基于FIASCO技术的合浦珠母贝微卫星标记分离与筛选研究_曲妮妮
马氏珠母贝Pm-miR-183序列分析及功能研究

马氏珠母贝Pm-miR-183序列分析及功能研究熊新威;郑哲;焦钰;邓岳文;杜晓东【摘要】用马氏珠母贝基因组序列,通过生物信息学方法获得Pm-miR-183的前体,长度76 bp,具有典型的茎环结构.成熟序列及前体序列的同源性分析均显示较高的种间保守性.Pm-miR-183前体序列(Pm-pre-miR-183)的系统进化树分析,Pm-pre-miR-183与帽贝同源性较高.靶向关系分析,Pm-miR-183与矿化基因及免疫相关基因均存在靶向作用位点.实时荧光定量分析,Pm-miR-183在马氏珠母贝的各组织中均有表达.注射 Pm-miR-183 mimics进行过表达,Pm-miR-183的表达量在套膜区(MP)中显著上调(P<0.05).扫描电子显微镜(scanning electron microscope,SEM)检测,Pm-miR-183过表达会导致贝壳珍珠层生长出现紊乱.%MiRNAs are non-coding small RNAs involved in negatively transcriptional regulation. In our previous study, we had identified a miR-183 homologue in Pinctada fucata martensii by Solexa deep sequencing.Based on bioinformatics results obtained from whole genome sequence of P. f.martensii, the precursor Pm-miR-183(Pm-pre-miR-183)obtained has a length of 76 bp and a typical stem-loop structure. Homology analysis of mature sequence and previous sequence indicated that they were highly conserved with other species. Phylogenetic analysis of precursor sequence showed that Pm-pre-miR-183 had a higher homology to Lottia gigante. The potential interactions among Pm-miR-183 and several mineralized genes and immune-related genes were obtained by target prediction. Pm-miR-183 was expressed in different tissues. In addition, after injection of Pm-miR-183 mimics, the expression of Pm-miR-183 was significantly up-regulated in the mantle pallial. The result of SEM showed that there were adnormal growth of nacre in shell. This study shows that Pm-miR-183 could be involved in the formation of pearl oyster in P.f.martensii.【期刊名称】《广东海洋大学学报》【年(卷),期】2017(037)006【总页数】8页(P11-18)【关键词】马氏珠母贝;Pm-miR-183;生物矿化;miRNA调控【作者】熊新威;郑哲;焦钰;邓岳文;杜晓东【作者单位】广东海洋大学水产学院//广东省珍珠养殖与加工工程技术研究中心,广东湛江 524088;广东海洋大学水产学院//广东省珍珠养殖与加工工程技术研究中心,广东湛江 524088;广东海洋大学水产学院//广东省珍珠养殖与加工工程技术研究中心,广东湛江 524088;广东海洋大学水产学院//广东省珍珠养殖与加工工程技术研究中心,广东湛江 524088;广东海洋大学水产学院//广东省珍珠养殖与加工工程技术研究中心,广东湛江 524088【正文语种】中文【中图分类】S917.4MicroRNA(miRNA)是在动植物以及病毒体内发现的一种参与转录后调控的非编码小RNA[1]。
2018年度市科委首都临床特色应用研究专项资助名单

12
2018.06-2021.05
PET/CT分子显像评价冠心病心肌存活性和脑神经细胞葡萄糖代谢水平的相关性研究
