神经肌肉兴奋原理及细胞膜结构
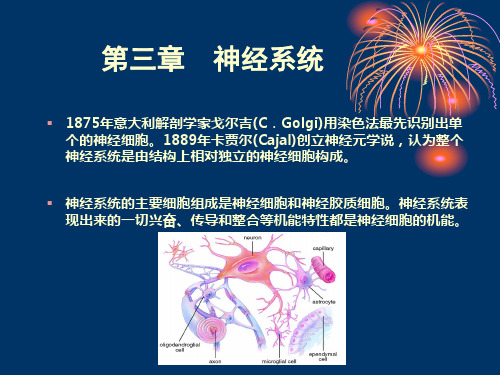
【公共】神经生物学 第三章 解剖生理学-神经系统
2. 骨骼肌的收缩机制
(1)骨骼肌收缩的肌丝滑行学说 (2)兴奋收缩偶联
生物电活动和机械收缩相伴随的事件。
3. 骨骼肌的机械收缩
(1).等张收缩与等长收缩
(2).单收缩与强直收缩 肌肉单收缩呈现等级性,但单条肌纤维收 缩符合“全或无”,收缩无等级性。
完全强直收缩和不完全强直收缩
人的随意活动是由不同程度强直收缩所构成的。
多巴胺循环通路经常和5-羟色胺通路在一些点上出现 交叉和融合,这两种通路可能会同时对某些行为产生影响。 例如,多巴胺与探索、外向、追求愉悦的行为有关,而5羟色胺则与抑制有关。这两个系统在某种意义上互相平衡。 一些药物可以作用于5-羟色胺系统,包括三环类抗抑 郁药和选择性5-羟色胺再摄抑制剂。这些药物被用于治疗 很多心理障碍,尤其是焦虑心境和饮食障碍。
四、骨骼肌的收缩
1. 骨骼肌的功能解剖和超微结构
粗肌丝和细肌丝构成肌原纤维 粗肌丝由肌球蛋白组成;细肌丝含有肌 动蛋白、 原肌球蛋白和肌钙蛋白。
骨骼肌纤维(细胞)的超微结构:
1、肌原纤维: 粗肌丝和细肌丝
2、肌膜:肌细胞膜
横小管(transverse tubule),又称T小管可将 肌膜的兴奋迅速同步地传导至肌纤维内部. 3、肌质网 ★结构:是肌纤维内高度发达的滑面内质网,形成 纵小管(longitudinal tubule),又称L小管; 终池 (terminal cisternae);三联体(triad )
5-羟色胺(5-HT)
5-羟色胺又名血清素,最早是从血清中发现的。脑 内5-HT具有广泛的功能,参与情绪调节、饮食、觉醒-睡 眠周期、痛觉、体温、性行为、梦和下丘脑-垂体的神经 内分泌活动的调节。 5-羟色胺系统的功能之一是缓和调节我们的反应。适 当的5-羟色胺的水平可以使饮食行为、性行为和攻击行为 等处于很好的控制之下。 如果大脑中的5-羟色胺循环通路受到损伤,会发现自 己对脑子里的每个念头和冲动都会付之于行动,使机体表 现得过分活跃:情绪不稳定、好冲动以及对环境过度反应 常常和5-羟色胺的活性极度降低联系在一起,攻击性行为、 自杀、过度饮食和活性降低有联系。
人体解剖生理学08第4章 神经和肌肉

反应
兴奋
刺激
反应
内在 特性
一、刺激与反应
刺激:能为人体所感受而产生反应的环境变 化。
种类:物理性刺激 化学性刺激 生物性刺激 社会心理性刺激
反应:由刺激而引起的机体细胞、组织、器 官或整体的活动状态的改变。 不同组织对刺激的反应速度差异很大。(see and memory) 其本质是:兴奋和抑制(动作电位 的产生)
(二)兴奋和抑制
兴奋和抑制:是反应具有的两种最基本的表现形式。
神经和肌肉等组织,受到有效刺激后在细胞膜上可以产生 一种快速的、可传导的电位波动,这称之为冲动 (impulse)。 生理学上把活组织因受到刺激而产生电冲动的反应称为兴 奋(excitation)。
如果受到刺激后组织的生理活动由原来的显著活动 状态转为相对静止状态、或者活动由强变弱,则称 为抑制(inhibition)。 抑制是组织不再活动? No!是兴奋程度的减弱。
RP: -90mV
极化
复极化
神经纤维
-100mV
超极化
二、动作电位
(一)动作电位的概念
