作为引物B.血液样品中HIV的RNA经逆转录后进行PCR检测C.可通过抗原-抗体杂交技术检测血液样品中HIV抗原D.与检测抗原、核酸相比,检测抗体能更早诊断HIV感染5.(2023·北京昌平·统考二模)转座子是基因组中可移动的DNA片段,玉米Ac转座子能编码转座酶而自主转座,Ds转座元件只有与Ac转座子同时存在时,才能从原位点切离并插入到新位点中。
研究者利用玉米转座子系统构建烟草突变体,下列叙述错误的是()A.推测Ds转座元件不具有编码转座酶功能B.可构建同时含有Ac/Ds的基因表达载体C.利用农杆菌转化法将基因表达载体导入烟草细胞D.Ds与其被插入的基因间发生基因重组6.(2023·北京朝阳·统考二模)下列生物学实验中,观察实验现象时需借助仪器的是()A.利用琼脂糖凝胶电泳鉴定PCR产物B.提取和分离菠菜叶片中光合色素C.利用酚红指示剂鉴定尿素分解菌D.用酸性重铬酸钾判断酵母菌呼吸方式7.(2023·北京朝阳·统考二模)利用蛋白质工程改造源于溶珊瑚弧菌的几丁质酶:保留该酶N端,在C端增加陆生真菌几丁质结合保守域,从而增加酶与底物的结合能力。
改造后的酶活性显著高于市售几丁质酶。
相关叙述错误的是()A.几丁质是海洋中含量丰富的多糖之一B.溶珊瑚弧菌通过分泌几丁质酶获取氮源C.直接操作对象是溶珊瑚弧菌的几丁质酶基因D.表达改造后的几丁质酶需要借助于受体细胞8.(2023·北京昌平·统考二模)“筛选”是生物学中常用的技术手段,下列叙述错误的是()A.用特定的选择培养基筛选杂交瘤细胞B.通过连续自交的方式筛选隐性纯合子植株C.可利用质粒上的标记基因筛选重组DNA分子D.可利用两种原生质体颜色差异筛选融合细胞9.(2023·北京昌平·统考二模)为确认鸡的肝脏和黑藻叶片存在的共性,相关实验和方法正确的是()A.利用光学显微镜观察细胞膜的结构B.利用双缩脲试剂检测是否含有还原糖C.利用二苯胺试剂鉴定是否存在染色体(2)为协调菌体生长与产物生产之间的关系,将构建好的重组质粒转入经______处理后的枯草芽孢杆菌(D 菌),在含______的培养基上筛选,得到枯草芽孢杆菌E(E菌)。
【国家自然科学基金】_氨肽酶_基金支持热词逐年推荐_【万方软件创新助手】_20140803
推荐指数 2 2 2 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2011年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34
2011年 科研热词 氨肽酶n 表达量 亚洲玉米螟 钙黏蛋白 酶活性 设计 肿瘤靶向治疗 结合能力 系统进化 氨肽酶n抑制剂 氨肽酶n同工酶 氨肽酶n1 森林类型 棉铃虫 昆虫 新生血管 抗性 微生物生物量 小菜蛾 基因功能 土壤可溶性有机质 哌啶二酮衍生物 合成 受体 克隆技巧 亲缘关系 ngr isodgr cry1ab crvlab bt毒素 bt cry毒素 bt apn3 推荐指数 3 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2014年 序号 1 2 3 4 5 6 7 8 9 10 11 12
2014年 科研热词 酪蛋白 胰岛素 结直肠癌 游离氨基酸 淋巴结转移 泌乳奶牛 体外培养 乳腺组织 irap glut4 dukes分期 2型糖尿病 推荐指数 1 1 1 1 1 1 1 1 1 1 1 1
2012年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
推荐指数 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29
人教版高中生物学选择性必修3生物技术与工程精品课件 第3章 基因工程 第2节 基因工程的基本操作程序
酶的主要原因是
。
答案:(1)解旋酶 加热至90 ℃以上 氢键
(2)Taq DNA聚合酶热稳定性高,而大肠杆菌DNA聚合酶在高温下会失活
知识点一
知识点二
知识点三
知识点四
解析:(1)体内进行DNA复制时,需用解旋酶破坏氢键使双链DNA解旋,而 利用PCR技术扩增目的基因时,是借助高温加热至90 ℃以上使DNA变性。 (2)PCR反应体系中进行互补链的合成时,温度需控制在72 ℃左右,则应使 用耐高温的DNA聚合酶,即Taq DNA聚合酶,而不能使用大肠杆菌DNA聚 合酶(高温下会失活)。
知识点一
知识点二
知识点三
知识点四
(2)在PCR技术中,为什么不直接加入解旋酶对DNA进行解旋?为什么要 求所加入的DNA聚合酶具有耐高温的特性?子链的合成为什么需要引物?
