糖原代谢及其调控
糖有氧氧化的调控机制

糖有氧氧化的调控机制糖是人类最主要的能量来源之一,它们在人体内通过氧化反应进行代谢产生能量。
糖的氧化是一个复杂的过程,受到多种调控机制的控制。
本文将详细介绍糖的有氧氧化的调控机制。
糖的有氧氧化是指糖分子在细胞内被氧化成二氧化碳和水释放能量的过程。
这一过程主要发生在线粒体内,通过好氧呼吸途径来完成。
糖的氧化分为两个阶段:糖代谢产生的丙酮酸、糖酸等化合物经过解羧酶作用形成乙酰辅酶A,然后在三羧酸循环中被进一步氧化,生成二氧化碳和高能形式的辅酶。
糖的有氧氧化受到多种调控机制的调节,这些机制可以分为内源性和外源性两个层面。
内源性调控机制主要包括酶系统和能量代谢途径的调控。
首先是酶系统的调控。
糖的有氧氧化过程中涉及到多种酶的参与,这些酶的活性受到多种因素的调控。
例如,三羧酸循环中的异戊二酸脱氢酶活性受到碳氮比的调控,当碳氮比较高时,酶活性增强,进一步促进糖的氧化。
另外,磷酸化酶和解磷酶对糖酸的磷酸化和解磷酸化也是有调控作用的。
其次是能量代谢途径的调控。
糖的有氧氧化是能量代谢途径的一部分,能量代谢途径的调控也会影响糖的有氧氧化。
例如,细胞内ATP/ADP比值升高会抑制糖的有氧氧化,因为高ATP/ADP比值代表细胞内能量充足,不需要过多的糖分解来产生能量。
外源性调控机制主要包括内分泌系统和神经系统的调控。
内分泌系统通过激素的分泌来调控糖的有氧氧化。
胰岛素是最为重要的糖调节激素之一,它主要通过促进葡萄糖转运蛋白的合成并促进葡萄糖的摄取,从而提高糖的有氧氧化。
胰高血糖素则有相反的作用,它促进糖原的分解和糖的释放,从而降低糖的有氧氧化。
神经系统通过神经递质的作用来调控糖的有氧氧化。
交感神经系统的活性增强会抑制糖的有氧氧化,而副交感神经系统的活性增强则会促进糖的有氧氧化。
除了内源性和外源性调控机制,糖的有氧氧化还受到基因表达调控的影响。
内源性和外源性因素可以通过调控与糖有氧氧化相关的基因的表达来影响糖的有氧氧化过程。
糖代谢的生物化学调节

糖代谢的生物化学调节糖代谢是生物体内一个重要的代谢过程,通过一系列的生物化学反应,将摄入的碳水化合物转化为能量和存储形式。
这一过程涉及多个关键酶的调节,以保持机体内部代谢平衡。
本文将探讨糖代谢的生物化学调节机制。
1. 糖代谢的基本过程糖代谢的基本过程主要包括糖的吸收、储存、释放和利用。
当我们进食含糖食物时,消化系统中的酶将复杂的糖类分解为单糖,如葡萄糖。
这些单糖通过细胞膜转运蛋白进入细胞内,并在细胞质中进行代谢。
2. 葡萄糖调节机制葡萄糖是糖代谢的主要物质,其浓度在血液中需要维持在一定的范围内。
当血糖浓度过高时,胰岛素释放,促进葡萄糖的摄入和利用。
胰岛素通过激活葡萄糖转运蛋白和糖原合成酶,促使葡萄糖转化为糖原储存起来。
当血糖浓度过低时,胰岛素的分泌减少,肝细胞将糖原分解为葡萄糖释放到血液中,以维持血糖水平。
3. 糖原和糖酵解的调节糖原是一种储存在肝脏和肌肉中的多糖,能够释放葡萄糖以满足机体能量需求。
糖原的合成受到胰岛素的促进,而其分解则受到胰高血糖素和肾上腺素的调节。
当机体需要能量时,肾上腺素的分泌增加,激活糖原磷酸化酶,使得糖原分解为葡萄糖。
4. 糖酵解调节糖酵解是将葡萄糖分解为乳酸或丙酮酸的过程,产生少量的ATP。
当氧气供应不足时,糖酵解是细胞的主要能源来源。
糖酵解的过程中,多个关键酶受到调节,如磷酸果糖激酶、葡萄糖激酶和磷酸三磷酸异构酶等。
这些酶的活性可以通过磷酸化、糖酮-糖磷酸酯循环以及底物浓度等因素进行调节。
5. 糖异生的调节糖异生是指在机体无法通过摄入糖类满足能量需求时,通过非糖类物质合成葡萄糖。
糖异生主要发生在肝细胞中,其中多糖、脂肪和氨基酸是糖异生的补给物。