首都医科大学附属北京安贞医院
张晓丽
16
2018.06-2021.05
导管消融治疗肥厚型心肌病心房颤动的临床研究
首都医科大学附属北京安贞医院
汤日波
16
2018.06-2021.05
八段锦运动处方对急性心肌梗死介入术后患者早期康复治疗的临床研究
神经根袖加固重建及骶管成形术治疗骶管囊肿随机对照研究
北京大学第三医院
刘彬
16
2018.06-2021.05
儿童紫癜性肾炎早期多维度预后评估新方法研究
首都儿科研究所附属儿童医院
陈朝英
16
2018.06-2021.05
应用3D打印微孔钛合金假体治疗四肢长骨大段骨缺损的临床研究
北京大学第三医院
田耘
100
2018.06-2021.05
中国人民解放军第三〇九医院
龚文平
5
2018.06-2021.05
困难脱机患者高流量湿化氧疗脱机方案与T管脱机的前瞻性队列研究
中国人民解放军第三〇九医院
张玉想
10
2018.06-2021.05
CT在呼吸衰竭儿童肺复张中新技术的建立与研究
首都儿科研究所附属儿童医院
闫淯淳
16
2018.06-2021.05
中国中医科学院广安门医院
李深
16
2018.06-2021.08
荧光胸腔镜在手汗症交感神经手术中的应用研究
北京市海淀医院
黄宇清
16
2018.06-2021.08
基于数据分析的六自由度机器人骨关节矫形与肢体重建临床优化研究
国家自然科学基金委员会生命科学部2016年度青年基金项目_0a55ef92_a4

第28卷第12期2016年12月V ol. 28, No. 12Dec., 2016生命科学Chinese Bulletin of Life Sciences文章编号:1004-0374(2016)12-1493-54国家自然科学基金委员会生命科学部2016年度青年基金项目DOI: 10.13376/j.cbls/20161861微生物学细菌基因组歧化和遗传界限的产生:新物种形成的分子基础唐 乐哈尔滨医科大学我国沿海盐田嗜盐古菌胞外蛋白酶多样性研究侯 靖江苏大学反硝化细菌的鉴定及其与胆固醇降解相关蛋白、基因的初探丁 滨浙江中医药大学大连新港石油污染海域沉积物中厌氧微生物种群和功能基因多样性与氮源响应规律研究陈 超大连民族大学苏云金芽胞杆菌晶胞粘连表型菌株资源及形成机制的多样性研究王月莹华中农业大学粪产碱菌杀线虫活性物质挖掘及作用线虫方式研究鞠守勇华中农业大学冰川稀有低温细菌Cryobacterium物种多样性、分类学及冷适应性研究刘 庆中国科学院微生物研究所捕食青枯菌的粘细菌资源分离与功能评价李安章广东省微生物研究所环境因子对花生根瘤菌遗传多样性和分布的影响机制研究李 岩中国科学院烟台海岸带研究所家蚕病原细菌多样性、分布规律及其与蚕体共生菌关系研究周洪英湖北省农业科学院杏褪绿卷叶植原体宿主植物内生细菌的群落变化及与宿主抗/感病性的相关性研究韩 剑新疆农业大学竹虫肠道微生物群落结构及纤维素酶基因多样性分析王彦伟农业部沼气科学研究所木质纤维素水解残渣厌氧消化过程中微生物群落的组成与功能分析汤晓玉农业部沼气科学研究所海绵复杂共生体中放线菌新分类单元及新活性物质发现李 蕾上海交通大学贵州喀斯特洞穴放线菌多样性及生物活性菌株筛选房保柱中山大学放线菌中五角多酚类化合物的基因组挖掘刘力伟中国科学院微生物研究所疣孢菌CRISPR/Cas9基因组编辑技术的建立和评价谢 峰中国科学院微生物研究所3种黄连属濒危植物内生放线菌多样性及其抗菌消炎活性初探田守征云南中医学院中国丽赤壳属Calonectria种类及系统发育研究张云霞仲恺农业工程学院东北地区红菇真菌子实体及其地下菌根分子生态学研究冀瑞卿吉林农业大学山东苹果主产区红富士果面酵母菌多样性及对苹果炭疽病的生防潜能陈 汝山东省农业科学院中国毛霉属系统发育研究及DNA条形码筛选王亚宁中国科学院微生物研究所壳二胞属物种分类研究陈 倩中国科学院微生物研究所食药用菌真菌病害病原鉴定及系统分类研究孙敬祖中国科学院微生物研究所七种鞘翅目昆虫共生蛇口壳目真菌资源的分类与分子系统发育研究殷明亮广东省微生物研究所木兰科植物的丛枝菌根真菌多样性及协同进化特征杨安娜安徽师范大学我国海洋性冰川低温真菌多样性研究王曼曼河北大学云芝新颖苔色酸木糖苷衍生物的糖基化机制研究朱丽萍青岛农业大学中国芒果炭疽菌多样性及致病力变异分子机制的研究李其利广西壮族自治区农业科学院赤水河流域枯枝暗色丝孢菌Dematiaceous hyphomycetes多样性研究李小霞遵义师范学院鹅膏科系统发育框架构建及营养方式演化研究蔡 箐中国科学院昆明植物研究所西南地区葡萄座腔菌科四个重要属的分类和分子系统学发育研究刘建魁贵州省农业科学院严紧反应下乙酰化修饰对大肠埃希菌DnaA降解调控的研究张秋芬上海交通大学蓝细菌能量-还原力代谢重平衡新策略及其生理影响研究栾国栋中国科学院青岛生物能源与过程研究所海洋细菌胞外多糖 EPS273 