提示:DNA解旋酶既可以打开双链模板DNA,又可以打开模板DNA与引 物结合的部分,会导致复制与解旋不断地重复,达不到扩增效果。因 此,PCR反应中利用高温代替解旋酶使DNA解旋。PCR反应需要高温变性, 因此需要耐高温的DNA聚合酶。DNA聚合酶只能将单个的脱氧核苷酸连 接到已有的短单链核酸上,因此需要一段已知序列的短单链核酸作为引物。
知识点一
知识点二
知识点三
知识点四
3.启动子、终止子与起始密码子、终止密码子的区别
项目 本质 位置
作用
启动子
终止子
起始密码子 终止密码子
有特殊序列结构 有特殊序列结构 mRNA上三个 mRNA上三个
的DNA片段
的DNA片段 相邻碱基
相邻碱基
基因上游
基因下游
mRNA首端 mRNA尾端
RNA聚合酶识别
和结合的部位,驱 终止转录
转基因抗虫棉的培育过程
转基因抗虫棉的培育过程随着全球人口的增长和农业需求的增加,农作物病虫害问题日益突出,传统农药防治手段面临着限制和挑战。
为了解决这一问题,科学家们研究出了转基因技术,通过转基因技术培育出抗虫棉,从而提高棉花产量和质量,降低农药使用量,减少环境污染。
转基因抗虫棉的培育过程主要包括以下几个步骤:1.选取抗虫基因:选择具有抗虫特性的基因作为转基因抗虫棉的材料。
这些基因可以通过自然产生的抗虫物质,在转基因抗虫棉中发挥相同的功能。
常用的抗虫基因包括Bt基因、Cry基因等。
2.构建转基因质粒:将选取的抗虫基因与转基因质粒载体进行连接。
转基因质粒是一种具有特定功能的DNA分子,可以将选取的基因导入到棉花细胞中。
常用的转基因质粒载体有pBI121、pCAMBIA1300等。
3.转化棉花胚性培养:将转基因质粒导入到棉花胚性组织中。
首先,通过切割幼嫩的棉花胚珠,得到碎片组织。
然后,将转基因质粒通过基因枪等手段导入到棉花胚性组织内。
转基因质粒会与棉花细胞的染色体融合,形成转基因组织。
4.再生植株培养:将转基因组织培养在含有适宜营养物质的培养基上进行培养。
转基因组织经过分裂和分化,最终发育为幼苗。
经过再生培养,可以得到许多转基因植株。
5.筛选抗虫转基因植株:通过PCR等分子生物学技术,筛选出带有抗虫基因的转基因棉花植株。
这些植株具有抗虫特性,可以抵抗常见的棉铃虫等害虫的攻击。
6.比较试验:将抗虫转基因植株与常规棉花品种进行比较试验。
比较试验包括生长特性、产量、纤维品质等方面的比较。
通过比较试验,可以评估抗虫转基因棉花的效果和优势。
7.田间试验:在田间条件下进行转基因抗虫棉的试验种植。
通过观察和分析抗虫转基因棉花在实际生长环境下的表现,可以更全面地评估其抗虫效果和农艺特性。
8.安全评估:进行对抗虫转基因棉花的安全评估,包括食品安全、环境影响等方面。
确保转基因抗虫棉花对人体和环境的影响符合安全标准。
9.市场推广:在安全评估通过后,将转基因抗虫棉花推广到市场。
棉铃虫载脂蛋白受体基因克隆及转录分析
棉铃虫载脂蛋白受体基因克隆及转录分析
刘香亚;杨蕊弘;张晶;彭英传;肖海军;张万娜
【期刊名称】《植物保护》
【年(卷),期】2023(49)1
【摘要】载脂蛋白受体(lipophorin receptor, LpR)在昆虫体内脂质转运中发挥重要作用。
为了明确棉铃虫载脂蛋白受体基因在其卵巢发育及饥饿胁迫中的功能,本研究克隆了棉铃虫HaLpR基因的cDNA序列(GenBank登录号KU355886.1),全长3 117 bp,开放阅读框为2 643 bp,编码880个氨基酸;其氨基酸序列包含典型的低密度脂蛋白受体结构域,与其他鳞翅目昆虫的LpR相似度>80%。
荧光定量PCR 检测结果显示,HaLpR在雌蛾的卵巢和脂肪体组织中相对转录水平最高;在雌蛾卵子发生期均有转录,在雌蛾羽化后3 d表达量达到最高峰;饥饿处理会抑制HaLpR在雌蛾卵巢和脂肪体中的转录。
