人肺癌相关成纤维细胞永生化
上皮间质转化在肺癌中的研究进展

上皮间质转化在肺癌中的研究进展张琨琨【期刊名称】《检验医学与临床》【年(卷),期】2012(009)021【总页数】2页(P2726-2727)【关键词】上皮间质转化;肺癌;肿瘤干细胞;转移【作者】张琨琨【作者单位】第三军医大学第二附属医院,重庆,400037【正文语种】中文肺癌是全球新发病例和死亡病例均居首位的恶性肿瘤,影响患者预后的主要因素是出现局部复发及远处转移。
新近研究表明,上皮间质转化(EMT)在肿瘤演进中是一个重要的决定性过程[1-2],与肿瘤细胞侵袭、转移乃至“干性”等密切相关。
1 EMT概念EMT是上皮细胞失去上皮特性获得间质细胞表型的一种生物现象。
EMT发生后,细胞间连接蛋白及上皮特异性蛋白表达下降,如 E-钙黏蛋白(E-cad)、β-连锁蛋白等;同时伴随间充质细胞特异性蛋白表达增高,如波形蛋白(vimentin)、N-钙黏蛋白(N-cad)等。
EMT在人胚胎干细胞分化过程中起着关键作用,这个过程在组织修复再生、器官纤维化和肿瘤发生时可被病理性激活[3]。
肿瘤细胞是否存在EMT现象,曾一度出现争论。
在一项乳腺癌研究中,研究者利用间质和上皮细胞特异性表达Cre重组酶转基因小鼠模型,首次提供了肿瘤细胞在体内演进中发生EMT的直接证据[4],从而使EMT从理论推测逐步走向肿瘤转化医学研究。
2 EMT相关信号通路EMT可被多种信号分子所诱导,如β生长转化因子(TGFβ)、肝细胞生长因子、成纤维细胞生长因子(FGF)以及Wnt、Notch与hedgehog通路蛋白等。
TGFβ是当前研究最广泛的EMT诱导因子,其与受体结合后,将发放跨膜信号,直接磷酸化C-末端激活Smad2和Smad3,后二者与Smad4形成三聚体转移进入核内,进而通过与DNA转录结合位点作用调控TGFβ应答基因表达[5]。
但是,由于Smad分子与DNA亲和力较低,这一过程中还需要Snail、ZEB等转录因子的协同作用,以获得对应答基因更高的亲和力和选择性。
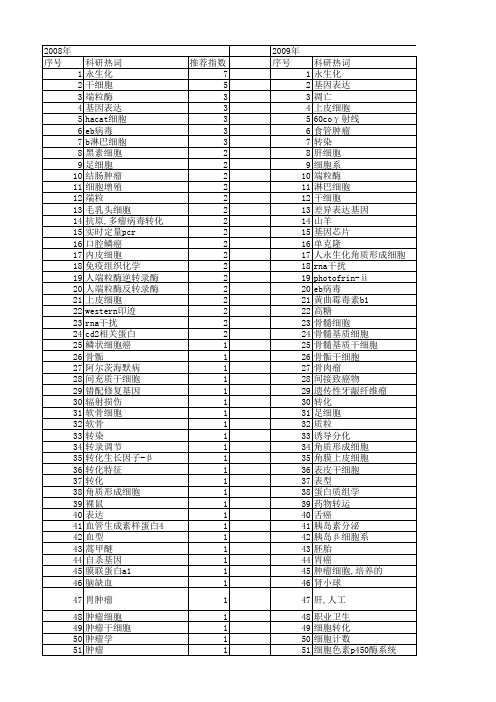
【国家自然科学基金】_永生化_基金支持热词逐年推荐_【万方软件创新助手】_20140730
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106
细胞系shee 细胞恶性转化 细胞培养 细胞分化 组织工程皮肤 组织工程 线粒体 紫外线(uvb) 糖尿病肾病 糖尿病 种子细胞 离体角膜 神经干细胞 碱性成纤维细胞生长因子 砷中毒 砷 石英 白三烯b4 癌,肝细胞 甲基化 生长抑制 琼脂糖凝胶电泳 环孢霉素a 猿肾病毒40 猴病毒40 淋巴结转移 流式细胞术 永生细胞系 永生化细胞系 永生化细胞 永生化淋巴母细胞系 氧化应激 栽体 核型分析 无饲养层培养 整合素β 1 放射性脑损伤 支气管 抗原,病毒,肿瘤 抑制性消减杂交 成纤维细胞 慢病毒属 恶性转化 差异表达 子宫内膜基质细胞 姜黄素 大鼠 外周血淋巴细胞 外周血单核细胞 基因表达谱 基因表达调控,肿瘤 基因治疗 基因,haph2β 基因,c-abl
模式生物 杀伤细胞,天然 星形细胞 整合素类 整合素α ν β 6 抗原 抑癌基因 微卫星不稳定性 强力霉素 幽门螺杆菌 小干扰 实验性 学习记忆 姜黄素 天然高本底地区 大鼠 基质金属蛋白酶类 基质细胞 基因表达调控 基因芯片 基因疗法 基因治疗 基因,ras 基因,myc 基因,htert 基因 培养的 四甲基偶氮唑蓝法 喉癌 启动子区 半乳糖转移酶类 半乳糖苷酶类 分离培养 免疫荧光 克隆形成 侵袭 传代 人骨髓间充质干细胞 人类白细胞抗原dr 人源性永生化肝细胞 乙型肝炎病毒x蛋白 乙型肝炎病毒 中波紫外线 wt1 synaptopodin sv40t抗原 sv40t基因 sv40t st6galnac2 sp细胞 smad4 sirna rna pig3
铁死亡与自噬在疾病中的研究进展

㊀㊀ʌ摘㊀要ɔ㊀铁死亡是一种铁依赖的㊁以谷胱甘肽过氧化物酶4活性丧失㊁脂质过氧化物沉积为特点的细胞死亡方式ꎮ自噬是一种高度保守的㊁用于降解和回收利用生物大分子或受损细胞器的过程ꎮ当自噬过度激活时ꎬ也会引起细胞的自噬性死亡ꎮ文章就铁死亡与自噬的相互关系及其在神经系统疾病㊁循环系统疾病和肿瘤中的研究进展作一综述ꎬ以期加深对铁死亡及自噬关系的认识ꎮʌ关键词ɔ㊀铁死亡ꎻ自噬ꎻ相互关系ꎻ铁离子ʌDOIɔ㊀10.3969/j.issn.1671 ̄6450.2019.12.023Researchprogressofirondeathandautophagyindiseases㊀PENGXiaꎬFANGCongcongꎬXUYuming.DepartmentofPediatricsꎬRenminHospitalofWuhanUniversityꎬHubeiProvinceꎬWuhan430060ꎬChinaCorrespondingauthor:YAOBaozhenꎬE ̄mail:professoryao@aliyun.com㊀㊀ʌAbstractɔ㊀Irondeathisaniron ̄dependentcelldeathmodecharacterizedbylossofglutathioneperoxidase4activityandlipidperoxidedeposition.Autophagyisahighlyconservativeprocessusedtodegradeandrecyclebiologicalmacromole ̄culesordamagedorganelles.Whenautophagyisoveractivatedꎬautophagicdeathofcellsisalsocaused.Thisarticlereviewstherelationshipbetweenirondeathandautophagyanditsresearchprogressinnervoussystemdiseasesꎬcirculatorysystemdis ̄easesandtumorsinordertodeepentheunderstandingoftherelationshipbetweenirondeathandautophagy.㊀㊀ʌKeywordsɔ㊀FerroptosisꎻAutophagyꎻCorrelationꎻIronion㊀㊀早在铁死亡被命名之前ꎬ研究者们就已经发现了erastin㊁RSL3可以诱导细胞以一种不同于凋亡的方式死亡ꎬ且这种细胞死亡方式可以被铁螯合剂及抗氧化剂抑制ꎬ从而明确了这种细胞死亡方式与细胞内铁和活性氧(ROS)有关ꎮ2012年ꎬDixon等[1]发现这种细胞死亡方式在形态学㊁生化和遗传学上均与凋亡㊁坏死及自噬不同ꎮ由于这种细胞死亡方式特异地依赖于细胞内铁ꎬ故将其命名为铁死亡ꎮ铁死亡的机制尚未完全阐明ꎬ研究发现铁死亡存在复杂的细胞内调控机制ꎬ铁代谢㊁氨基酸代谢及脂质代谢都参与其中ꎮ自噬广泛存在于真核生物中ꎬ是依赖于细胞内溶酶体分解衰老或损伤的大分子或细胞器的过程ꎬ从而维持细胞内稳态[2]ꎮ然而ꎬ当细胞面对各种不利因素刺激时ꎬ自噬会过度激活ꎮ过度激活的自噬可以导致细胞发生自噬性死亡[3]ꎮ铁死亡和自噬作为2种不同的生物过程ꎬ在细胞内发挥着各自重要的作用ꎮ在铁死亡发现之初ꎬ研究者们认为这是一种与自噬截然不同的细胞死亡方式ꎮ近年来ꎬ虽然仍然存在很多争议ꎬ但是越来越多的研究在逐渐揭示铁死亡与自噬的关系ꎬ并在不同的疾病中证实ꎮ1㊀铁死亡的发生机制1.1㊀细胞内铁与铁死亡㊀铁作为生命必需的微量元素之一ꎬ主要以二价和三价铁离子的形式存在于机体内ꎮ正常情况下ꎬ小肠吸收或红细胞降解释放出的亚铁离子(Fe2+)被氧化为三价铁离子(Fe3+)ꎬFe3+经膜上转铁蛋白(transferrinꎬTF)㊁膜蛋白转铁蛋白受体1(transferrinreceptorprotein1ꎬTFR1)的作用后内吞入胞体ꎮ内吞入细胞内的Fe3+被还原成亚铁离子Fe2+ꎬ然后由二价金属离子转运蛋白1(divalentmetaltrans ̄porter1ꎬDMT1)或锌铁调控蛋白家族8/14(ZRT/IRT ̄likeproteins8/14ꎬZIP8/14)介导Fe2+储存到细胞内不稳定的铁池(labileironpoolꎬLIP)中和铁蛋白轻链多肽(ferritinlightchainꎬFTL)与铁蛋白重链多肽1(ferritinheavychain1ꎬFTH1)组成的铁储存蛋白复合物中ꎮ剩余部分亚铁离子将被氧化成Fe3+出胞ꎬ参与体内铁再循环ꎬ严格把控细胞内铁稳态ꎮ㊀㊀铁参与了生物体很多重要的功能ꎬ包括新陈代谢㊁氧转运㊁抗氧化反应及DNA合成等一系列生物过程ꎮFe ̄S是线粒体电子传递链中氧化还原酶类的重要辅助因子ꎬ如NADH㊁辅酶Q等ꎮ活性氧(reactiveoxygenspeciesꎬROS)的产生与铁稳态的扰动密切相关ꎬ而铁硫团簇(iron ̄sulfurclusterꎬISC)机械系统功能障碍明显加剧了ROS的产生[4]ꎮ然而ꎬ细胞内过多的铁通过芬顿反应产生羟自由基ꎬ从而促进不饱和脂肪酸(PU ̄FAs)的羟化ꎬ由此产生的脂质过氧化物和氢过氧化物严重影响细胞膜的结构和功能[5]ꎬ这个过程就是铁死亡ꎮ铁死亡过程可以被铁螯合剂阻断ꎬ说明细胞内铁与铁死亡密切相关[6]ꎮ而这些有毒的脂质过氧化物ꎬ在谷胱甘肽和谷胱甘肽过氧化物酶4(GPX4)的作用下才可以转变为为无毒的醇类物质ꎬ从而避免其对细胞的杀伤作用[7]ꎮ㊀㊀虽然细胞内铁与铁死亡密切相关ꎬ但是细胞内铁在铁死亡中的具体作用机制至今仍不明确ꎮDixon等[8]认为铁螯合剂抑制铁死亡最有可能的解释是阻止了铁向氧化物传递电子ꎬ从而抑制活性氧的生成ꎮ也有报道提出ꎬ铁螯合剂能够直接作用于含铁离子的酶ꎬ其中以脂氧合酶的可能性最大[9]ꎮ1.2㊀XC ̄系统与铁死亡㊀胱氨酸 谷氨酸反向转运系统(XC ̄系统)是一个广泛分布于磷脂双分子层的氨基酸逆向转运体ꎬ由轻链xCT(SLC7A11)和重链4F2(SLC3A2)组成[10]ꎮ通过XC ̄系统ꎬ胱氨酸与谷氨酸(GLu)以1ʒ1的比例在细胞内外进行交换ꎮ在细胞内ꎬ胱氨酸被谷胱甘肽(GSH)或硫氧还蛋白还原酶1还原为半胱氨酸ꎬ在γ ̄谷氨酰半胱氨酸合成酶和谷胱甘肽合成酶的作用下进一步合成GSH[11]ꎮ细胞摄取半胱氨酸是谷胱甘肽合成的关键步骤ꎬGSH的产生和维持对保护细胞免受氧化应激反应所造成的损伤至关重要ꎬ而半胱氨酸的含量需要转硫途径的调节[12 ̄13]ꎮKang等[14]发现高浓度Glu孵育神经细胞可以抑制XC ̄系统ꎬ从而抑制胱氨酸的摄入ꎬ引起细胞内谷胱甘肽减少和ROS的聚集ꎬ而抑制铁死亡可以抑制Glu引起的神经元死亡ꎮ这些研究都表明XC ̄系统参与了铁死亡过程ꎮ当XC ̄系统被抑制时ꎬ胱氨酸不能转入细胞内ꎬ谷胱甘肽合成减少ꎬ不能将有毒的脂质过氧化物还原成无毒的醇类物质ꎬ进而诱导了铁死亡的发生ꎮ1.3㊀GPX4与铁死亡㊀抑制XC ̄系统导致GSH减少和活性氧的聚集ꎮ当XC ̄系统功能正常且GSH正常合成时ꎬGSH也必须在GPX4的作用下将有毒的脂质过氧化物还原成无毒的醇类物质ꎮGPX4是铁死亡关键的调节因子ꎬ在预防脂质过氧化中起着至关重要的作用[15]ꎮ研究发现ꎬGPX4是唯一一种能够降低生物膜内脂质过氧化氢的酶ꎬ所以当GPX4功能受限时ꎬGSH并未耗竭ꎬ但脂质ROS明显升高ꎻ其次ꎬGSH是GPX4活性的一个必要的辅助因子[16]ꎮGPX4被抑制时将导致脂质ROS的形成及脂质过氧化ꎬ最后诱导铁死亡的发生[17]ꎮ1.4㊀脂质过氧化物与铁死亡㊀不饱和脂肪酸在氧存在情况下很容易发生脂质过氧化ꎬ这种过氧化反应在铁的存在下会加剧[18]ꎬ其中与铁死亡密切相关的不饱和脂肪酸包括花生四烯酸和肾上腺酸ꎮ酯酰基辅酶A合成酶长链家族成员4(AcylCoAsynthetaselong ̄chainfamilymember4ꎬACSL4)和脂质重塑相关的溶血卵磷脂酰基转移酶3(lysophosphatidyl ̄cholineacyl ̄transferase3ꎬLPCAT3)是参与脂质过氧化物形成的2种关键酶ꎬ在它们的作用下ꎬ不饱和脂肪酸转化成脂质过氧化物[19 ̄20]ꎮ脂质过氧化的积累会不可避免地会造成很大的损伤ꎮ丙二醛(MDA)是活性氧作用于生物膜不饱和脂肪酸而产生的脂质过氧化反应的最终产物ꎮ其积累可引起蛋白质与核酸的交联聚合ꎬ导致膜结构的不可逆破坏ꎬ最终导致细胞死亡[21]ꎮ2㊀自噬的过程㊀㊀相较于铁死亡ꎬ自噬的研究则更深入ꎮ目前研究认为ꎬ自噬主要有3种形式:(1)巨自噬ꎬ细胞内损坏的蛋白质㊁细胞器及胞内病原体等被细胞质产生的膜结构包裹ꎬ形成自噬体ꎬ最终与溶酶体融合后被降解ꎻ(2)微自噬ꎬ被降解物直接被溶酶体通过变形运动进行内吞ꎬ使其降解ꎬ这个过程不形成自噬体ꎻ(3)分子伴侣介导的自噬ꎬ待降解物需要与分子伴侣结合ꎬ然后被溶酶体上的溶酶体相关膜蛋白(lysosomalassociatedmembraneproteinꎬLAMPs)识别ꎬ并最终被溶酶体降解[22]ꎮ目前研究较多的自噬类型为巨自噬(以下简称为 自噬 )ꎮ自噬是一个复杂的过程ꎬ具体包括4个步骤:自噬的诱发ꎻ隔膜的延伸㊁闭合ꎬ形成自噬体ꎻ自噬体与溶酶体结合ꎬ形成自噬溶酶体ꎻ自噬体和内部物质的降解[23]ꎮ㊀㊀随着对自噬研究的深入ꎬ人们发现自噬也是有选择性的ꎮ细胞内存在这一些特殊的自噬ꎬ它们在某些特定的条件下对某种大分子或者细胞器进行特定的降解[24]ꎬ如线粒体自噬㊁内质网自噬㊁过氧化物酶体自噬㊁核糖体自噬和脂类自噬ꎮ3㊀铁死亡与自噬的关系㊀㊀铁死亡在发现之初ꎬ人们认为它是一种在生化㊁形态及基因水平与凋亡㊁坏死及自噬不同的细胞死亡途径[25]ꎬ然而随着研究的不断推进ꎬ越来越多的证据表明铁死亡的发生需要自噬机制的参与[26]ꎮHou等[27]通过在永生化小鼠胚胎成纤维细胞(MEFs)㊁人胰腺癌细胞系(PANC1和PANC2.03)和人纤维肉瘤细胞系HT ̄1080中敲低ATG5和ATG7抑制自噬后发现细胞内游离铁水平和脂质过氧化终产物(如MDA)水平均显著下降ꎬ而细胞内稳定铁蛋白标志物FTH1的表达显著上升ꎬ该研究首次从基因层面揭示了自噬和铁死亡关系ꎮZhou等[28]通过实验进一步证实了这种关系ꎬ发现这种特殊的自噬过程是以铁蛋白为底物的ꎮ之后ꎬ许多研究证实与铁死亡密切相关的大分子物质参与自噬的发生过程中ꎮGSH㊁GPX4及脂质过氧化物都是铁死亡过程中关键的大分子物质ꎬ研究表明在饥饿和氧化应激等条件下ꎬ自噬发生时伴随着GSH的下降[29]ꎻ而GPX4过表达可抑制ROS介导的自噬的发生ꎬ脂质过氧化物可以促进自噬体的形成[30 ̄31]ꎮ核受体辅激活因子4(NCOA4)是铁自噬过程中的选择性受体[32]ꎬ而下调NCOA4可以抑制erastin诱导的铁死亡ꎮ脂质自噬介导的脂滴降解可以促进铁死亡的发生[33]ꎮ其他如SLC7A11㊁NRF2㊁p53㊁HSPB1和ACSL4等铁死亡调控因子已经被证实是自噬的潜在调控因子ꎮ目前越来越多的研究证实过度的自噬可以促进铁死亡ꎮ然而ꎬ也有一些学者通过实验证实ꎬ铁死亡的发生可独立于自噬ꎮ4㊀铁死亡与自噬在疾病中的研究4.1㊀铁死亡与自噬在神经系统疾病中的研究㊀蛛网膜下腔出血(subarachnoidhemorrhageꎬSAH)是神经系统疾病中重要的一种ꎮ梁译丹等[33]研究证实ꎬ在SAH的大鼠模型中ꎬ通过侧脑室注射慢病毒沉默ATG5mRNA的表达从而抑制自噬ꎮ与单纯SAH组相比ꎬ慢病毒干预组PCR显示ATG5mRNA显著降低ꎻWestern ̄blot检测结果显示ATG5和LC3Ⅱ/Ⅰ蛋白表达量显著降低(P<0.05)ꎬ表明自噬被成功抑制ꎮ与此同时ꎬ与SAH组相比ꎬ慢病毒干预组的铁死亡标志物FTH1和GPX4表达升高(P<0.05)ꎬGSH含量提高ꎬ细胞内铁沉积减少ꎬ铁含量㊁MDA减少ꎮ上述研究结果表明自噬通过降解铁蛋白ꎬ促进铁死亡ꎮ该研究认为铁死亡的发生依赖于自噬的介导ꎬ在这之中铁蛋白发挥重要的中介作用ꎮ与之前的研究结果一致[28 ̄29]ꎬ即自噬通过降解铁蛋白ꎬ增加细胞内游离铁离子的浓度ꎬ从而促进铁死亡ꎮ这项研究为铁死亡和自噬的相关性提供了实验依据ꎮ4.2㊀铁死亡与自噬在循环系统疾病中的研究㊀Chen等[34]研究证实ꎬ在心力衰竭模型中ꎬ自噬铁死亡是同时发生的ꎮ激活的自噬与铁死亡与心力衰竭的发生发展密切相关ꎮ而Baba等[35]在心肌梗死的研究中得出了不同的结论ꎬ认为雷帕霉素机制靶点(mTOR)可以抑制心脏细胞中铁死亡的发生ꎬ从而发挥保护性作用ꎮ与此同时ꎬmTOR介导的对铁死亡的抑制过程中并没有伴随着自噬标志物的改变ꎬ这提示铁死亡可独立于自噬发生ꎮ4.3㊀铁死亡与自噬在肿瘤中的研究㊀香蒲新苷(typhaneoside)是一种可显著降低急性髓系白血病(acutemyeloidleukemiaꎬAML)细胞活力的药物ꎮ研究表明ꎬ香蒲新苷可通过作用于AMP活化蛋白激酶(AMP ̄activatedproteinkinaseꎬAMPK)信号通路激活自噬ꎬ从而进一步激活自噬依赖的铁蛋白的降解ꎬ最终诱导铁死亡的发生ꎬ而抑制自噬后则可以抑制这种细胞死亡方式[36]ꎮ这充分说明在香蒲新苷促进AML细胞发生铁死亡的过程是依赖于自噬的激活的ꎮ该研究再次为铁死亡和自噬的相互关系提供了充分的实验依据ꎮ而铁死亡与自噬在乳腺癌中的研究得出了不一样的结论ꎮ联合使用西拉米辛和拉帕替尼可以显著诱导乳腺癌细胞发生铁死亡ꎬ也可诱导细胞发生自噬性死亡ꎮ但是Ma等[37]通过实验证实这种联合用药诱导乳腺癌细胞发生自噬性死亡在时间上是落后于铁死亡的ꎮ这表明在西拉米辛和拉帕替尼治疗乳腺癌的过程中ꎬ铁死亡与自噬是独立发生的ꎮ铁死亡的发生并不总是依赖于自噬ꎬ铁死亡与自噬的关系仍未明确ꎬ其内在联系更为复杂ꎬ需要学者们更多的研究与探索ꎮ5㊀小㊀结㊀㊀铁死亡作为一种新型的细胞死亡方式ꎬ与多种疾病的发生发展密切相关ꎮ自噬作为一种保守的大分子或细胞器的降解过程ꎬ在病理情况下也会导致细胞的死亡ꎮ关于铁死亡与自噬的关系ꎬ目前尚无统一结论ꎮ虽然越来越多的研究证实铁死亡的发生伴随着自噬的激活ꎬ但是也有不少的研究表明铁死亡可独立于自噬的激活而存在ꎮ铁死亡与自噬的关系目前仍有许多未知的问题ꎬ且它们在疾病中的关系也处在研究的初级阶段ꎬ未来将在进一步的研究中深入揭示两者间的联系ꎮ参考文献[1]㊀DixonSꎬLembergKꎬLamprechtMꎬetal.Ferroptosis:AnIron ̄De ̄pendentFormofNonapoptoticCellDeath[J].Cellꎬ2012ꎬ149(5):1060 ̄1072.DO:10.1016/j.cell.2012.03.042.[2]㊀TurcoEꎬFracchiollaDꎬMartensS.RecruitmentandActivationoftheULK1/Atg1KinaseComplexinSelectiveAutophagy[J].JMolBi ̄olꎬ2019ꎬPii:S0022 ̄2836(19)30471 ̄1.DOI:10.1016/j.jmb.2019.07.027.[3]㊀HouJꎬRaoMꎬZhengWꎬetal.AdvancesonCellAutophagyandItsPotentialRegulatoryFactorsinRenalIschemia ̄ReperfusionInjury[J].DNACellBiolꎬ2019ꎬ38(9):895 ̄904.DOI:10.1089/dna.2019.4767.[4]㊀YangLꎬMihNꎬAnandAꎬetal.Cellularresponsestoreactiveoxygenspeciesarepredictedfrommolecularmechanisms[J].ProcNatlAcadSciUSAꎬ2019ꎬ116(28):14368 ̄14373.DO:10.1073/pnas.1905039116.[5]㊀ZhuTꎬShiLꎬYuCꎬetal.FerroptosisPromotesPhotodynamicThera ̄py:SupramolecularPhotosensitizer ̄InducerNanodrugforEnhancedCancerTreatment[J].Theranosticsꎬ2019ꎬ9(11):3293 ̄3307.DOI:10.7150/thno.32867.[6]㊀YuHꎬGuoPꎬXieXꎬetal.Ferroptosisꎬanewformofcelldeathꎬanditsrelationshipswithtumourousdiseases[J].JournalofCellularandMolecularMedicineꎬ2016.21(4):648 ̄657.DOI:10.1111/jcmm.13008.[7]㊀GongYꎬWangNꎬLiuNꎬetal.LipidPeroxidationandGPX4Inhibi ̄tionAreCommonCausesforMyofibroblastDifferentiationandFer ̄roptosis[J].DNACellBiolꎬ2019ꎬ38(7):725 ̄733.DOI:10.1089/dna.2018.4541.[8]㊀DixonSJꎬStockwellBR.Theroleofironandreactiveoxygenspeciesincelldeath[J].NatureChemicalBiologyꎬ2013ꎬ10(1):9 ̄17.DOI:10.1038/nchembio.1416.[9]㊀WangHꎬLiJꎬFollettPLꎬetal.12 ̄Lipoxygenaseplaysakeyroleincelldeathcausedbyglutathionedepletionandarachidonicacidinratoligodendrocytes[J].EuropeanJournalofNeuroscienceꎬ2004ꎬ20(8):2049 ̄2058.DOI:10.1111/j.1460 ̄9568.2004.03650.x. [10]㊀PitmanKEꎬAlluriSRꎬKristianAꎬetal.Influxrateof18F ̄fluoroamin ̄osubericacidreflectscystine/glutamateantiporterexpressionintumourxenografts[J].EurJNuclMedMolImagingꎬ2019ꎬ40(10):2190 ̄2198.DOI:10.1007/s00259 ̄019 ̄04375 ̄8.[11]㊀BridgesRJꎬNataleNRꎬPatelSA.Systemxc( ̄)cystine/glutamateantiporter:anupdateonmolecularpharmacologyandroleswithintheCNS[J].BrJPharmacolꎬ2012ꎬ165(1):20 ̄34.DOI:10.1111/j.1476 ̄5381.2011.01480.x.[12]㊀WangLꎬLiuYꎬDuTꎬetal.ATF3promoteserastin ̄inducedferropto ̄sisbysuppressingsystemXc[J].CellDeathDifferꎬ2019.DOI:10.1038/s41418 ̄019 ̄0380 ̄z.[13]㊀YuHꎬGuoPꎬXieXꎬetal.Ferroptosisꎬanewformofcelldeathꎬanditsrelationshipswithtumourousdiseases[J].JournalofCellularandMolecularMedicineꎬ2016ꎬ21(4):648 ̄657.DOI:10.1111/jcmm.13008.[14]㊀KangYꎬTizianiSꎬParkGꎬetal.CellularprotectionusingFlt3andPI3Kαinhibitorsdemonstratesmultiplemechanismsofoxidativeglu ̄tamatetoxicity[J].NatureCommunicationsꎬ2014ꎬ5:3672.DOI:10.1038/ncomms4672.[15]㊀ForcinaGCꎬDixonSJ.GPX4attheCrossroadsofLipidHomeostasisandFerroptosis[J].Proteomicsꎬ2019ꎬ19(18):e1800311.DOI:10.1002/pmic.201800311.[16]㊀Brigelius ̄FloheRꎬMaiorinoM.Glutathioneperoxidases[J].BiochimBiophysActaꎬ2013ꎬ1830(5):3289 ̄3303.DOI:10.1016/j.bba ̄gen.2012.11.020.[17]㊀ForcinaGCꎬDixonSJ.GPX4attheCrossroadsofLipidHomeostasisandFerroptosis[J].Proteomicsꎬ2019ꎬ19(18):e1800311.DOI:10.1002/pmic.201800311.