多个酶参与糖异生的调节,其中磷酸烯醇式还原酶和磷酸果糖-6-磷酸酶是关键酶,其活性受到内分泌激素和底物浓度的调控。
总结:糖代谢的生物化学调节涉及多个酶的活性调控,其中胰岛素和肾上腺素是重要的调节激素。
胰岛素在血糖浓度高时促进糖的储存和利用,而肾上腺素则在能量需求增加时促进糖原分解和糖酵解。
糖代谢与调控机制

糖代谢与调控机制糖代谢是维持生物体能量平衡的重要过程。
通过摄入食物,人体获得葡萄糖等糖类物质,这些糖类物质在机体内被分解、合成和储存,以提供能量和维持各种生物功能。
糖类物质的代谢过程受到多个调控机制的影响,以确保能量平衡和正常生理功能的维持。
糖的消化和吸收食物中的淀粉和蔗糖等多糖在消化系统中被酶水解为葡萄糖。
这些葡萄糖分子进入肠道绒毛上皮细胞,通过转运蛋白进入细胞内,并进一步通过转运蛋白进入血液循环。
糖的分解和合成在细胞内,葡萄糖经过糖酵解途径被分解为乳酸或丙酮酸。
这些代谢产物进一步参与能量产生的过程。
此外,葡萄糖也可以通过糖异生途径转化为葡萄糖酮体,以供应特定组织的能量需求。
糖的储存多余的葡萄糖可以在肝脏和肌肉中以糖原的形式储存起来。
当机体需要能量时,糖原会被分解为葡萄糖,并通过糖酵解途径供给能量。
糖代谢的调控机制糖代谢的调控主要由激素、酶活性和细胞信号传导等机制完成。
1. 激素调控:胰岛素和胰高血糖素是体内最重要的糖代谢调控激素。
胰岛素促进葡萄糖的摄取、利用和储存,而胰高血糖素则有利于血糖的升高和糖原的分解。
2. 酶活性调控:糖代谢酶的调节也是糖代谢调控的重要机制。
例如,磷酸果糖激酶和磷酸果糖醛酸酯酶是糖酵解途径中的速率限制酶,它们的活性受到多种信号的调节。
3. 细胞信号传导调控:糖代谢还受到细胞内信号传导通路的调控,如AMP激活蛋白激酶、PI3K/Akt通路等。
这些糖代谢和调控机制相互作用,共同维持机体内糖类物质的平衡和能量供给的适应性。
深入了解糖代谢与调控机制有助于我们更好地理解糖类物质对人体健康的影响,并为疾病的预防和治疗提供理论基础。
糖原代谢和糖异生

另有10%葡萄糖残基要经己糖 激酶催化生成葡萄糖-6-磷酸 进行糖酵解的。
个
15.3 糖原合成
糖原的生物合成不是糖原降解的逆过程,而是通过 另外一条途径。
糖原合成需要的能量是由尿嘧啶核苷三磷酸(UTP) 提供的。
糖原合成的底物是UDP-葡糖。
G o' = -16.3 kJ/mol
旁路III:葡糖-6-磷酸水解生成葡萄糖
葡糖-6-磷酸在葡糖-6-磷酸酶作用下水解为 葡萄糖 和无机磷酸。
G o'= -13.8 kJ/mol
糖
葡糖-6-磷酸酶
异
生
途
径
总
览
图
糖 异 生 与 糖 酵 解 过 程 能 量 变 化
糖异生是个需能过程,由2分子丙酮酸合成1分子葡萄 糖需要4分子ATP和2分子GTP,同时还需要2分子NADH。 总反应方程式为:
3. 糖异生的调控
磷酸果糖激酶I(PFK-I)和果糖-1,6-二磷酸酶的调节
果糖-2,6-二磷酸可以激活PFK-1,加快糖酵解;而抑制 果糖-1,6-二磷酸酶(FBPase-1),进而抑制糖异生。
当ATP和柠檬酸水平高时,PFK-I受抑制,降低糖酵解速 率;柠檬酸增加果糖-1,6-二磷酸酶活性,从而增加糖异 生速率。当AMP水平高时,PFK-I激活,加快糖酵解,果 糖-1,6-二磷酸酶受抑制,糖异生关闭。
但糖酵解途径中由丙酮酸激酶、磷酸果糖激酶I和己 糖激酶催化的三个高放能反应是不可逆的。
1. 糖异生反应
旁路I:丙酮酸转化为磷酸烯醇式丙酮酸
(1)丙酮酸羧化生成草酰乙酸 在丙酮酸羧化酶(生物素作为辅基)催化下,丙酮酸
糖原代谢及其调控精品PPT课件
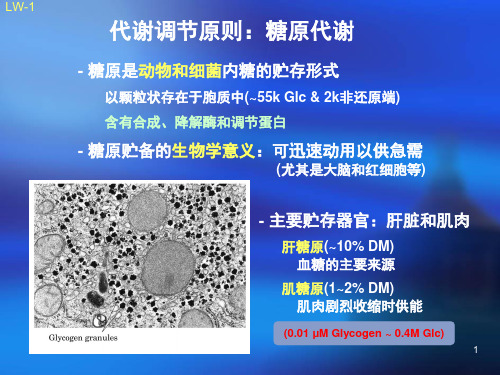
内质网腔