抑制铜绿假单胞菌生物膜形成的分子机理研究吴仕梅中国科学院青岛生物能源与过程研究所1494生命科学第28卷里氏木霉sorbicillinoid类次级代谢产物生物合成机制的研究齐飞飞中国科学院青岛生物能源与过程研究所枯草芽胞杆菌产生羊毛硫细菌素subtilomycin促进其在植物体内定殖的机制邓 运华中农业大学沙雷氏菌属新种YD25中环脂肽类化合物与灵菌红素生物合成的共调控机制研究苏 春陕西师范大学青枯菌生物素合成途径中一种新型甲酯庚二酸单酰ACP酯酶的鉴定及其生物学功能研究贾 佳南京医科大学硫转移蛋白在嗜酸热硫氧化古菌Metallosphaera cuprina硫传递网络中的功能研究刘丽君西安医学院希瓦氏菌中D型β-内酰胺酶诱导表达及其耐药机制的研究音建华南昌大学酿酒酵母生物合成中长链二元酸的动态调控及其作用机制韩 丽郑州轻工业学院土壤杆菌胞外水溶性β-1,3-葡聚糖的生物合成机制研究程 瑞南京理工大学基于AFM力谱与SPR技术的启动子强度高效表征策略的研究张晓娟江南大学P450单加氧酶AveE催化阿维菌素呋喃环形成的机制研究马 莉中国科学院青岛生物能源与过程研究所丁香假单胞菌海藻糖合成与抗水分胁迫相关性研究余希岚湖北大学解脂耶氏酵母中蛋白激酶Snf1参与氮饥饿启动赤藓醇合成的调控机制刘晓燕淮阴师范学院转录因子RpoD调控运动发酵单胞菌乙醇耐受性的机制研究谭芙蓉农业部沼气科学研究所酮基合酶MarO催化maremycin中哌嗪二酮的酰胺键形成和成环释放机制黄婷婷上海交通大学多模块持续性内切纤维素酶CcCel9A的持续性驱动力研究张坤迪中国科学院青岛生物能源与过程研究所里氏木霉外切纤维素酶CBH I的水解机理研究和理性设计王业飞中国科学院青岛生物能源与过程研究所单酶催化多步连续反应中底物结合模式研究姚明东天津大学灰盖鬼伞胞外β葡聚糖苷酶BGL2不同变体产生机制和生理功能的研究刘中华南京师范大学蛹虫草隐花色素Cry-DASH功能及作用机制王 芬中国科学院微生物研究所酶分子对黄姜细胞壁降解及皂苷释放的影响机理研究魏 蜜湖北工程学院基因内置特征影响密码子偏好对重组蛋白表达调控的研究周 勉华东理工大学大肠杆菌脂多糖转运关键蛋白LptFG的功能研究向泉桔四川农业大学金黄色葡萄球菌七异戊二烯二磷酸合成酶SaHepPPs晶体结构和抗菌药物开发李 倩中国科学院天津工业生物技术研究所以产油微藻海洋微拟球藻为模式的二酰甘油酰基转移酶功能机制研究辛 一中国科学院青岛生物能源与过程研究所聚酮合酶pks7和pks11对海洋草酸青霉合成Oxalicumone A及其环境适应性的调控机制研究王 洁中国科学院南海海洋研究所大肠杆菌中galE转录暂停调控下游ρ依赖型转录终止的分子机制王 璕华中农业大学XsfP催化的膜内受控蛋白水解在黄单胞菌致病过程中的调控功能邓超颖中国科学院微生物研究所鞘脂合成调控因子Orm1在球孢白僵菌中的功能鉴定及其致病机理研究王娟娟济南大学糖多孢红霉菌中与红霉素合成相关调控因子间的干扰机制研究汪焰胜安徽大学特异腐质霉转录因子HiProA调控纤维素酶表达机制的研究徐欣欣中国农业科学院生物技术研究所利用代谢拨动开关调控大肠杆菌芳香型氨基酸的合成古鹏飞济南大学CRISPR/Cas9介导的基因组进化构建固态发酵耐热酵母及机理研究李鹏松清华大学代谢工程改造枯草芽孢杆菌合成N-乙酰神经氨酸关键问题的研究刘延峰江南大学米曲霉Sedolisin家族基因相关联的外源蛋白表达分泌机制的研究朱 琳江苏大学L-异亮氨酸合成途径基因在谷氨酸棒杆菌中的模块化协调表达和代谢调控机制研究尹良鸿浙江农林大学酿酒酵母合成灵芝酸类似物的研究肖 晗上海交通大学聚3-羟基丁酸-4-羟基丁酸酯的全新合成代谢通路的构建与优化尹 进清华大学二硫吡咯酮生物合成途径中N4甲基转移酶的表征及组合生物合成应用黄 胜湖北民族学院大肠杆菌中植物CPR:P450模块化自主共价连接系统的构建及其在咖啡酸合成中的应用李宜奎江苏省中国科学院植物研究所胆囊癌微环境菌群结构特征和功能学的初步研究吴文广上海交通大学国家自然科学基金委员会生命科学部2016年度资助项目第12期1495单细胞水平高产虾青素雨生红球藻高通量筛选新方法研究王喜先中国科学院青岛生物能源与过程研究所基于功能活性可视化追踪的芳烃降解微生物高通量筛选方法方 云广东省微生物研究所微生物漆酶数据挖掘及底物杂泛性分析张寅良安徽大学亚硝酸盐还原酶转录调控蛋白NsrR去阻遏机制研究令桢民兰州大学异丙甲草胺脱烷基酶基因的克隆及脱烷基酶特性和功能的研究陈 青枣庄学院面向人工定向重构纤维素菌群的共生分子机理研究杜 然清华大学迪茨氏菌转录调控蛋白AlkX对烷烃降解过程的全局调控研究梁洁良中山大学GDSL酯酶的定向进化与分子改造研究丁俊美云南师范大学整合宏组学方法研究番茄秸秆堆肥生境中的关键微生物及其功能张小梅青岛农业大学假单胞菌CNB-12分解代谢3-氯硝基苯的机理研究闵 军中国科学院烟台海岸带研究所假单胞菌LY1降解3-吲哚乙酸上游途径分子机理研究于 