[18]㊀PrattDAꎬTallmanKA.Freeradicaloxidationofpolyunsaturatedlip ̄ids:Newmechanisticinsightsandthedevelopmentofperoxylradicalclocks[J].AccChemResꎬ2011ꎬ44(6):458 ̄467.DOI:10.1021/ar200024c.[19]㊀DixonSJꎬWinterGEꎬMusaviLSꎬetal.HumanHaploidCellGeneticsRevealsRolesforLipidMetabolismGenesinNonapoptoticCellDeath[J].ACSChemicalBiologyꎬ2015ꎬ10(7):1604 ̄1609.DOI:10.1021/acschembio.5b00245.[20]㊀CongcongFꎬLijuanGꎬDanielSꎬetal.TheInterrelationbetweenRe ̄activeOxygenSpeciesandAutophagyinNeurologicalDisorders[J].OxidativeMedicineandCellularLongevityꎬ2017ꎬ2017:8495160.DOI:10.1155/2017/8495160.[21]㊀QomaladewiNPꎬKimMYꎬChoJY.Autophagyanditsregulationbyginsengcomponents[J].JGinsengResꎬ2019ꎬ43(3):349 ̄353.DO:10.1016/j.jgr.2018.12.011.[22]㊀杨静亚ꎬ高俊玲.自噬及其在疾病中的研究进展[J].世界最新医学信息文摘ꎬ2019ꎬ19(52):29 ̄30ꎬ34.DOI:10.19613/j.cnki.1671 ̄3141.2019.52.016.[23]㊀JohansenTꎬLamarkT.SelectiveAutophagy:ATG8FamilyProteinsꎬLIRMotifsandCargoReceptors[J].JMolBiolꎬ2019ꎬpiiꎬS0022 ̄2836(19):30445.DOI:10.1016/j.jmb.2019.07.016.[24]㊀YuHꎬGuoPꎬXieXꎬetal.Ferroptosisꎬanewformofcelldeathꎬanditsrelationshipswithtumourousdiseases[J].JournalofCellularandMolecularMedicineꎬ2016ꎬ21(4):648 ̄657.DOI:10.1111/jcmm.13008.[25]㊀KangRꎬTangD.AutophagyandFerroptosis ̄What'stheConnection[J].CurrentPathobiologyReportsꎬ2017ꎬ5(2):153 ̄159.DOI:10.1007/s40139 ̄017 ̄0139 ̄5.[26]㊀GaoMꎬMonianPꎬPanQꎬetal.Ferroptosisisanautophagiccelldeathprocess[J].Cellresearchꎬ2016ꎬ26(9):1021 ̄1032.DOI:10.1038/cr.2016.95.[27]㊀HouWꎬXieYꎬSongXꎬetal.Autophagypromotesferroptosisbydeg ̄radationofferritin[J].Autophagyꎬ2016ꎬ12(8):1425 ̄1428.DOI:10.1080/15548627.2016.1187366[28]㊀ZhouQꎬFuXꎬWangXꎬetal.AutophagyplaysaprotectiveroleinMn ̄inducedtoxicityinPC12cells[J].Toxicologyꎬ2017ꎬ394:45 ̄53.DOI:10.1016/j.tox.2017.12.001.[29]㊀GargADꎬDudekAMꎬFerreiraGBꎬetal.ROS ̄inducedautophagyincancercellsassistsinevasionfromdeterminantsofimmunogeniccelldeath[J].Autophagyꎬ2013ꎬ9(9):1292 ̄307.DOI:10.4161/auto.25399.[30]㊀HillBGꎬHaberzettlPꎬAhmedYꎬetal.Unsaturatedlipidperoxida ̄tion ̄derivedaldehydesactivateautophagyinvascularsmooth ̄musclecells[J].BiochemicalJournalꎬ2008ꎬ410(3):525 ̄534.DOI:10.1042/BJ20071063.[31]㊀QuilesDelReyMꎬManciasJD.NCOA4 ̄MediatedFerritinophagy:APotentialLinktoNeurodegeneration[J].FrontNeurosciꎬ2019ꎬ13:238.DOI:10.3389/fnins.2019.00238.[32]㊀MouYꎬWangJꎬWuJꎬetal.Ferroptosisꎬanewformofcelldeath:op ̄portunitiesandchallengesincancer[J].JournalofHematology&Oncologyꎬ2019ꎬ12(1):34.DOI:10.1186/s13045 ̄019 ̄0720 ̄y.(下转1292页)10.011.[12]㊀BrazNFTꎬRochaNPꎬVieiraELMꎬetal.Musclestrengthandpsychi ̄atricsymptomsinfluencehealth ̄relatedqualityoflifeinpatientswithmyastheniagravis[J].JClinNeurosciꎬ2018ꎬ50:41 ̄44.DOI:10.1016/j.jocn.2018.01.011.[13]㊀GuptillJTꎬSoniMꎬMeriggioliMN.CurrentTreatmentꎬEmergingTranslationalTherapiesꎬandNewTherapeuticTargetsforAutoim ̄muneMyastheniaGravis[J].Neurotherapeuticsꎬ2016ꎬ13(1):118 ̄131.DOI:10.1007/s13311 ̄015 ̄0398 ̄y.[14]㊀DhawanPSꎬGoodmanBPꎬHarperCMꎬetal.IVIGVersusPLEXintheTreatmentofWorseningMyastheniaGravis:WhatistheEvi ̄dence[J].ACriticallyAppraisedTopicNeurologistꎬ2015ꎬ19(5):145 ̄148.DOI:10.1097/NRL.0000000000000026.[15]㊀刘敏ꎬ邢昂ꎬ丛志强.血浆交换与大剂量丙种球蛋白治疗重症肌无力危象疗效的评价[J].中华神经科杂志ꎬ2005ꎬ10(38):658 ̄659.DOI:10.3760/j.issn:1006 ̄7876.2005.10.016.[16]㊀GamezJꎬSalvadoMꎬCasellasMꎬetal.Intravenousimmunoglobu ̄linasmonotherapyformyastheniagravisduringpregnancy[J].Jour ̄naloftheNeurologicalSciencesꎬ2017ꎬ383:118 ̄122.DOI:10.1016/j.jns.2017.10.037.[17]㊀BharathVꎬEckertKꎬKangMꎬetal.Incidenceandnaturalhistoryofintravenousimmunoglobulin ̄inducedasepticmeningitis:aretrospec ̄tivereviewatasingletertiarycarecent[J].Transfusionꎬ2015ꎬ55(11):2597 ̄2605.DOI:10.1111/trf.13200.