当细胞受到一个有效刺激之后,其膜电位会在静息电位 的基础上发生一次可以沿着细胞膜快速传导的一过性的 电位波动,这种发生在细胞膜上的电波称为动作电位。 特点: 动作电位是细胞受刺激后处于兴奋状态的标志,脉冲式 产生。 动作电位是电位连续快速变化的过程,有 “全或无” 现象 动作电位一经产生便会沿着细胞膜向四周快速传播,呈 现不衰减性传导。
兴奋(Excitatory)
a b c d
兴奋(Excitatory
抑制(Inhibitory)
牛蛙实验报告实验原理

一、实验背景牛蛙作为一种两栖动物,因其独特的生理结构和生活习性,成为生理心理学实验研究的理想动物模型。
牛蛙的坐骨神经腓肠肌标本制备实验,是生理心理学实验中常见的实验之一。
本实验旨在通过观察不同强度和频率的刺激对牛蛙腓肠肌收缩的影响,探讨神经肌肉兴奋传导的原理。
二、实验原理1. 神经肌肉兴奋传导原理神经肌肉兴奋传导是指神经纤维上的动作电位通过神经肌肉接点传递到肌肉细胞膜,使肌肉细胞产生收缩的过程。
在神经肌肉兴奋传导过程中,坐骨神经作为传入神经,将神经冲动传递到腓肠肌细胞膜上的受体,引起肌肉细胞的兴奋和收缩。
2. 刺激与肌肉收缩的关系肌肉收缩是肌肉细胞对神经冲动的反应。
在实验中,通过给予不同强度和频率的刺激,观察腓肠肌的收缩情况,可以分析刺激与肌肉收缩之间的关系。
3. 牛蛙坐骨神经腓肠肌标本制备原理牛蛙坐骨神经腓肠肌标本制备实验,主要利用牛蛙坐骨神经与腓肠肌之间的解剖关系。
坐骨神经作为传入神经,将神经冲动传递到腓肠肌细胞膜上的受体,引起肌肉细胞的兴奋和收缩。
通过制备坐骨神经腓肠肌标本,可以直观地观察刺激与肌肉收缩之间的关系。
4. 不同强度和频率刺激对肌肉收缩的影响(1)刺激强度:刺激强度是指给予神经肌肉接点的电流强度。
在一定范围内,刺激强度与肌肉收缩幅度呈正相关。
当刺激强度低于阈值时,肌肉不产生收缩;当刺激强度超过阈值时,肌肉收缩幅度随着刺激强度的增加而增大。
(2)刺激频率:刺激频率是指单位时间内给予神经肌肉接点的刺激次数。
在一定范围内,刺激频率与肌肉收缩频率呈正相关。
当刺激频率低于一定值时,肌肉不产生收缩;当刺激频率超过一定值时,肌肉收缩频率随着刺激频率的增加而增大。
三、实验方法1. 标本制备(1)处死牛蛙,迅速取出坐骨神经和腓肠肌。
(2)将坐骨神经和腓肠肌放入生理盐水中,以保持标本的新鲜度。
(3)用手术刀将坐骨神经和腓肠肌分离,暴露出腓肠肌细胞膜。
2. 刺激与观察(1)将坐骨神经连接到刺激器上,腓肠肌连接到肌电图记录仪上。
神经系统的细胞结构与功能

神经系统的细胞结构与功能神经系统是人体重要的组成部分,它通过特定的细胞结构和功能来实现信息的传递和处理。
了解神经系统细胞的结构和功能对于理解大脑和神经生物学至关重要。
本文将详细介绍神经系统细胞的结构和功能,帮助读者更好地理解神经系统的运作机制。
一、神经元的结构和功能神经元是神经系统的基本单位,它具有接收、传导和传递神经信号的功能。
神经元主要由细胞体、树突、轴突和突触组成。
细胞体包含细胞核和细胞质,是神经元内部最重要的结构,负责维持细胞的生理功能。
树突广泛分布于细胞体周围,是接收其他神经元传递来的信号的主要部位。
轴突较长,负责传导信号到其他神经元或靶组织。
突触位于轴突末梢,是神经元之间传递信号的联系部分。