Glc质膜载体 毛 细 血 管
6
H13.4
§2. 糖原合成
- High [Pi] in cell favors glycogen breakdown & prevents from glycogen synthesis in vivo.
- Needs another way to activate Glc for transferring to glycogen chain.
糖原磷酸化酶 断裂(14)
游离异头C (还原端)
- 该过程可重复进 行至离某个分支 点相隔 4 Glc
- Ionized G1P can’t diffuse out of cell - Glc is phosphorylated: no ATP needs to be
consumed to permit entry into glycolysis
directly adding to a chain like this, or needing a primer with at least 8/6 Glc residues
⊿G’o = -13.4 kJ/mol
9
15-9
• Branch synthesis in glycogen
= 糖原分支酶从一段至少有11 Glc残基的分 支上转移6~7个残基给该分支或邻近分支 还原端某个残基的C6上以形成新的分支
activity 1 = 糖基转移酶 脱
支 酶 activity 2 = (16)糖苷酶
4
15-5
• 磷酸葡糖变位酶作用机制
(cf. Fig. 13-7)
- 该酶需以活性位点的Ser 残基已被磷酸化的形式 参与反应
酵母菌对糖代谢的调控机制

酵母菌对糖代谢的调控机制酵母菌广泛应用于生物学研究和工业生产中,其糖代谢调控机制是研究的热点之一。
糖代谢是细胞生命活动的重要组成部分,酵母菌通过调节糖代谢可以适应不同的生存环境。
本文将从糖的运输、糖的调控以及ATP能量供应等方面,探讨酵母菌对糖代谢的调控机制。
一、糖的运输酵母菌利用多种载体蛋白将外源糖从环境中吸收入细胞内部进行代谢。
其中,主要的载体蛋白有Hxt、Gal2、Stl1等。
这些载体蛋白担负着将外源糖通过细胞膜扩散到胞质中的重要任务。
Hxt是葡萄糖、果糖、甘露糖等单糖的共同载体蛋白,其表达水平和糖浓度相关。
当糖的浓度较低时,Hxt的表达水平较高;当糖浓度较高时,Hxt的表达水平下降。
Gal2是氧化型半乳糖和D-半乳糖的载体蛋白,其表达受到Gal3和Gal80的共同调节。
Stl1是L-松针糖和D-木糖的载体蛋白,其表达和糖浓度和ATP水平相关。
通过调节这些载体蛋白的表达水平,酵母菌可以有效控制外源糖物质的吸收入细胞的速度以及代谢产物的积累。
二、糖的调控酵母菌中的糖分解途径主要有两条通路,即酵母菌糖(YP)途径和磷酸戊糖(EMP)途径,它们分别通过碳源和酶的来源不同,对细胞代谢产生不同的影响。
糖的代谢调控主要依赖于以下几个方面:1. 糖信号通路酵母菌中的糖信号通路主要通过六种糖酵解产物参与,它们分别是葡萄糖-6-磷酸(G6P)、二磷酸甘露醇(F-2,P)、香豆酸(U)、3-磷酸甘油(PGA)、丙酮酸(PYR)和乳酸(LAC)。
其中,G6P和F-2,P被认为是主要的糖信号分子,通过信号转导途径逐渐转化为细胞内部的信号代表物,参与调控与糖代谢相关的基因表达和酶活性。
2. Grr1蛋白相关机制Grr1蛋白是酵母细胞中的一种RWD(RING finger and WD40 domain-containing protein)型蛋白,与Ubiquitin连接酶一起参与酵母中糖代谢调控。
Grr1蛋白可以与一系列与糖代谢相关的蛋白结合,包括Hxt1/3/4、Glc7、Hxk2等,从而参与调控这些蛋白的降解和稳定性。