[18]㊀StetefeldHRꎬSchroeterM.MyastheneCrisis[J].FortchrNeurolPsychiatrꎬ2018ꎬ86(5):301 ̄307.DOI:10.1055/a ̄0599 ̄0811. [19]㊀黄玲ꎬ王磊ꎬ尹东涛ꎬ等.环磷酰胺序贯治疗伴胸腺瘤的重症肌无力50例临床分析[J].神经疾病与精神卫生ꎬ2018ꎬ18(8):583 ̄586.DOI:10.3969/j.issn.1009 ̄6574.2018.08.012.[20]㊀QiGꎬLiuPꎬDongHꎬetal.MetastaticThymoma ̄AssociatedMyas ̄theniaGravis:FavorableResponsetoSteroidPulseTherapyPlusIm ̄munosuppressiveAgent[J].MedicalScienceMonitorꎬ2017ꎬ23ꎬ1217 ̄1223.DOI:10.12659/MSM.902442.[21]㊀冯慧宇ꎬ刘卫彬ꎬ邱力ꎬ等.中剂量环磷酰胺联合甲泼尼龙治疗重症肌无力危象的随机对照临床研究[J].中华医学杂志ꎬ2012ꎬ92(35):2473 ̄2476.DOI:10.3760/cma.j.issn.0376 ̄2491.2012.35.007.[22]㊀RodaRHꎬDohertyLꎬCorseAM.stoppingoralsteroidsparingagentatinitiationofrituximabinmyastheniagravis[J].NeuromusculDis ̄ordꎬ2019ꎬ29(7):554 ̄561.DOI:10.1016/j.nmd.2019.06.002. [23]㊀IorioRꎬDamatoVꎬAlboiniPEꎬetal.Efficacyandsafetyofritux ̄imabformyastheniagravis:asystematicreviewandmetaanalysis[J].JNeurolꎬ2015ꎬ262(5):1115 ̄1119.DOI:10.1007/s00415 ̄014 ̄7532 ̄3.[24]㊀KoulRꎬFutaisiAꎬAbdwaniR.Rituximabinsevereseronegativeju ̄venilemyastheniagravis:reviewoftheliterature[J].PediatriNeu ̄rolꎬ2012ꎬ47(3):209 ̄212.DOI:10.1016/j.pediatrneurol.2012.05.017.[25]㊀JingSꎬSongYꎬSongJꎬetal.Responsivenesstolowdoserituximabinrefractorygeneralizedmyastheniagravis[J].JNeuroimrnunolꎬ2017ꎬ311:14 ̄21.DOI:10.1016/j.jneuroim.2017.05.021. [26]㊀TopakianRꎬZimprichFꎬIglsederSꎬetalꎬHighefficacyofrituximabformyastheniagravis:acomprehensivenationwidestudyinAustria[J].JNeurolꎬ2019ꎬ266(3):699 ̄706.DOI:10.1007/s00415 ̄019 ̄09191 ̄6.[27]㊀OnukiTꎬUedaSꎬOtsuSꎬetal.ThymectomyduringMyasthenicCrisisunderArtificialRespiration[J].AnnThoracCardiovascSurgꎬ2019ꎬ25(4):215 ̄218.DOI:10.5761/atcs.cr.17 ̄00176.[28]㊀薛银萍ꎬ乞国艳ꎬ董会民ꎬ等.参芪扶正注射液在重症肌无力激素冲击治疗过程中的应用[J].广东医学ꎬ2016ꎬ37(24):3764 ̄3756.DOI:10.3969/j.issn.1001 ̄9448.2016.24.035.[29]㊀徐鹏ꎬ吕志国ꎬ王健ꎬ等.重症肌无力中医循证性临床诊疗指南修订实践研究[J].中华中医药杂志ꎬ2018ꎬ33(5):1979 ̄1983.DOI:CNKI:SUN:BXYY.0.2018 ̄05 ̄066.[30]㊀吴周烨ꎬ吴颢昕ꎬ何骁隽ꎬ等.益气升提法治疗实验性自身免疫性重症肌无力大鼠免疫机制研究[J].中华中医药杂志ꎬ2017ꎬ32(6):2746 ̄2749.DOI:CNKI:SUN:BXYY.0.2017 ̄06 ̄112. [31]㊀方学君ꎬ梁艺ꎬ刘晓曼ꎬ等.强肌健力方对重症肌无力大鼠Th17/Treg平衡的影响[J].广州中医药大学学报ꎬ2019ꎬ36(2):240 ̄245.DOI:10.13359/j.cnki.gzxbtcm.2019.02.019.[32]㊀王艳君ꎬ孟庆芳ꎬ王思青ꎬ等.蒿素对实验性自身免疫性重症肌无力大鼠R97 ̄116抗体及细胞因子的影响[J].中国神经免疫学和神经病学杂志ꎬ2016ꎬ23(3):167 ̄170.DOI:10.3969/J.issn.1006 ̄2963.2016.03.002.(收稿日期:2019-06-14)(上接1287页)[33]㊀梁译丹ꎬ覃王ꎬ黄豪ꎬ等.自噬通过降解铁蛋白促进神经元铁死亡参与蛛网膜下腔出血后早期脑损伤[J].第三军医大学学报ꎬ2019ꎬ41(15):1407 ̄1414.DOI:10.16016/j.1000 ̄5404.201902124.[34]㊀ChenXꎬXuSꎬZhaoCꎬetal.RoleofTLR4/NADPHoxidase4path ̄wayinpromotingcelldeaththroughautophagyandferroptosisduringheartfailure[J].BiochemBiophysResCommunꎬ2019ꎬ516(1):37 ̄43.DOI:10.1016/j.bbrc.2019.06.015.[35]㊀BabaYꎬHigaJKꎬShimadaBKꎬetal.ProtectiveEffectsoftheMecha ̄nisticTargetofRapamycinagainstExcessIronandFerroptosisinCardiomyocytes[J].AJPHeartandCirculatoryPhysiologyꎬ2017ꎬ314(3):H659 ̄H668.DOI:10.1152/ajpheart.00452.2017. [36]㊀ZhuHYꎬHuangZXꎬChenGQꎬetal.Typhaneosidepreventsacutemyeloidleukemia(AML)throughsuppressingproliferationandin ̄ducingferroptosisassociatedwithautophagy[J].BiochemBiophysResCommunꎬ2019ꎬ516(4):1265 ̄1271.DOI:10.1016/j.bbrc.2019.06.070.[37]㊀MaSꎬDielschneiderRFꎬHensonESꎬetal.Ferroptosisandautophagyinducedcelldeathoccurindependentlyaftersiramesineandlapatinibtreatmentinbreastcancercells[J].PLoSOneꎬ2017ꎬ12(8):e0182921.DOI:10.1371/journal.pone.0182921.(收稿日期:2019-05-27)。
临床病理学考试题及答案