研究结果有助于解析HaLpR参与棉铃虫卵巢发育过程中的脂质转运以及应对饥饿胁迫的作用。
【总页数】9页(P213-221)
【作者】刘香亚;杨蕊弘;张晶;彭英传;肖海军;张万娜
【作者单位】江西农业大学;北京林业大学
【正文语种】中文
【中图分类】Q963
【相关文献】
1.棉铃虫嗅觉受体基因的克隆及组织特异性表达
2.水牛促卵泡素受体(FSHR)基因启动子克隆、生物信息学分析及转录活性检测
3.棉铃虫离子型受体基因HarmIR8a的克隆及表达定位
4.β3肾上腺素受体调节肝细胞核因子活性促进肝细胞载脂蛋白A-Ⅰ基因的转录
5.抗Bt棉棉铃虫幼虫Bt受体氨肽酶N(APN2)基因克隆
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
抗Bt 棉棉铃虫幼虫Bt 受体氨肽酶N(APN 2)基因克隆3苏建亚 周晓梅 沈晋良33(南京农业大学农业部病虫监测与治理重点开放实验室农药科学系 南京 210095)摘要 通过对Bt 棉抗性和敏感棉铃虫幼虫中肠氨肽酶N 的克隆和测序,鉴定了氨肽酶N 基因家族的1个成员Haapn2,其cDNA 序列具有3209个核苷酸,含有3096bp 的开放阅读框,编码产生1032个氨基酸的蛋白质。
其推定氨基酸序列具有锌结合模体HEXXHX 18E ,N 2末端具有17个氨基酸的疏水性信号序列,C 2末端还具有22个氨基酸的糖基磷酯酰肌醇(G PI )添加信号肽。
比对抗性和敏感棉铃虫cDNA 的开放阅读框,抗性品系的开放阅读框中,有57个点突变,共导致了15个氨基酸的改变,其中2个突变(谷氨酰胺137→谷氨酸、缬氨酸173→苏氨酸)位于Cry1A 毒素结合区域,可能与棉铃虫对转Bt 基因棉产生抗性有关。
报道的氨肽酶cDNA 序列已提交G enBank ,AY 346383和AY 279535分别是Bt 抗性和敏感品系的Haapn2。
关键词 棉铃虫 氨肽酶N Bt 受体收稿日期:2004206228 修回日期:20042082093国家自然科学基金项目(30270889);教育部博士点基金项目(2000030703)33通讯作者,电子信箱:jlshen @ 苏云金杆菌(Bacillus thuringiensis )是防治害虫的重要微生物杀虫剂,其产生杀虫毒素的基因也已被广泛用于培育抗虫的转基因植物。
我国华北地区大量栽培的转Bt 基因棉在棉铃虫的防治中已发挥了重要作用。
由于转Bt 基因棉在整个生长期都表达Bt 毒蛋白,持续地给棉铃虫田间种群较高的选择压力,使棉铃虫产生适应性的抗性进化,这种抗性风险已成为转基因棉能否在生产上持续有效应用的最严重威胁[1],因此必须实施配套的抗性治理方案,弄清抗性的分子机理是制订有效抗性治理对策与开展早期抗性监测所不可缺少的基础。
昆虫对Bt 毒素的抗性主要与昆虫中肠毒素结合位点的丧失或与毒素亲和性下降有关[2]。
在鳞翅目昆虫中氨肽酶N 是Bt 毒素蛋白的一类重要受体,用RNA 干涉技术(RNAi )抑制氨肽酶的表达可降低斜纹夜蛾对Bt 毒素的敏感性[3],将敏感昆虫的氨肽酶基因通过增强子捕捉技术在果蝇中肠表达,可使原来对Cry1Ac 毒素不敏感的果蝇变得对毒素非常敏感[4],表明在Bt 毒素杀虫过程中氨肽酶起着非常重要的作用,为了探明棉铃虫对转Bt基因棉(表达Cry1Ac )产生抗性的分子机理,我们对转Bt 基因棉抗性品系和敏感品系棉铃虫幼虫中肠的氨肽酶基因分别进行了克隆鉴定。
1 材料与方法111 供试昆虫棉铃虫敏感品系Y CS 是1991年采自河南偃师市棉田第二代,在室内不接触任何农药,经人工饲料饲养120代供测试。