浩青岛农业大学一种二苯醚类污染物直接开环角度双加氧酶的分子机制研究蔡 舒江苏省农业科学院根际促生解淀粉芽孢杆菌SQR9组氨酸激酶KinA-E响应的根系分泌物信号鉴定刘云鹏中国农业科学院农业资源与农业区划研究所膜囊泡介导Geobacter sulfurreducens胞外电子传递过程及机制刘 星福建农林大学短短芽孢杆菌Spo0A蛋白介导的生物膜调控通路中阻遏基因的功能分析侯启会山东农业大学高盐环境下盐单胞菌(Halomonas)降解偶氮染料的机制研究郭 光南京工程学院纳米氧化铝促海洋枯草芽孢杆菌抗菌物质合成机理研究于秀霞山东大学红树林湿地生态系统中MCG古菌mcrA基因的时空变化规律和环境效应研究潘 杰深圳大学基于秀丽隐杆线虫模型的抗菌性海洋益生菌筛选及其机理研究李英秀山东大学滇池水华中控藻菌的杀藻相关基因及其功能研究杨彩云西南大学太湖蓝藻群体颗粒附生细菌的宏基因组学研究张军毅东南大学稻田藻-菌生物膜中胞外聚合物对水体营养水平变化的响应机制刘俊琢中国科学院南京土壤研究所高原藏族人特异性皮肤菌群与紫外辐射适应关系研究曾 博四川农业大学学龄前儿童“口腔菌群年龄”的遗传基础和过程机制滕 飞中国科学院青岛生物能源与过程研究所地衣共生系统中地衣细菌群落结构及其功能研究司红丽山东师范大学广东省凡口铅锌矿矿山酸性废水中古菌的多样性与功能研究陈林兴中山大学鸡肠道微生物代谢黄曲霉毒素B1的分子机制研究汪玲玲华南农业大学西藏热泉环境栖热菌类群多样性及其生理生态功能研究周恩民中山大学中华蟾蜍蝌蚪肠道菌群的结构及其影响因素研究宋晓威信阳师范学院转录激活因子PilR在σ54调控水稻白叶枯病菌毒性中的功能研究余 超中国农业科学院植物保护研究所TLR介导的ATP释放在副猪嗜血杆菌感染上调IL-1β中的作用研究于 江山东省农业科学院分枝杆菌aceE基因影响细胞壁合成代谢的机制研究陈素婷首都医科大学碳青霉烯耐药鲍曼不动杆菌新优势克隆ST208毒力因子鉴定及功能研究陈 燕浙江中医药大学沙门氏菌毒素效应蛋白SifA C-端结构域毒力功能机制的研究赵伟栋新乡医学院MexS调控铜绿假单胞菌III型分泌系统分子机制的研究靳永新南开大学基于炭疽芽孢杆菌S-层蛋白自组装的双抗高灵敏检测纳米材料的研究王旭颖武汉血液中心c-di-AMP信号通路在炭疽杆菌致病机制中的功能研究胡 葭中国科学院武汉病毒研究所生物被膜在沙门菌逃逸肠黏膜树突状细胞免疫监视中的作用和机制阴银燕扬州大学结核分枝杆菌广谱胁迫蛋白Rv1996介导的异烟肼耐药机制研究胡新玲中国科学院微生物研究所一种肺炎链球菌磷壁酸合成相关蛋白的功能鉴定吴凯峰遵义医学院NO调控鲍曼不动杆菌多重耐药的机制研究邓珊珊成都医学院禾谷镰刀菌FgPrp6调控剪接体激活的作用机制研究金巧军西北农林科技大学高渗胁迫下应激活性蛋白激酶afSakA对黄曲霉毒素合成的调控机制研究袁 军福建农林大学低铁环境下白色念珠菌核质转运受体Nmd5调控转录因子Sef1异常核输出的分子机制研究黄新华中国科学院上海巴斯德研究所Fasciclin-1结构蛋白在红色毛癣菌生长发育和侵袭宿主细胞中的作用机制研究占 萍南昌大学生命科学第28卷1496白念珠菌灰菌细胞的菌丝生长调控机制研究管国波中国科学院微生物研究所水稻条纹病毒Pc2蛋白在病毒侵染介体中的功能研究赵淑玲扬州大学南方水稻黑条矮缩病毒(SRBSDV) P7-1形成的管状结构进入细胞壁的机制及其在病毒运动网络中的功能谢 礼浙江省农业科学院反向长链非编码RNA调控NIA基因响应CMV侵染的机制研究赵建华中国科学院微生物研究所植物液泡脂质调控CMV病毒基因组复制的机制顾周杭浙江理工大学Dicer-2调控抗病毒Toll免疫通路的作用机制研究王赵玮武汉大学利用MDV载体递呈靶向ALV-J的CRISPR/Cas9系统预防MDV和ALV-J感染的研究李 凯中国农业科学院哈尔滨兽医研究所NS1蛋白与人核仁磷酸化蛋白1互作在A型流感病毒感染过程中的作用研究朱春玉辽宁大学乙型肝炎病毒HBx类泛素化Neddylation修饰的功能及生物学意义刘宁宁中国科学院微生物研究所Nrf2/ARE信号通路对RHDV感染中肝脏氧化应激的作用胡 波江苏省农业科学院汉滩病毒核衣壳蛋白核质转运信号的鉴定及其在病毒感染中功能的研究叶 伟中国人民解放军第四军医大学免疫缺陷病毒Vpx蛋白识别宿主CRL4 (DCAF1) E3泛素连接酶的作用机制研究魏 伟吉林大学新型EV71病毒抑制剂靶向病毒3D蛋白的作用机制及其抗病毒功能研究张 伟苏州大学乙型肝炎病毒C蛋白促进Src/PI3K/Akt通路介导的肝细胞周期进程及机制刘 伟三峡大学基于"HCV core-ZEB相互作用促进EMT"探讨HCV感染相关性肝癌的发生机制张利军重庆医科大学抗肿瘤化合物tyrphostin AG490抑制丙型肝炎病毒NS5B聚合酶活性的分子机理研究杨 