二、突触的结构和功能突触是神经元之间传递信号的连接点,主要包括突触前细胞膜、突触间隙和突触后细胞膜。
突触前细胞膜位于信息传递的起始端,负责释放神经递质。
突触间隙是突触前细胞膜和突触后细胞膜之间的间隙,神经递质通过突触间隙传递到下一个神经元。
突触后细胞膜接收神经递质,将信息传递给下一个神经元。
三、神经系统的功能神经系统具有多种功能,包括感知、运动、思维和记忆等。
感知功能是指通过感受器接收外部刺激,并将其转化为神经信号传递到大脑进行处理。
运动功能是指通过神经系统控制肌肉的收缩和松弛,实现身体的运动和动作。
思维功能是指通过神经网络对外界信息进行加工和分析,形成思考和判断能力。
记忆功能是指通过神经系统储存和提取信息,使得人类可以学习和记忆。
四、神经系统疾病与细胞结构功能的关系神经系统的细胞结构和功能异常会导致神经系统疾病的发生。
例如,帕金森病是由于多巴胺神经元的退化导致的,会引起运动障碍和肌肉僵硬。
阿尔茨海默病则是与神经元突触连接异常相关,导致记忆力和认知能力的下降。
对神经系统细胞结构和功能的深入研究可以帮助科学家更好地理解神经系统疾病的发生机制,并有望为疾病的治疗提供新的思路和方法。
总结:神经系统的细胞结构和功能对于神经信号的传导和处理起着至关重要的作用。
神经-肌肉接头传递

神经-肌肉接头传递动物最显著的特点是运动功能,各种运动都是由肌肉收缩完成的。
骨骼肌属于随意肌,在中枢神经控制下接受躯体运动神经的支配。
只有当神经纤维上有传出神经冲动,并经骨骼肌的神经-肌接头把兴奋传递给骨骼肌,才能引起骨骼肌的兴奋和收缩。
神经-肌肉接头(neuromuscular junction)概念和结构概念:神经-肌肉接头是由运动神经纤维末稍和它接触的骨骼肌细胞膜所构成,是一种特化的突触(synapse)。
神经末梢在接近骨骼肌细胞处失去髓鞘,每一个裸露的轴突末梢进入肌肉后又广泛分支形成大量末端呈膨大的突触前终扣(presynaptic terminal button),每个终扣各嵌入一条与它相对应的、有肌膜向内下陷形成的凹陷(或称终板)中,共同形成一个神经-肌接头。
组成部分:①接头前膜(prejunctional membrane):嵌入肌细胞膜凹陷中的突触前终扣的膜;②接头后膜(postjunctional membrane):与接头前膜相对应的肌膜,也称为终板膜(endplate membrane);③接头间隙(junctional cleft):接头前膜与接头后膜之间的一个达50 nm的间隙,充满细胞外液。
突触前终扣的胞质内存在大量突触囊泡(synaptic vesicle),直径约50~60 nm,每个囊泡含有6000到10000个乙酰胆碱(acetylcholine,Ach)分子。
ACh 分子能够与终板膜上的烟碱型乙酰胆碱受体(nicotinic acetylcholine receptor,nAChR)特异性结合。
nAChR集中分布于终板膜皱褶的顶部,属于化学门控阳离子通道。
N-M接头处兴奋传递的主要步骤N-M接头之间的信号传递是通过神经递质乙酰胆碱的介导完成的。
概括为“神经-乙酰胆碱-肌肉”或者“电信号-化学信号-电信号定向转换”过程。