临床病理学习题部分一、最佳选择题1.下列哪点不是疵状癌的特点()A.较为少见B.为腺癌的亚型C.癌细胞分化较成熟D.生长慢E.很少转移2.以下哪一种不是炎性假瘤的特点()A.炎症导致的局部组织增生B.为癌前病变C.表现为肿瘤样团块D.大体和影像学不能与肿瘤区别E.好发于肺和眼眶3.痰涂片阳性率最高的肺癌类型是()A.鳞状细胞癌B.腺癌C.小细胞癌D.大细胞癌E.细支气管肺泡癌4.颅内最常见的原发性肿瘤是()A.脑膜瘤B.胶质瘤C.垂体腺瘤D.神经鞘瘤E.髓母细胞瘤5.关于髓母细胞瘤的叙述哪点是错误的()A.多见于小儿B.常位于小脑C.属原始神经上皮肿瘤D.肿瘤细胞可形成菊形团E.恶性程度低,预后很好6.下列骨肿瘤中属于高度恶性的是()A.尤文(Ewing)瘤B.骨母细胞瘤C.软骨母细胞瘤D.皮质旁骨肉瘤E.巨细胞瘤7.前列腺癌患者血清中增高的物质是()A.酸性磷酸酶B.碱性磷酸酶C.癌胚抗原D. CA19一9E .AFP8.关于肇丸肿瘤下述哪点是错误的()A. 90%以上为生殖细胞来源B.以精原细胞瘤最常见C.胚胎癌为高度恶性D.绒毛膜癌与妊娠有关,不会发生在翠丸E.非生殖细胞肿瘤以淋巴瘤多见9.以下关于甲状腺髓样癌的叙述是错误的()A.较乳头状腺癌少见B.恶性程度较乳头状腺癌高C.患者多为中老年人D.肿瘤可分泌降钙素E.起源于滤泡上皮细胞10.牙眼瘤在性质上属于()A.恶性肿瘤B.交界性肿瘤C.良性肿瘤D.癌前病变E.炎性修复11.隆突性纤维肉瘤的特点不包括()A.好发于真皮B.肿瘤细胞呈梭形,轮辐状C. WHO软组织肿瘤分类中属‚中间型‛D.复发率较高E.远处转移多见12.软组织肉瘤的特点不包括()A.分布广泛、类型多B.发病率和癌大致相等C.肿瘤形态相似者多,鉴别有时困难〃D.恶性肿瘤的组织起源较难确定E.区别良恶性较癌困难13.间变性大细胞淋巴瘤特征性的阳性免疫标记是()A. CD20B. CD30C. CD68D. CD56E. CD9914.下列哪点不是霍奇金淋巴瘤的特征()A.病变从一个或一组淋巴结开始,逐步扩散B.原发于结外常见C.瘤组织成分多样D.含有独特的R一S细胞E.占全部淋巴瘤的10%一20%15.对滤泡型淋巴瘤的描述哪点要除外()A.恶性程度通常较低B.肿瘤细胞形成滤泡状结构C.来源于T细胞D.常有BCL一2蛋白高表达E.我国的发病率较西方国家低16.Burkitt淋巴瘤来源于下列何种细胞()A. T淋巴细胞B. B淋巴细胞C. R一S细胞D. NK细胞E.单核细胞17.多发性骨髓瘤是由下列何种细胞恶变而来()A. T淋巴细胞B.单核细胞C.嗜酸性粒细胞D.嗜碱性粒细胞E.以上都不是18.下列肿瘤中血液中可出现M蛋白的是()A.霍奇金淋巴瘤B.恶性组织细胞增生症C.组织细胞增生症XD.毛细胞白血病E.多发性骨髓瘤19.非霍奇金淋巴瘤最少见的组织类型是()A.滤泡型淋巴瘤B.粘膜相关淋巴瘤C.间变性大细胞淋巴瘤D.组织细胞性淋巴瘤E.外周T细胞淋巴瘤20.中枢神经系统白血病最常见的是()A.急性粒细胞性B.急性淋巴细胞性C.急性单核细胞性D.慢性粒细胞性E.慢性淋巴细胞性21.成人肾恶性肿瘤最常见的是()A.移行细胞癌B.肾细胞癌C.肾母细胞瘤D.鳞状细胞癌E.血管肉瘤22.肾细胞癌侵犯的特点是()A.常侵人肾静脉B.常侵人肾动脉C.常侵人输尿管D.常侵犯肾上腺E.常侵犯肾周淋巴结23.成人的星形细胞瘤最好发的部位是()A.额叶B.顶叶C.颖叶D.枕叶E.小脑24.下列肿瘤中不属于神经内分泌肿瘤的是()A.化学感受器瘤B.甲状腺髓样癌C.嗜铬细胞瘤D.肝细胞癌E.胰岛细胞瘤25.下列肿瘤属于神经内分泌肿瘤的是()A.甲状腺乳头状腺癌B.卵巢颗粒细胞癌C.肾上腺皮质腺瘤D.皮肤Merkel细胞癌E.胰腺腺癌26.脑膜瘤最好发的部位是()A.后颅凹B.小脑桥脑角C.前额部D.蝶骨脊侧面E.矢状窦旁大脑镰两侧27.关于视网膜母细胞瘤的描述哪点是错误的()A. 6岁以上儿童常见B.多有抑癌基因Rb的突变C.可见于单侧,也可双侧眼内D.恶性程度高,预后较差 E.累及视神经预后更差28.绒毛膜上皮癌常可产生()A. AFPB. CEAC. HCGD. CA一125E. PSA29.鉴别前列腺癌转移最有意义的标记是()A.前列腺特异性抗原B.特异性酸性磷酸酶C.表皮生长因子D. p53蛋白E.癌胚抗原30.胰腺癌最常发生的部位是()A.胰头B.胰体C.胰尾D.整个胰腺E.各部位均匀分布31.下列哪一项不符合肺小细胞癌?()A.多发生于肺周边部B.细胞小、排列密集C.恶性度高D.肿瘤细胞内可见神经内分泌颗粒E.起源于嗜银细胞32.不符合鼻息肉的特点的是()A.良性肿瘤B.肉眼呈灰白色C.间质水肿D.腺体增生,并可呈囊状扩张E.间质炎性细胞侵润33.鼻腔内翻性乳头状瘤的特点的是()A.慢性炎症性增生B.不易恶变C.鳞状上皮团呈柞状向间质内生长D.与HPV感染无关 E.术后不复发34.关于结节性甲状腺肿的记述,哪一项是错误的?()A.结节具有完整的包膜B.结节对周围甲状腺组织无明显压迫作用C.可见纤维组织增生D.结节内常有出血E.年轻女性多发35.不符合慢性淋巴细胞性甲状腺炎的描述是()A.又称桥本甲状腺炎B.为自身免疫性疾病C.引起甲状腺肿大D.大量淋巴细胞浸润E.滤泡上皮增生36.不属于APUD瘤的是()A.肺小细胞癌B.胃肠道类癌C.胃泌素瘤D.胰岛细胞瘤E.神经纤维瘤37.不符合嗜铬细胞瘤的描述是()A.瘤细胞呈大多角形,可见瘤巨细胞,有一定多形性B.排列为条索状、片状C.间质为血窦D.可有包膜E.异型性显著为恶性指标38.癌转移至淋巴结时,首先出现在()A.髓窦B.边缘窦C.淋巴滤泡内D.副皮质区E.淋巴结门部39.癌前病变不包括()A.大肠腺瘤B.甲状腺腺瘤C.膀胧乳头状瘤D.阴茎乳头状瘤E.家族性腺瘤性息肉病40.不符合皮肤基底细胞癌的描述是()A.好发于面部B.多见于老年人C.易发生转移D.与紫外线照射有关E.可形成溃疡41.不符合毛细血管瘤特征的是()A.儿童常见B.多为先天性发生C.多见于皮肤D.成年后可停止发展,甚至可以自行消退E.膨胀性生长42.不符合骨瘤特点的是()A.良性肿瘤B.好发于头面骨及领骨C.由成熟的骨质组成D.骨母细胞异型性明显E.可引起压迫症状43.有关纤维瘤病的描述,错误的是()A.是一种恶性肿瘤B.呈浸润性生长C.可复发‘D.不转移E.肿瘤无包膜44.平滑肌肉瘤的诊断依据中,最重要的是.()A.肿瘤细胞丰富B.肿瘤粘液变性C.核分裂像多D.细胞的多形性明显E.肿瘤出血坏死45.男性,30岁,右胫骨上端病理性骨折,局部被囊性肿物破坏。
BMI_1基因与肿瘤关系研究进展

综述・讲座¬BMI21基因与肿瘤关系研究进展胡中华, 胡义德第三军医大学新桥医院全军肿瘤中心,重庆400037Study condition of the relationship between BMI21gene and tumorHU Zhong2hua,HU Yi2deDepartment of Oncology,Xinqiao Hos pital of Thi rd Military Medical University,Chongqing400037,P.R.China【摘要】 BMI21基因为一种致癌基因,属于Pc G家族成员,通过抑制IN K4a2ARF 位点,从而与多种实体瘤的发生发展密切相关;BMI21基因在维持正常干细胞自我更新及多向分化方面起重要作用;具有促进白血病干细胞增殖的能力。
肿瘤干细胞是形成不同分化程度肿瘤细胞和肿瘤不断扩展的源泉,靶向杀死肿瘤干细胞应成为肿瘤治疗的根本目的。
中华肿瘤防治杂志,2006,13(23):1824-1827[ABSTRACT] BMI21,a member of the Polycomb Group(Pc2G) family genes,is a human proto2oncogene.Through inhibiting IN K4a2 ARF,BMI21gene has a deep relationship with many solid tumors. BMI21gene play an important role in maintaining self2renewal and multipolar differentiation of normal stem cells,as well as in promo2 ting proliferation of the leukemic stem cells.Tumor stem cells are the source of forming diverse tumor cells and cotinuously spread of tumors,so aimed killing of tumor stem cells should become ultimate purpose of the therapy of tumors.Chin J Cancer Prev T reat,2006,13(23):1824-1827【关键词】 核蛋白质类;原癌基因蛋白质类;肿瘤;综述文献[KE YWOR DS] nuclear proteins;proto2oncogene proteins;neoplasms;review literature【中图分类号】 R730 【文献标识码】 A 【文章编号】 1673-5269(2006)23-1824-04 BM I21(B2cell2specific moloney leukemia virus insert site1)基因为一种属于Pc G(Polycomegroup)家族成员的致癌基因,自1991年在鼠淋巴瘤中被发现后即引起生物医学领域的高度关注。