娜中国科学院海洋研究所GADD45G蛋白通过组蛋白修饰调控HSV-1感染的分子机理研究陈晓庆广州医科大学新型H3N2人流感的进化及受体结合特性研究路希山中国科学院天津工业生物技术研究所用VLP表达HPIV3的HN和F蛋白及其介导的免疫应答对小鼠的保护作用研究张光媛重庆医科大学CCHFV囊膜蛋白Gc结构域III与病毒入侵相关位点研究张怀东中国科学院武汉病毒研究所乙型脑炎病毒RNA元件的宿主特异性研究刘思情中国科学院武汉病毒研究所HIV-1利用CypA躲避TRIM11加速脱衣壳作用的机制研究袁 婷中国科学院武汉病毒研究所ISlncRNA-23促进HIV-1复制的机制研究董银慧中国科学院生物物理研究所发热伴血小板减少综合征病毒对IL-10和IL-11抗炎信号的抑制机制宁云佳中国科学院武汉病毒研究所HCMV通过IE1调控神经干细胞中Hes1表达及节律的机制李小军中国科学院武汉病毒研究所柠檬酸铁的抗病毒作用与机制研究王洪斌中国科学院上海巴斯德研究所长链非编码RNA A VIRL在抗流感病毒感染中的作用机制研究陈玉海中国科学院微生物研究所噬菌体SWU1杀菌过程中ROS的作用及产生机制研究樊祥宇济南大学gp38蛋白决定大肠杆菌噬菌体Bp7宽宿主谱的机制张 灿青岛农业大学鹦鹉热嗜衣原体两个TMH家族蛋白结构与功能研究伍海英南华大学黑龙江立克次体表面蛋白与血管内皮细胞(宿主细胞)表面分子相互作用的研究齐 永中国人民解放军南京军区军事医学研究所2植物学NAC转录因子调控荔枝果柄离区细胞凋亡的机理研究李彩琴华南农业大学细胞壁组分与复合多层结构对小麦籽粒水分及品质调控机理的研究应瑞峰南京林业大学水稻极度矮化基因STD1的图位克隆与功能分析房静静中国农业科学院作物科学研究所复苏植物牛耳草DnaJ蛋白参与水分胁迫下叶绿体保护的作用机理刘 杰中国科学院植物研究所PRSL1通过蛋白磷酸酶PP1调控拟南芥植株形态的分子机理秦倩倩兰州大学拟南芥WUSCHEL互作因子WIC1调控茎端分生组织功能的机理研究周 超山东农业大学虎耳草科虎耳草属石荷叶组的分类学研究张卓欣华南农业大学国家自然科学基金委员会生命科学部2016年度资助项目第12期1497中国景天科山景天组的分类学研究孟世勇北京大学日本蛇根草和广州蛇根草及其近缘类群的系统分类学研究吴 磊中南林业科技大学万寿竹属的分类学和系统学研究朱鑫鑫信阳师范学院豆科甘草属分子系统发育与生物地理学研究段 磊中国科学院华南植物园苍山冷杉复合体的物种划分与气候响应模式研究邵毅贞河南农业大学泛喜马拉雅地区鼠尾草属的分类修订胡国雄贵州大学豆科长柄山蚂蝗属的分子系统学研究宋柱秋中国科学院华南植物园中国长篦藻属硅藻的分类修订及分子系统学研究刘 琪山西大学平叶多褶苔和变异多褶苔的物种界定以及洲际间断分布格局的形成原因师雪芹安徽师范大学绿藻门橘色藻目的系统分类学研究朱 欢中国科学院水生生物研究所北半球间断分布植物珊瑚菜的亲缘地理学和保护遗传学研究李密密江苏省中国科学院植物研究所东亚植物区系空间分化的分子机制——棣棠花和粉花绣线菊复合群的谱系地理学研究罗 冬中国科学院昆明植物研究所热带亚洲鞭苔属的系统分类学研究董珊珊华南农业大学溯祖方法在系统发育基因组学分析中的可靠性席祯翔四川大学基于ddRAD-seq技术的悬竹属时空演化格局研究张宪智西北农林科技大学银莲花属西南银莲花组和鹅掌草组的分子系统学研究张 煜湖南科技大学双盖蕨属(蹄盖蕨科)的系统发育和物种多样性分化研究卫 然中国科学院植物研究所小檗科(Berberidaceae)系统基因组学及叶绿体基因组进化研究孙延霞中国科学院武汉植物园鬼灯檠属的进化历史研究马祥光中国科学院昆明植物研究所浅苞橐吾、云南橐吾和大黄橐吾之间的自然杂交和基因渗入研究余姣君中国科学院昆明植物研究所芸薹属油菜类作物细胞质基因组单倍型的精细解析及其协同进化分析乔江伟中国农业科学院油料作物研究所荨麻科叶绿体系统发育基因组学研究吴增源中国科学院昆明植物研究所大豆脂肪酸脱氢酶FAD2家族酶活性差异的进化与功能研究赵 嫚浙江工业大学印度-西太平洋地区红树基因组渐渗与物种分化机制研究何子文中山大学青藏高原—蒙古高原—中亚地区砾玄参复合群的亲缘地理学研究王瑞红浙江理工大学小麦杀配子现象中存在的转录组变异及miRNAs在其中的调控作用研究白 琰哈尔滨师范大学水玉簪属(Burmannia L.)