神经冲动沿神经纤维传到轴突末梢时,接头前膜首先发生去极化;膜的去极化引起该处膜上存在的电压门控钙通道开放,钙离子内流,接着接头前膜胞质内钙离子浓度快速增高;钙浓度的增高促使突触小泡向接头前膜内侧移动、进而小泡膜与接头前膜融合、融合处出现小孔,经胞出过程将小泡中的ACh分子全部释放至接头间隙;ACh分子经扩散与终板膜上的nAChR结合,并激活这种受体而使其分子结构中的通道样结构开放,于是出现钠离子内流为主的跨膜离子移动,使终板膜发生去极化,产生终板电位(endplate potential, EPP);EPP以电紧张形式扩布至临近的肌细胞膜,引起肌细胞爆发动作电位,最终完成电信号由接头前膜到肌细胞膜的一次兴奋传递。
神经肌肉一般生理学

K+,分解ATP。
• Na+泵有2个亚基,2个亚基, 亚基有与离子、哇巴因 (ouabain)结合位点,有ATP 酶活性;但解离亚基,Na+泵失 活。
• 3Na+与泵结合,ATP酶激活, ATP分解,泵磷酸化,泵构象变 化,3Na+移出胞外, 2K+与泵结 合,去磷酸化。
• 取决于通道是否开放及开放的程度及数量 • 取决于膜两侧的浓度差或电位差
通道是否开放: ①电压/化学/机械变化; ②时间 功能:不是转运代谢物,而是离子流动引起电位变化,
将外来信号转变为细胞自身信号——电变化
细胞膜的转运功能
主动转运(active transport)
• 原发性主动转运(primary active transport)
细胞的跨膜信号转导
细胞跨膜转导的类型 跨膜信号转导的途径与机制 跨膜信号转导系统相互影响 细胞通讯 细胞信号转导的基本特征
细胞跨膜转导的类型
虽然跨膜信号转导涉及多种刺激信号,在多种细 胞引发多种功能变化,但转导途径是有限的。
转导途径一般有两种: • 根据感受和传导过程分 • 根据受体存在的部位分
不同内细肌胞醇有磷可酸能信G号相系同统,;有G可g:能味不觉同细。胞G。t:视杆细胞;Go:脑 基本结构:100KD,、、三个亚基,主要是,既是GTP
结合点,又是GTP酶。 过去认为起锚钉作用,仅对亚基功能起调节作用,现在 发现也可激活胞内靶分子。除调节AC、PLC、离子通道 外,还可参与激活TKR转导系统。也能与效应器酶结合, 对亚基起协调拮抗作用。有些细胞毒素可修饰亚基,改 变生理特性。
• 鸟苷环化酶受体(GC-R) 第二信使有:cAMP/ cGMP/IP3 / DG / Ca2+ PK有:PKA / PKG / CaMK / PKC / Ca2+-PK
生理学动作电位肌肉收缩原理

第二信使学说
第一信使+R
G蛋白-GDP G蛋白-GTP
蛋白激酶 及其他
第二信使
第二信使前体 细胞功能改变
效应器酶
1 第一信使:激素、递质等 2 效应器酶:腺苷酸环化酶、磷酯酶C等 3 第二信使:cAMP、IP3、DG
①受体-G蛋白-AC途径: cAMP ② 受体-G蛋白-PLC途径: IP3、DG
意义:是细胞处于兴奋状态的标志。 阈电位:能触发动作电位的膜电位临界值 。阈电位大约比
正常静息电位的绝对值小10~20mV 。
动作电位的产生条件:静息电位去极化达到阈电位水平。
神经纤维AP
心肌细胞AP
动作电位的特点
⑴动作电位呈“全或无”现象:动作电位一旦产生 就达到它的最大值,其变化幅度不会因刺激的加 强而增大;
由受体完成的跨膜信号传递
受体:细胞中能与某些化学物质特异性结合,引发细胞 特异生物学效应的特殊结构。
受体本质:蛋白质或酶 受体特征: ①特异性
②饱和性 ③可逆性
第二信使学说:激素+受体→第二信使→酶激活
激素的作用机制:
1、通过膜受体传递调控信息: 第二信使学说
2、通过胞内受体传递调控信息: 基因表达学说
主动转运
被动转运
需由细胞提供能量 逆电-化学势差 使膜两侧浓度差更大
不需外部能量 顺电-化学势差 使膜两侧浓度差更小
(四)入 胞 和 出 胞
第二节 细胞的跨膜信号转导功能
跨膜信号转导方式分为三类: ① 离子通道介导的信号转导。 ② G蛋白耦联受体介导的信号转导; ③ 酶耦联受体介导的信号转导; 每类都通过各自不同的细胞信号分子完成信号转导。
⑶ 能影响肌肉收缩效果的肌肉内部功能状态。
第一章肌肉的兴奋与收缩

返
细肌丝
回
肌肉的收缩过程
兴奋——收缩耦联 横桥运动引起的肌丝滑行
兴奋——收缩耦联
动作电位沿横管系统 传向肌细胞深部
三联管结构传递 信息
纵管系统对Ca2+的 释放和再聚积
返回
返
横桥运动引起的肌丝滑行
回
1. 当肌浆中离子浓度升高,Ca2+与肌钙蛋白结合,肌动蛋白失去 钩子作用,原肌球蛋白的双螺旋体从肌动蛋白双螺旋结构的沟沿滑 到沟底,暴露出肌动蛋白上与横桥的结合点,含有ATP的横桥与位 点结合,形成肌动蛋白、肌球蛋白——ATP复合体。与此同时,横 桥中的肌球蛋白ATP酶受肌动蛋白激活,使横桥中的ATP迅速水解 成ADP+Pi;放出能量,引起横桥头部向粗肌丝中心方向摆动,牵 拉细丝向肌节中央滑行。
当冲动从神经纤维传至轴突末梢时,轴突末梢出现除 极化,改变神经膜的通透性,使细胞外液中Ca2+进入末梢内, 引起轴浆中200~300个突触小泡破裂,释放出乙酰胆碱,进入 接头间隙。当乙酰胆碱经接头间隙到达终板膜表面时,立即与 膜上的特殊受体(R)相结合,形成R—ACH复合体,引起膜
对Na+、K+通透性改变,而导致除极化,进而触发一个 可传导的动作电位沿肌膜传播至整个肌纤维,引起整 条肌纤维收缩。
局部兴奋的特点
不呈现“全或无”定律
电紧张性扩布 没有不应期 有总和现象
返回
6 兴奋在神经肌肉接 头的传递
神经和肌肉之间无直接的原生质联系, 但神经冲动可传递给肌肉引起肌肉收缩。 这种机能联系是通过神经肌肉接点而进行 的。
神经肌肉接头的结构
突触前膜
突触后膜 突触间隙
兴奋在神经肌肉接头传递的
机制
动作电位的产生机制
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
坐骨神经-腓肠肌模型 •将分离的坐骨神经-腓肠肌模型浸 入任氏液( —种比较接近两栖动物 内环境的液体)备用;
•用沾有任氏液的锌铜弓触及一下坐 骨神经或用鑷子夹持坐骨神经中枢 端,腓肠肌发生迅速而明显的收缩;
•然后再用食盐(或1% H2SO4滤纸) 刺激坐骨神经中枢端(或肌肉), 腓肠肌发生迅速而明显的收缩。
– 其中最重要的成分是磷酸甘油脂(phosphoglycerides)
磷脂分类
• 主要有四种:
– 磷脂酰乙 醇胺、
– 磷脂酰丝 氨酸、
– 磷脂酰胆 碱(卵磷 脂)、
– 鞘磷脂
胆固醇
糖脂位于细胞膜的外层
Gal: 半乳糖
Glc: 葡萄糖
GalNAc: 乙酰半乳 糖胺
神经节苷脂是一 类重要的糖脂
膜脂的性质
• 1895年Overton研究各种未受精的卵细胞,发现脂 溶性物质容易通过细胞膜
– 说明细胞膜是由脂类构成的,离子很难通过 – 细胞外表面是亲水的 – 说明……
• 脂质、胆固醇 • 植物研究中的启发
– 你猜测细胞膜应该是什么样的构造?