抗性品系Y CR 用Y CS 室内人工饲料饲养53代后的群体在室内用转Bt 基因棉叶连续筛选50多代,对Bt 原毒素Cry1Ac 和21%MVP Ⅱ可湿性粉剂的抗性达1600倍以上,其抗性表现为单个主效基因的不完全隐性遗传[5]。
试虫采用人工饲料饲养供测试。
饲养条件:温度(27±1)℃,相对湿度60%~80%,光周期14∶10(L ∶D )h 。
112 cDNA 合成、PCR 扩增与测序具体按照苏建亚等[6]介绍的方法进行。
根据G enBank 中已登录的其他鳞翅目昆虫的氨肽酶N序列设计3对引物,分别扩增其5′末端(上游:G CG TGG TG TT AT CC AAACG,下游:G AT ACC ACTG A 2AG TG T CCTG )、中部(上游:TGG TGGG AT AACG T CT 2GG AT C ,下游:CC AATG TTG C AAG CG AATG AC )和3′第24卷第10期中 国 生 物 工 程 杂 志CHI NA BI OTECH NO LOGY2004年10月末端(上游:G AC ACC AG TT ACT ACT CG TGG,下游:C AAC ATTG CG AG T ACTG T CC )。
所有PCR 均在PT C 2200DNA engine (M J Research ,Inc 1)上进行,50μl 反应体系中含:5μl 10×Reaction Buffer ,2m m ol ・L-1MgCl 2,012m m ol ・L -1・each dNTP ,1μm ol ・L -1上Π下游引物,1μl cDNA 模板,215U Ex T aq DNA 聚合酶(T akara 公司)。
反应条件为:95℃预变性3min ,94℃变性1min ,55℃退火1min ,72℃延伸115min ,35个循环。
图1 棉铃虫幼虫中肠氨肽酶(H aapn2)的氨基酸序列及其结构特征Fig.1 Deduced amino acid sequence of aminopeptid ase cDNA H aapn2isolated from Helicoverpa armigera H übner下划双线为信号肽序列,下划单线为Cry1A 毒素结合区域,下划点线为锌结合基序,黑框氨基酸为敏感品系与抗性品系有差异的氨基酸APN2-S =clone from susceptible strain (AY 346383),APN2-R =clone from resistant restrain (AY 279535).The putative signal peptides are double 2underlined ,Cry1A toxin binding region is underlined ,The zinc binding m otif is dotted underlined ;Different am ino acid between susceptible and resistant strain are boxed113 序列分析用BioEdit 软件进行cDNA 和氨基酸序列的分析工作,序列同源性分析在http :ΠΠw w w 1ncbi 1nlm 1g ov Πblast Π网站上进行。
另外用Signal P 程序(http :ΠΠw w w.cbs.dtu.dk Πservices ΠSignalP )和Big 2πProdictor 程序(http :ΠΠmendel.im p.univie.ac.at Πsat Πgpi Πgpiserver.html )分析氨肽酶的N 2末端和C 2末端信号序列。
2 结 果211 棉铃虫幼虫中肠氨肽酶的序列特征通过对敏感棉铃虫幼虫中肠氨肽酶N 的克隆,我们鉴定了氨肽酶Haapn2的cDNA 序列(G enBank 登录号:AY 346383),其cDNA 序列具有3209个核苷酸,包含3096bp 的开放阅读框,编码产生1032个氨基酸的蛋白质。