植物与丛枝菌根真菌协同进化研究赵中涛中国科学院华南植物园结合几何形态学方法探索泽泻科植物心皮的发育与进化黄岚杰湖北大学荠属种内叶形自然变异的研究杨 丽中国科学院植物研究所姜科唇瓣的发生、发育及其分子机理研究李秀梅广东省农业科学院农业生物基因研究中心同域分布老鹳草属植物花冠开口方向的分化机制王 慧华中农业大学Hlips在保护蓝藻光系统II免受氨损伤中的机制研究戴国政华中师范大学管藻黄素型LHCII参与的假根羽藻非光化学淬灭机制研究王文达中国科学院植物研究所D1蛋白周转过程的调控机制研究-以短命植物为例涂文凤中国科学院植物研究所糖基化调控水稻乙醇酸氧化酶与过氧化氢酶互作及过氧化氢信号发生的机理研究张智胜湖南农业大学水稻镉吸收和积累相关基因的发掘和功能鉴定杨 猛华中农业大学硫-TOR信号通路介导拟南芥生长的分子机理研究徐 萍中国科学院上海生命科学研究院木薯碱性/中性转化酶MeNINV1的酶活性调节机制研究姚 远中国热带农业科学院热带生物技术研究所拟南芥转录因子NAC103在逆境胁迫响应中的生物学功能及其调控机理孙 玲江苏大学基于ssRNA-seq的木薯抗旱lncRNA的挖掘及相关基因调控网络的研究丁泽红中国热带农业科学院热带生物技术研究所BHLH转录因子HBI1调控植物生长和免疫抗病动态平衡的分子机制研究樊 敏山东大学转录因子基因LbCPC参与二色补血草盐腺分化的功能研究袁 芳山东师范大学GSK3类蛋白激酶SGK1通过SOS2调节植物耐盐性的分子机制研究周华鹏四川大学生命科学第28卷1498拟南芥内质网膜蛋白ROOT HAIR DEFECTIVE 3 (RHD3)调控花青素代谢分子机理王 静北方民族大学大豆转录因子SNAC的无序序列区对耐盐相关基因表达调控的分子基础刘国宝深圳大学光周期下拟南芥CAT2蛋白降解的分子机制研究苏 彤山东师范大学拟南芥类受体蛋白激酶CRKN1在脱落酸信号转导中的功能研究梁 杉清华大学一氧化氮调控拟南芥体内硼含量稳态的分子机制研究夏金婵河南中医学院植物高温响应的表观记忆机制刘军钟中国科学院上海生命科学研究院拟南芥光信号转录因子FAR1与EDS1互作调控植物免疫的机理研究王晚晴北京联合大学非传统G蛋白及其激活蛋白的晶体结构与水稻应答盐胁迫机制的关系解析苗 锐福建农林大学拟南芥MYB102通过调控细胞壁扩展蛋白的表达增强植物耐旱的分子机理研究周 成安徽科技学院大豆高盐响应蛋白GmOSM的调控机理研究万 群江苏省农业科学院拟南芥新型液泡阴离子通道(VSAC1和VSAC2)介导细胞水势调控的分子机制研究张海纹北京市农林科学院野生番茄响应昆虫唾液中FAC诱导物的遗传基础研究申国境中国科学院昆明植物研究所膜结合转录因子NAC091调控植物内质网胁迫应答的分子机理研究杨正婷贵州师范大学拟南芥丝氨酸羧肽酶SCPL41基因在干旱胁迫中的作用及机制贾艳霞中国科学院昆明植物研究所小麦类钙调素调节植物耐盐性的功能研究周 硕河北省农林科学院遗传生理研究所拟南芥RopGEF7的互作蛋白eIF4E1参与生长素介导的植物发育的分子机制刘太波华南农业大学拟南芥CKRW1调节内源生长素水平稳态平衡的分子机理研究武 磊兰州大学拟南芥乙酰转移酶HLS1在BR与Auxin协同调控植物生长发育中的功能研究刘晓磊中国科学院上海生命科学研究院GA与BR共同调控拟南芥纤维素合成的分子机制研究王 昕沈阳大学独脚金内酯信号通路D53-like SMXLs下游转录因子的鉴定与功能分析王 冰中国科学院遗传与发育生物学研究所一氧化氮与细胞分裂素信号通路互作调节植物适应性生长的分子机制张燕香中国科学院遗传与发育生物学研究所转录因子GhKNOX1-1调控棉花叶片形态建成的分子机制研究肖光辉陕西师范大学拟南芥WRKY71转录因子调控叶片衰老的分子机制研究于延冲青岛农业大学RLF1亚硝基化介导生长素调控水稻侧根发育的分子机制孙爱珍中国科学院上海生命科学研究院油菜生长素合成相关基因BnaA.YUCCA6调控分枝角度的机理研究成洪涛中国农业科学院油料作物研究所类受体激酶SIT1调节水稻叶片衰老的分子机制王 耕河北师范大学拟南芥隐花色素CRY2在介导蓝光依赖的生物钟调控中的作用机理研究曹世江福建农林大学周质微丝在植物网格蛋白介导内吞中的功能研究范路生福建农林大学IDD5的O-GlcNAc糖基化修饰在赤霉素信号转导通路中的作用KihyeShin福建农林大学甲基茉莉酸响应的bHLH类转录因子在青蒿素生物合成中的调控机制研究沈 乾上海交通大学糖基转移酶UGT78H2的活性鉴定及在黑莓类黄酮代谢中的功能分析陈 清四川农业大学玉米糖基转移酶UFGT2调节黄酮合成与耐逆的功能研究李燕洁山东大学FtMYB2对荞麦类黄酮生物合成的代谢调控机制及抗逆功能研究李晓华华中农业大学金柑类黄酮糖基转移酶基因功能分析及其调控网络构建刘小刚西南大学何首乌中芪合酶、白藜芦醇羟化酶基因的功能研究生书晶广东第二师范学院拟南芥LMBD2基因突变回复mur3-3表型的分子机制丁安明中国农业科学院烟草研究所博落回根中苄基异喹啉生物碱合成、转运和积累的细胞类型特异性定位及分子机制研究郑亚杰湖南农业大学拟南芥核质反向信号参与表观遗传调控的分子机理沈 杰中国科学院植物研究所灵芝三萜酸下游合成路径关键CYP450s基因挖掘与功能分析陈方方中国科学院武汉植物园文冠果性别分化的内源激素与microRNA调控机制敖 妍北京林业大学一个CCCH锌指蛋白调控水稻雄性生殖发育中胼胝质代谢的研究方瑞秋华南农业大学转录因子TDF1对拟南芥分泌型绒毡层发育与功能的转录调控楼 悦上海师范大学。