条件重编程细胞在肿瘤治疗中的研究进展

条件重编程细胞在肿瘤治疗中的研究进展张卉卉【摘要】癌症是由于致癌因子导致的原癌基因和抑癌基因突变引发的一系列异质性疾病的统称.尽管近年来高通量测序技术与靶向治疗对癌症治疗取得了突破性的进展,但临床转化研究的高失败率使得抗肿瘤药物的创新进展有限.肿瘤细胞系培养技术的出现有力推动了肿瘤生物学研究的发展,但在肿瘤新疗法的临床转化研究中,肿瘤细胞系的预测能力有限且不稳定.条件重编程细胞(conditional reprogrammed cells,CRCs)为从患者组织建立衍生的正常和肿瘤上皮细胞培养物,能准确地反映原始肿瘤细胞异质性及遗传信息多样性,为临床前药效个性化筛选评估以及临床靶向治疗后耐药的新型药物探索提供了新的研究资源.本文对CRC及其在肿瘤新疗法中的应用、挑战性及局限性以及在肿瘤精准医学中的研究进展进行综述.【期刊名称】《中国肿瘤临床》【年(卷),期】2018(045)015【总页数】4页(P808-811)【关键词】条件重编程细胞;癌症;个性化治疗;精准医学【作者】张卉卉【作者单位】宁波大学医学院附属鄞州医院呼吸内科浙江省宁波市315000【正文语种】中文据国家癌症中心数据统计,中国恶性肿瘤发病率为380.4万/年,癌症已成为中国乃至全球主要的死亡原因,给社会带来了沉重的经济负担。
癌症是一种高度异质性的疾病。
近年来高通量测序技术不仅阐明了多种癌症的突变过程,而且提供了一个全面的癌症基因目录[1],目前基因组分析和靶向治疗已经应用到几乎所有类型的肿瘤治疗,但随之出现肿瘤耐药性、耐药的原因以及新型肿瘤药物的研发已经成为临床重点关注的问题。
迄今为止,传统建立的肿瘤细胞系一直是细胞、分子和癌症生物学的支柱,然而肿瘤细胞系经长期体外培养后,其生物学行为及基因谱表达水平、肿瘤异质性都与原始肿瘤组织存在较大的差异,不能准确地反映原始肿瘤异质性和肿瘤细胞微环境等特点,因此不适用于筛选临床新型化疗药物,以及对肿瘤患者进行个体化治疗。
端粒酶与细胞永生化
Te o e a e a d e l a m m o t lz to lm r s n c l ul r i raia in
W ANG a — Gu H n o, ZHA NG i Xn
( ole tm tl i,T eF ut layMe i l nvr t X ’ n7 0 3 hn ) C l g o o aoo y h o r Mitr dc i sy, i a 1 0 2 C ia g fS g h i a U ei
[ b ta t A s c] r Etbi me t fi mo ai dcl l ei oeo h o a ddfc l sbet i elbooy s l h n m r l e el i s n fteh t n iiut u jcs nc l i g . a s o t z n l
T l me e tc r mo o n s i u a y t s sa l h h o s me T l me a e,a e o r s a h o s me e d n e k r oe tb e t e c r mo o . e o r s n RNA — p o e n c mp e r t i o lx,a d ds mu t l e o r e e t o i 一p me e d b s n l p e t l me i r p a st s3 i c t i r n y u i g RNA c mp n n s tmp a e o o e ta e lt .T u c o i x r s i n o e t — h s e t p c e p e so ft e h
在大多数多细胞 真核生物 , 着 体细胞 与生殖 随 细胞 的分离 , 细胞 分 化 赋 予 了体 细 胞 有 限复 制 及 生 殖 细 胞 无 限 增 殖 的 能 力 。在 正 常 体 细 胞 向永 生 化 细 胞 (mmotle el) 肿 瘤 细 胞 的 转 化 过 程 中 i r i dcl 和 az s 可 能 也 存 在 着 与 生 殖 细 胞 类 似 的 机 制 。 端 粒 (e t—
人永生化胰腺星形细胞系的构建及其功能的验证
人永生化胰腺星形细胞系的构建及其功能的验证韩世纪1,2,练国达1,2,陈少杰1,2,黄开红1,2*,李佳佳1,3*[摘要]目的构建人永生化胰腺星形细胞(PSCs)系并探讨其生物学功能。
方法采用outgrowth方法提取人原代胰腺星形细胞(hPSCs),慢病毒感染方法导入SV40LT基因和hTERT基因,嘌呤霉素筛选稳定高表达两个基因的细胞(im PSCs),qRT⁃PCR和Western bolt方法检测表达水平;CCK8法、核型分析、免疫荧光、体内成瘤实验检测im PSCs生长曲线,染色体数目,胶原基质蛋白表达和有无恶性表型转化;共培养方法、CCK8法、细胞划痕、迁移侵袭方法检测imPSCs促瘤作用。
结果hPSCs呈多角形和梭形,慢病毒感染的im PSCs,显微镜下有亮绿色荧光;im PSCs细胞高表达SV40LT基因和hTERT基因;im PSCs生长曲线呈典型的“S”型,染色体数目增多,体内不能成瘤,表达α⁃SMA、Collagen I、Fibronectin,具有PSCs的生物学功能和特点;其与胰腺癌细胞共培养后,明显促进肿瘤生长,与人原代PSCs功能相当。
结论采用慢病毒转染SV40LT基因和hTERT基因可成功构建人胰腺星形细胞永生化细胞系,为胰腺星形细胞本身及胰腺癌微环境相关研究提供实验材料和模型。
[关键词]胰腺癌;胰腺星形细胞;永生化细胞系doi:10.3969/j.issn.1009⁃976X.2021.01.007中图分类号:R735.9文献标识码:AConstruction and functional verification of immortalized human pancreatic stellate cellsHAN Shi⁃ji1,2,LIAN Guo⁃da1,2,CHEN Shao⁃jie1,2,HUANG Kai⁃hong1,2*,LI Jia⁃jia1,3*1.Key Laboratory of Epidemiology and Gene Regulation of Malignant Tumors,Sun Yat⁃sen MemorialHospital,Sun Yat⁃sen University,Guangzhou510120,China;2.Department of Digestive System Depart⁃ment,Sun Yat⁃sen Memorial Hospital,Sun Yat⁃sen University,Guangzhou510120,China;3.Depart⁃ment of Nephropathy Department,Sun Yat⁃sen Memorial Hospital,Sun Yat⁃sen University,Guangzhou510120,ChinaCorresponding author:HUANG Kai⁃hong,*****************;LI Jia⁃jia,****************[Abstract]Objective To construct the human pancreatic stellate cell line immortalization anddiscuss their biological function.Methods We extracted human primary pancreatic stellate cells(hPSCs)by outgrowth and then infected the cells with SV40LT and hTERT