植物新陈代谢途径及其调控

植物新陈代谢途径及其调控新陈代谢是指生物体内的化学反应过程。
植物的新陈代谢包括许多不同的化学反应,用于合成、分解和转化生物分子。
这些反应是保证植物正常生长和发育的必要条件,还可用于植物的适应性反应,以适应不同的环境条件。
本文将介绍植物的新陈代谢途径及其调控。
1. 光合作用和光呼吸光合作用是植物维持生命所必需的重要途径。
在光合作用中,光能被捕获,用于产生高能的化学键,从而合成养分,如葡萄糖和淀粉。
光合作用分为光反应和暗反应两个部分,其中光反应发生在叶绿素中,利用光能产生ATP和NADPH;暗反应在叶绿体基质中进行,利用ATP和NADPH,将CO2转化为葡萄糖和其他有机物。
光呼吸是光合作用的一种反应,仅在缺氧或光能量不足时发生。
它涉及到叶绿体电子传递链的一部分,产生ATP。
尽管光呼吸影响了光合作用的效率,但它也有助于植物维持能量供应。
2. 糖代谢糖代谢是植物的另一种重要途径,用于合成、分解和转化糖类化合物。
葡萄糖是植物体内最常见的糖,但植物也可以合成其他糖类,如果糖、蔗糖和木糖。
糖类产生与分解的速度会受到多种因素的影响,如温度、光照、水分和化学信号。
在糖代谢过程中,植物通过糖原(淀粉)形式储存葡萄糖,当需要时再释放出来,用于供能和碳源。
糖原代谢有大部分在叶绿体中进行,其中包括淀粉的合成和降解。
淀粉的合成可以通过糖原合成酶的作用进行,而淀粉的降解则可以通过树突酶进行。
3. 氨基酸代谢氨基酸是蛋白质的组成部分,也是一些存储和运输分子的基础。
氨基酸代谢过程包括氨基酸合成、分解和转化。
对于植物来说,关键的氨基酸包括谷氨酸、丝氨酸、松香酸和精氨酸。
氨基酸的合成是由多种酶参与的逐步过程。
其中一个重要的反应是谷氨酸合成,它涉及到谷氨酰磷、谷氨酸合成酶和一氧化氮合酶等酶。
当植物遭受到环境压力时,例如高盐、干旱和营养限制,它们的氨基酸代谢过程会发生变化,以提供必要的调节和适应性反应。
4. 脂类代谢脂类代谢过程是植物维持生命所必需的反应之一,是合成和降解脂肪酸、甘油三酯和磷脂分子的过程。
生物化学学习题碳水化合物代谢途径

生物化学学习题碳水化合物代谢途径生物化学学习题:碳水化合物代谢途径碳水化合物代谢是生物体内一系列复杂的生化过程,包括糖原合成、糖原分解、糖酵解、柠檬酸循环和呼吸链等途径。
这些途径相互关联,构成了生物体内碳水化合物代谢的重要组成部分。
本文将详细介绍这些代谢途径及其相关的反应和调控。
1. 糖原合成糖原合成是将多余的葡萄糖合成为糖原储存在肝脏和肌肉中,以供生物体在需要能量时进行分解。
糖原合成主要发生在肝脏和肌肉细胞内,通过磷酸化作用将葡萄糖-6-磷酸转化为糖原。
2. 糖原分解糖原分解是将储存在肝脏和肌肉中的糖原分解为葡萄糖,以供生物体在能量需求增加时进行利用。
糖原分解主要发生在肝脏和肌肉细胞内,通过糖原磷酸化酶的作用将糖原逐步分解为葡萄糖-1-磷酸,然后通过磷酸葡萄糖异构酶和蔗糖酶的作用将其转化为葡萄糖。
3. 糖酵解糖酵解是将葡萄糖分解为乳酸或乙酸,产生少量ATP的过程。
糖酵解主要发生在细胞质中,包括糖酵解途径中的十个反应。
在细胞质中,葡萄糖经过一系列酶催化反应,生成两个丙酮酸分子,然后通过乳酸脱氢酶的作用将丙酮酸转化为乳酸。
4. 柠檬酸循环柠檬酸循环是将乙酸等中间产物氧化为二氧化碳和水,并生成较多的ATP的过程。
柠檬酸循环主要发生在线粒体的基质中,通过一系列酶催化反应将乙酸转化为柠檬酸,再逐步氧化为苹果酸、柠檬酸、草酰乙酸等,最终生成辅酶NADH和FADH2,为呼吸链提供电子。
5. 呼吸链呼吸链是将柠檬酸循环和糖酵解过程中生成的辅酶NADH和FADH2的电子流转化为ATP的过程。