2010年全国优秀博士学位论文提名论文名单
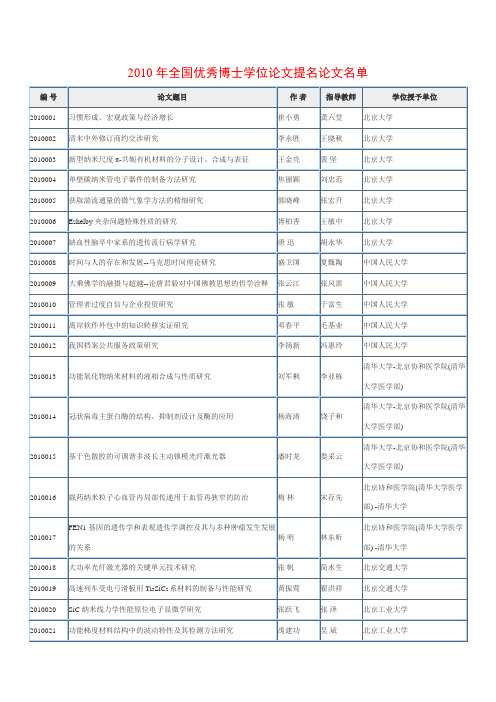
李晔
东北师范大学
2010087
非线性偏微分方程的渐近极限
张国敬
张凯军
东北师范大学
2010088
锂离子电池及其电极材料的研究
谢海明
王荣顺
东北师范大学
2010089
两种果蝠对植物种子的传播及行为学研究
唐占辉
盛连喜
东北师范大学
2010090
福柯微观权力思想研究--兼论马克思哲学的微观视域
赵福生
衣俊卿
简水生
北京交通大学
2010019
高速列车受电弓滑板用Ti3SiC2系材料的制备与性能研究
黄振莺
翟洪祥
北京交通大学
2010020
SiC纳米线力学性能原位电子显微学研究
张跃飞
张泽
北京工业大学
2010021
功能梯度材料结构中的波动特性及其检测方法研究
禹建功
吴斌
北京工业大学
2010022
形变碳纳米管的结构及晶格振动特性研究
Perelman的泛函在Ricci流中一些新的应用
张振雷
方复全
南开大学
2010062
高性能铌酸锂晶体与光电器件产品化的关键技术研究
孙军
许京军
南开大学
2010063
两亲性树枝状嵌段共聚物的合成及其超分子自组装研究
杨淼
王维
南开大学
2010064
头孢噻肟钠结晶技术研究
张海涛
王静康
天津大学
2010065
流动的土地--明清以来黄河小北干流区域社会研究
郁惠蕾
许建和
华东理工大学
2010122
新型环糊精分子梭的合成及性能研究
马骧
适于玉米杂交种纯度鉴定的SSR核心引物的确定及纯度鉴定图谱绘制

农业生物技术学报 Journal of Agricultural Biotechnology 2007,15(6):964~969*基金项目: 北京农业育种基础研究创新平台、 北京市自然科学基金项目 (No.YZPT0206)、 北京市科技新星项目 (No.2003B23) 和吉林省科技 计划项目 (No.20030555) 资助。
**通讯作者。
Author for correspondence.赵久然, 研究员, 主要从事玉米遗传育种及品种鉴定研究。
Email:<jiuran@>; 戴景瑞, 院士, 教授, 主要从事玉米遗传育种研究。
Email:<daijr@>. 收稿日期:20061121 接受日期:20070131 ·研究论文· 适于玉米杂交种纯度鉴定的 SSR 核心引物的确定 *王凤格 1,2 , 赵久然 1 **,王 璐 1 , 易红梅 1 , 郭景伦 1 , 戴景瑞 2**, 原亚萍 3 , 卢柏山 1 ,杨国航 1 (1.北京市农林科学院玉米研究中心, 北京 100097; 2.中国农业大学农学与生物技术学院, 北京 100094;3.吉林大学植物科学学院, 长春 130062)摘要: 为了确定一套适于玉米杂交种纯度鉴定的核心 SSR 引物, 利用 10 个 SSR 引物对 420 份已知玉米 ( ) 杂交种进行 DNA 指纹分析。
综合考虑多态性和杂合率两个指标,确定 bnlg161和bnlg1450为玉米杂交种纯度鉴定的首选核心引 物,利用这两个引物进行筛选,412 个品种 (占 98%) 能够找到具有杂合带型的鉴定引物, 确定 5 个引物 bnlg439、 bnlg125、 umc2105、 umc1705和 bnlg1792为玉米杂交种纯度鉴定的备选核心引物。
微卫星DNA技术评估绵羊品种的遗传多样性

微卫星DNA技术评估绵羊品种的遗传多样性侯冠彧;周汉林;王东劲【期刊名称】《热带农业工程》【年(卷),期】2016(40)4【摘要】利用30个SSR标记对乌珠穆沁羊、苏尼特羊、内蒙古细毛羊、宁夏滩羊、小尾寒羊和无角多赛特羊6个绵羊品种共237个体进行遗传多样性分析表明:6个绵羊品种平均期望基因多样度为0.671 1、观察基因多样度为0.565 9、平均多态信息含量为0.596 1;6个绵羊品种间遗传分化系数为11.74%;系统发生树将6个绵羊品种划为4个分支,即苏尼特羊和乌珠穆沁羊、内蒙古细毛羊和小尾寒羊、宁夏滩羊、无角多赛特羊.【总页数】7页(P33-39)【作者】侯冠彧;周汉林;王东劲【作者单位】中国热带农业科学院热带作物品种资源研究所海南儋州 571737;中国热带农业科学院热带作物品种资源研究所海南儋州 571737;中国热带农业科学院热带作物品种资源研究所海南儋州 571737【正文语种】中文【中图分类】S826【相关文献】1.利用微卫星DNA标记分析7个绵羊群体遗传多样性与系统发育 [J], 成述儒;罗玉柱;韩建林;Olivier Hanotte2.绵羊mtDNA D-环序列与微卫星DNA遗传多样性和系统发育分析 [J], 成述儒;罗玉柱;韩建林;Olivier Hanotte;胡江3.八个绵羊品种遗传多样性分析及引进肉用品种与地方品种杂种优势预测 [J], 毛杨毅;张李俊;罗惠娣;郭慧慧;狄冉;李希;张丽;周胜花;李春艳;马月辉4.中国西部七个地方绵羊群体微卫星DNA的遗传多样性研究 [J], 吕慎金;马月辉;杨燕;耿社民5.9个绵羊品种微卫星DNA遗传多样性分析 [J], 娄渊根;刘丑生;刘建学;刘刚;韩旭因版权原因,仅展示原文概要,查看原文内容请购买。