genes using lentivirus infection.Thecells(named imPSCs)with stable and high expression of SV40LT and hTERT genes were screened with puromycin.The next,qRT⁃PCR and Western Bolt methods were used to detect the expression levelsof the two K8method,karyotype analysis and immunofluorescence method were conducted respectively to detect the growth curve,chromosome number,protein expression ofα⁃SMA,collagen Iand fibronectin of the imPSCs.We also performed tumor formation assay in vivo to detect whether theimPSCs had malignant phenotypic transformation.The tumorigenesis of imPSCs was detected by transwellcell co⁃culture,CCK8methods,wound healing assay,migration and invasion methods.Results Humanprimary PSCs were polygonal and spindle shaped,and bright green fluorescence was observed under the microscope after successfully infected lentivirus.In imPSCs,SV40LT and hTERT genes were stably andhighly expressed in mRNA and protein levels.The imPSCs had typed“S”growth curve and increased基金项目:国家自然科学基金青年科学基金项目(82003175);广东省医学科学技术研究基金项目(A2019447)作者单位:1.中山大学孙逸仙纪念医院,广东省恶性肿瘤表观遗传学与基因调控重点实验室,广州510120;2.中山大学孙逸仙纪念医院消化内科,广州510120;3.中山大学孙逸仙纪念医院肾内科,广州,510120*通讯作者:黄开红,Email:*****************;李佳佳,Email:****************胰腺癌恶性程度高,肿瘤间质纤维化是其重要病理特征,纤维化程度与不良预后相关[1]。
NF-κB通路激活在肿瘤发生发展中的作用
NF-κB通路激活在肿瘤发生发展中的作用郑宏伟,袁劲松(综述);高燕宁,程书钧(审校)(中国医学科学院肿瘤研究所病因及癌变研究室,北京100021)【摘要】 NF-κB是一组重要的转录因子,参与细胞中多种生理过程,具有复杂的调节机制。
近年来的研究发现NF-κB在在肿瘤发生发展过程中具有重要而且复杂的作用,总体来说,NF-κB通路的异常激活可导致一系列与肿瘤相关基因的异常表达,从而抑制肿瘤细胞凋亡,促进正常细胞转化及肿瘤血管形成和转移等,直接影响恶性肿瘤的发生和发展,其抑制剂的开发亦为肿瘤治疗提供了一种新的选择。
【关键词】 NF-κB; 肿瘤; 凋亡; 转化; 转移中图分类号: R730.231 R734.2 文献标识码: A 文章编号: 1004-616X(2006)05-0407-04NF-κB是一组具有特殊DNA结合序列的转录因子,主要调节炎症和自身免疫反应。
最近在基因工程小鼠中进行的研究表明,NF-κB对某些上皮来源器官的精确发育也具有重要作用[1]。
近些年很多研究发现NF-κB能够控制细胞增殖和癌变,调控细胞周期及凋亡,影响细胞分化,促进肿瘤转移。
在多数白血病和实体肿瘤中,NF-κB或IκB(Inhibitor of kappaB)家族成员往往通过遗传学改变或信号转导异常,而持续性激活或出现异常的核定位[2]。
NF-κB-Rel家族包括5个成员,即RelA(p65)、RelB、c-Rel、p105-p50(NF-κB1)和p100-p52(NF-κB2),在所有细胞中都有表达。
它们的N-末端均包含一个约300个氨基酸的高度同源序列,称为Rel同源结构域(RHD),介导其与DNA结合及二聚化。
p105和p100通过蛋白酶解加工产生具有活性的DNA结合形式——p50 和 p52。
在未受到刺激的正常细胞中,NF-κB亚基与I-κBα、β或γ结合,以非活性形式被阻滞在胞浆中。
p105 和 p100的未加工形式的C-末端含有锚蛋白重复序列,此序列也能和I-κBs一样将p105 和 p100的二聚体阻滞在胞浆中[3,4]。
肿瘤细胞的培养
20
1、取材
人肿瘤细胞来自外科手术或活检瘤组织。取材 部位非常重要,体积较大的肿瘤组织中有退变 或坏死区,取材时尽量避免,要挑选活力较好 的部位。 癌性转移淋巴结或胸腹水是很好的培养材料。
21
取材后尽快将癌组织进行培养,一般4小时内 细胞存活最好。标本在4℃存放,不宜超过24 小时。
22
人癌细胞建系必须要有完整的记录,包括组织 来源、病人姓名、住院号、年龄、性别、临床 诊断、病理诊断(应明确分类分期)、术前放 化疗情况,为细胞建系的重要材料。 为了鉴定建立的细胞系来源和生物学特性,最 好留有病人正常组织和血标本。
18
② 肿瘤细胞的自分泌也会因分散培养而被稀释,达不到 肿瘤生长的需求,降低肿瘤细胞的生长增殖力。 ③ 并非所有肿瘤细胞都有强的生长活力和长的生命周 期,只有干细胞才有强的增殖生长能力,但这些细胞 数量很少。 ④ 离体培养肿瘤细胞可能需求与体内相似的特殊生存条 件。
19
(一)癌细胞原代培养
癌细胞建系的方法和程序相似于正常细胞,包括 标本收集和处理、原代培养、传代、换液、冻 存、复苏等过程。 培养成功关键在于:取材、成纤维细胞的排除、 选用适宜的培养液和培养底物等几个方面。
34
⑤ 其它方法
聚丙烯酰胺抑制成纤维细胞生长 密度梯度离心等
35
(2)选择性培养基
选择性培养基是针对特殊细胞生长的营养要求 设计的。 在癌细胞建系中,选择性培养基的应用是提高 癌细胞体外存活、抑制成纤维细胞生长的关键 技术改进。
HITES选择培养基是一种改良RPMI-1640培养基,含有氢化可
的松、胰岛素、转铁蛋白、雌激素、硒等成分。HITES培养基适 合用于人小细胞肺癌建系。
具体做法是:
1)收集培养的肿瘤细胞,用无血清培养液洗一遍;2)计数 后将0.2ml含2×106~2×107个细胞的细胞悬液注射到动物皮下。 约一个月左右(最短在3天后),可见注射局部有肿瘤出现,甚至 有转移瘤形成。经病理组织学检查可以确定肿瘤是否由接种的细 胞产生。也可接种到腹腔或通过鼠尾静脉注射。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
人肺癌相关成纤维细胞永生化
人肺癌相关成纤维细胞的永生化是指在体外培养条件下,肺癌组织来源的成纤维细胞经过一系列基因改造或处理,使其具有无限增殖能力和长寿命的特性。
在正常的细胞生长中,细胞有一定的增殖次数,当达到执行信号时,细胞会进入细胞凋亡或细胞老化的过程。
然而,肺癌细胞常常失去了自我调控的能力,使得其能够无限增殖并逃脱凋亡的过程。
这种现象被称为细胞永生化。
为了实现肺癌相关成纤维细胞的永生化,研究人员通常采用以下方法:
1. 过表达促进细胞增殖的基因:研究人员可能会将促进细胞增殖的基因如TERT等转染至成纤维细胞中,来增加其增殖的能力。
2. 抑制细胞凋亡相关的基因:研究人员可能会通过抑制细胞凋亡相关的基因,如p53、BAX等,来阻止细胞凋亡的发生,从而延长细胞寿命。
3. 体外培养条件的优化:研究人员可能会优化细胞培养基的配方,例如添加适当的生长因子和细胞因子,以提供细胞增殖所需的营养和信号。
使用这些方法能够永生化肺癌相关的成纤维细胞,有助于深入研究肺癌的发生机制、细胞增殖和凋亡的调控机制,以及新的
治疗策略的开发。
然而,需要注意的是,这些方法在实际应用中存在伦理和安全性的问题,需要严格的监管和控制。