细胞膜的发现
细胞膜的发现
人工制作的磷脂双分子层小 泡——脂质体
细胞膜的发现
• 1925年Gorter 和Grendel用丙酮提取红细胞膜脂, 并将其在空气/水界面上展开,萃取得到了物质可 以铺展成两倍红细胞膜的面积
细胞膜的发现
• 你们怎么评价Gorter 和Grendel的实验? • 有什么问题?
– 丙酮萃取出的膜脂仅占总膜脂的70% – 单层油脂分子的间距是可变的 – 他们用血图片的办法测量红细胞表面积 – Langmuir Trough的使用方式 – 后来的更有趣的实验(Zwall, et al., 1975),脂膜对糖脂酶
的耐受,给出更确切的估计
细胞膜的发现 • Danielli and Davson 模型(1935)
Danielli and Davson (1935) proposed a membrane model including proteins. They used their model to interpret the observation of different membrane permeabilities of ions and hydrocarbons. In particular, they assumed that the membrane has both a lipophilic and a hydrophilic character. Watercontaining regions in the membrane give rise to ion transport depending on water content of the membrane and its charge; lipophilic parts are responsible for the transport of water-insoluble molecules.
membranes – considered the role of hydrophobic amino acids in α-
helices
细胞膜的发现
• Domain Formation and Protein Clusters
During lipid melting transitions the thickness of lipid membranes and the lateral lipid distribution changes. This is of extreme interest for regulation and signal transduction purposes in such membranes
细胞膜的发现
• The Fluid Mosaic Model of Singer and Nicolson (1972)
– the structures of a number of soluble proteins were solved by X-ray crystallography
– many membrane proteins have a high α-helical content – that labeled proteins form isolated spots in some
• 脂膜是一种二维液体 • 膜脂在脂膜中很容易侧向运动(流动) • 但是除胆固醇之外,所有其它膜脂都无法
快速地从内侧翻转到外侧,反之亦然 • 脂膜的流动性受到膜脂成分的影响 • 双层膜的厚度受到膜脂特性的影响
(A) 卵磷脂与鞘磷脂1:1混合制 备的脂质体中,两种膜脂均匀 分布;(B) 卵磷脂、鞘磷脂、胆 固醇1:1:1混合制备的脂质体中, 鞘磷脂与胆固醇形成脂筏
•18世纪末期的发现——蛙腿论战 •关键人物伽伐尼\伏打\诺伯里
触发神经——肌肉反射的物质是什么?
背景:人们已经发现了电的 基本原理
1. 电信号、离子? 2. 传播方式? 3. 是离子在细胞中运动吗? 4. 细胞是一个整体,离子怎
样进入?
关于细胞膜的背景
• 离子在细胞中流动?
– 细胞膜的发现似乎不支持这种看法
细胞膜的发现 • Robertson 电镜照片(1958)
Robertson (1959) collected electron microscopy images of many cells and organelles. His picture of a biological membrane resembles that of Danielli and Davson (1935). However, it is now based on microscopic evidence.
细胞膜的发现
• 现代细胞膜模型
细胞膜的基本结构特点
• 细胞பைடு நூலகம்由膜脂和蛋白质构成 • 细胞有500-1000种不同的膜脂,是细胞膜基
本结构 • 这些膜脂可以分为三大类:
– 磷脂、胆固醇、糖脂 – 它们的结构和功能有各自的特点
• 蛋白质完成膜的主要功能,包括
– 受体、酶、转运蛋白
磷脂
• 细胞膜脂的主要成分