呼吸链主要发生在线粒体内膜上的电子传递链上,通过电子传递过程中的氧化还原反应,将电子从高能态的NADH和FADH2转移至终电子受体氧分子,产生水分子,并伴随着质子泵的作用,形成质子梯度,通过ATP合酶酶的作用合成ATP。
这些碳水化合物代谢途径共同参与了生物体内碳水化合物的合成、分解和能量转化等重要生化过程。
不同组织和不同生理状态下,这些代谢途径受到多种调控机制的精细调节,以满足生物体的能量需求和代谢平衡。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
- 先由酶将其磷酰基转移 给G1P而生成G-1,6-BP
- 再由G-1,6-BP将其C1位 磷酰基转移给酶并释出 G6P
自学
(cf. phosphoglycerate mutase with His in Glycolosis ⑧)
15-6
(无机)焦磷酸酶
⊿G’o = -20~27 kJ/mol
15-8
• Glycogen synthesis = glycogen chain elongated
by glycogen synthase
糖原合酶不能从头开始而将两个游 离的UDP-Glc直接连接起来
transferring the Glc residue from UDP-Glc to the nonreducing end of a glycogen branch to make a new (1→4) linkage
directly adding to a chain like this, or needing a primer with at least 8/6 Glc residues
⊿G’o = -13.4 kJ/mol
15-9
• Branch synthesis in glycogen
= 糖原分支酶从一段至少有11 Glc残基的分 支上转移6~7个残基给该分支或邻近分支 还原端某个残基的C6上以形成新的分支
the reaction forward and
irreversibly
核苷二磷酸糖 ⊿G’o ≈ 0 kJ/mol 焦磷酸化酶
- 核苷二磷酸糖在寡糖和多糖 的生物合成中作为糖基供体
- UDP-Glc for glycogen synthesis in animals
- ADP-Glc for starch synthesis in plants and glycogen synthesis in bacteria
= 引发蛋白+葡糖基转移酶
葡糖基转移活性
转移酶与糖 原合酶结合
糖原合酶活性
合酶与分 支酶活性
葡糖基延长活性
Glycogen core
15-11
自学
• 糖原生成(起始)蛋白反应机制
- 在葡糖基转移酶活性作用 下,Tyr194-OH亲核攻击 UDP-Glc的C1而生成糖基 化的Tyr (~非还原末端)
- 非还原末端Glc的C4-OH对 另一UDP-Glc亲核攻击以 形成(1→4)糖苷键
- 达到8个残基后由糖原合酶 继续延长及分支
15-10
• Muscle glycogenin (dimer)
UDP-Glc, bound to Mn2+ through its phosphates
自学
用作e–对受体以稳 定离去基团UDP
15-4
• 糖原脱分支
- Product of glycogen degradation = G1P (85%) & free Glc (15%)
G1P
- Debranching enzyme
phospho-
= bifunctional enzyme glucomutase
(as PFK-2)
磷酸葡糖
- Needs another way to activate Glc for transferring to glycogen chain.