用Signal P 程序分析Haapn2的蛋白质序列表明,N 2末端具有17个氨基酸的疏水性前导信号序列,该信号肽引导新合成的氨肽酶穿过细胞膜,到胞外后即在丙氨酸172天冬氨酸18之间断裂。
在其C 2末端还有22个氨基酸的疏水性信号序列,该信号序列前是一个小氨基酸(丝氨酸1010),糖基磷酯酰肌醇(G PI )锚即添加在该氨基酸上,氨肽酶通过该G PI 锚固定在质膜的外侧。
该酶还存在所有氨肽酶均具有的锌离子结合模体(HEXXHX 18E ),但未在该蛋白序列上发现糖基化位点(见图1)。
212 抗性品系与敏感品系棉铃虫氨肽酶序列差异通过比较氨肽酶APN2的cDNA 序列,在敏感品系与抗性品系(AY 346383和AY 279535)间共发现有57个碱基替代,在推定的氨基酸序列中则仅有15个氨基酸的变化,即异亮氨酸72→亮氨酸、丙氨酸95→苏氨酸、苏氨酸108→丝氨酸、丝氨酸121→苏氨酸、谷氨酰胺137→谷氨酸、缬氨酸173→苏氨酸、精氨酸220→甘氨酸、精氨酸222→赖氨酸、苏氨酸225→天冬氨酸、天冬氨酸259→天冬酰胺、甘氨酸475→精氨酸、甲硫氨酸574→缬氨酸、亮氨酸638→苯丙氨酸、丝氨酸862→脯氨酸、丝氨酸874→甘氨酸,其中有2个突06中 国 生 物 工 程 杂 志第24卷变(谷氨酰胺137→谷氨酸和缬氨酸173→苏氨酸)位于Cry1A毒素结合区域。
213 序列同源性分析对Haapn2的氨基酸序列利用Blast进行同源搜索,结果发现这个氨肽酶与许多其他昆虫的氨肽酶有很高的相似性。
其相似性为:小菜蛾(AF222699)63%,烟芽夜蛾(AF378666)39%,舞毒蛾(AF317619)40%,斑实夜蛾(AF217249)40%,斜纹夜蛾(Spodoptera litura)(AF320764)39%,烟草天蛾(AF498996)40%,家蚕(AB013400)38%,冈比亚按蚊(X M313484)35%,果蝇(CG31198)32%,埃及伊蚊(Aedes aegypti)(AF378117)32%。
与氨肽酶基因家族中棉铃虫其它氨肽酶的同源性为:APN1 (AY358034)37%、APN3(AY279537)34%、APN4 (AY279534)40%。
3 讨 论Bt毒素杀虫的机制涉及到几个过程,即毒素蛋白的活化、与受体结合、形成寡聚体、插入细胞膜形成膜孔等。
毒素与敏感昆虫幼虫中肠上的氨肽酶N受体高亲和性结合是毒素插入细胞膜形成膜孔的前题[2],为探明毒素蛋白与受体的结合机理,在国外已有多种鳞翅目昆虫的氨肽酶N基因被克隆鉴定,如家蚕(Bombyx mori)[7]、小菜蛾(Plutella xylostella)[8]、烟芽夜蛾(Heliothis virescens)[9]、舞毒蛾(Lymantria dispar)[10]、烟草天蛾(Manduca sexta)[11]。
用生物信息学方法分析Haapn2序列,认为Haapn2具有氨肽酶N的典型特征,是棉铃虫氨肽酶N基因家族中的一员。
其N末端具有引导新合成的多肽穿过细胞膜到胞外的信号肽序列;C末端具有糖基磷酯肌醇(G PI)添加信号位点,氨肽酶被运送到胞外后即在此位置添加G PI锚,并以G PI锚固定在细胞膜上;还具有金属蛋白酶家族所具有的共有序列———HEXXH模体,该模体是锌离子结合位置,但在该氨肽酶序列中没有发现糖基化位点。
鳞翅目昆虫幼虫中肠肠道氨肽酶N目前已知的至少有4个成员,本文报道的是其中一个,棉铃虫的另外2个氨肽酶(apn3和apn4)也同样具有氨肽酶N的典型特征[6],但apn1没有G PI添加信号肽序列(待发表)。