much better leaving group
糖原 合酶
磷酸葡糖变位酶 UDP-Glc 焦磷酸化酶
UDP-Glc
Luis Leloir 1906-1987 1970 NP in Chem.
断裂(1→4)键
糖原分支的生物学意义 - 增加糖原的可溶性 - 增加非还原端数量
糖原分支酶
形成(1→6)键
20-16(3rd)
ADP-Glc 焦磷酸化酶
• Starch synthesis
自学
- 除了活化底物是ADP-Glc之 外,合成机制与糖原的类似
淀粉合酶
20-14
自学
• 由糖原生成(起始)蛋白开始的糖原颗粒形成
G6P 变位酶
- G6P去路 肝、肾细胞中水解成Glc 脑、肌细胞中直接进入酵解
(without G6Pase) - 糖原颗粒不会被完全分解,
一般是分支减少/分子变小
activity 1 = 糖基转移酶 脱
支 酶 activity 2 = (16)糖苷酶
15-5
• 磷酸葡糖变位酶作用机制
(cf. Fig. 13-7)
G15.15
Structure of Glycogen Phosphorylase
monomer (842 AA)
别构剂 结合点
与另一亚基的 别构部位接触
PLP
自学 dimer
糖原颗粒 结合位点
催化 部位
磷酸化位点: 高活性a型 磷酸化 低活性b型 去磷酸化
辅基磷酸吡哆醛(PLP) 共价结合于Lys680,其 磷酰基以广义酸-碱催化 方式促进Pi攻击 (1→4) 糖苷键 (cf. Fig. 13-5)
糖原代谢中Glc激活方式不同:
- 降解时磷酸解成G1P - 合成时核苷酰化成UDP-Glc
15-7
自学
• 核苷二磷酸糖/糖核苷酸的形成
= O– on the sugar phosphate
attacking nucleophilicly P of NTP and displacing PPi, which hydrolysis pulling
本单体Asp162先 亲核攻击形成过 渡态中间物
Asp162 Tyr194
另一单体Tyr194再 亲核攻击完成反应
LW-2
小结:糖原代谢
• 糖原以颗粒形式储存于肌肉和肝脏,颗粒中还含有 糖原代谢及调节的各种酶
• 糖原磷酸化酶催化糖原链非还原端残基磷酸解断裂 (1→4)键而生成G1P,去分支酶将分支转移到主链 并以游离Glc形式释出(1→6)分支点残基
自学
• 肝糖元降解可以补充血糖
G6P酶仅存在于肝脏和肾脏,为内质网 膜上的整合蛋白(可能有九个跨膜螺旋区 段),活性点位于腔内侧(why?)
T1/G6P酶的任一遗传 缺失均将导致糖原代 谢紊乱并最终引发Ia 型糖原贮积病
内质网腔
Glc质膜载体 毛 细 血 管
H13.4
§2. 糖原合成
- High [Pi] in cell favors glycogen breakdown & prevents from glycogen synthesis in vivo.