基于高通量测序的鹿角杯形珊瑚微卫星标记的开发
Vol. 45 No. 4Aug. 2020第45卷第4期2020年8月广西大学学报(自然科学版) Journal of Guangxi University (Nat Sci Ed)doi :10.13624/j. cnki. issn. 1001-7445. 2020.0958基于高通量测序的鹿角杯形珊瑚微卫星标记的开发罗燕秋53/,黄雯13,余克服*1,2,3,李明1,2,3,武茜】,2,3,王英辉1,2,3 ,李扬VS 4 ,陶永健I",陈胤民1,2,3(1.广西大学珊瑚礁研究中心,广西南宁530004;2.广西南海珊瑚礁研究重点实验室,广西南宁530004;3.广西大学海洋学院,广西南宁530004;4.广西大学林学院,广西南宁530004)摘要:为了更深入开展珊瑚遗传结构及其多样性研究,本研究采用高通量测序对鹿角杯形珊瑚进行微卫星标 记的开发。
首先用人工白化的方法获得污染较低的鹿角杯形珊瑚,然后使用该样品DNA,基于高通量测序平 台对鹿角杯形珊瑚简化基因组进行了测序,最终设计得到鹿角杯形珊瑚微卫星引物2 786对。
利用海南三亚 鹿角杯形珊瑚对其中6对多态性较高的微卫星标记进行了评价。
每个微卫星位点的有效等位基因数(Ne)为2. 198 -5.000;观测杂合度(Ho)为0.531 -0. 875;期望杂合度(He)为0. 512 -0. 816;Shannon 多样性指数为0. 856 -1. 829;多态信息含量(PIC)为0. 441 -0. 777,其中有4对微卫星标记呈现为高度多态性(PIC> 0. 5) ,2 对微卫星标记为中度多态性。
经过Bonferroni 校正后,只有微卫星标记LB11偏离哈迪一温伯格平衡;连锁不 平衡检验结果表明,位点之间不存在连锁不平衡现象。
关键词:鹿角杯形珊瑚;人工白化;高通量测序;微卫星标记;多态性中图分类号:Q38 文献标识号:A 文章编号:1001-7445 (2020) 04-0958-08Development of microsatellite in Pocillopora damicomisusing high-throughput sequencingLUO Yan-qiu 1*2'3'4, HUANG Wen 1'2'3, YU Ke-fu*1*2'3, LI Ming 1 >213, WU Qian 1'2'3,WANG Ying-hui 1'2'3, LI Yang 1'2'3'4, TAO Yong-jian 1'2'3, CHEN Yin-min 1'2'3(1. Coral Reef Research Center of China, Guangxi University , Nanning 530004, China ;2. Guangxi Laboratory on the Study of Coral Reefs in the South China Sea,Guangxi University , Nanning 530004, China ;3. School of Marine Sciences , Guangxi University, Nanning 530004, China ;4. Forestry College , Guangxi University , Nanning 530004, China)Abstract : To get a further study in coral genetic structure and diversity , this study used high-收稿日期:2019-02-11 ;修订日期:2019-04-18基金项目:国家自然科学基金资助项目(41866006);广西重大专项(桂科AA17204074);广西自然科学基金资助项目(2016GXNSFBA380028,2017GXNSFAA198196);广西重点研发计划项目(桂科 AB17129022);中国博士后基金资助项目(2016M592606,2017T100663)通讯作者:余克服(1969—),男,湖北公安人,广西大学教授,博士生导师;E-mail : kefuyu@ scsio. ac. cn o引文格式:罗燕秋,黄雯,余克服,等.基于高通量测序的鹿角杯形珊瑚微卫星标记的开发[j].r 西大学学报(自然科学版),2020,45(4) :958-965,第4期罗燕秋等:基于高通量测序的鹿角杯形珊瑚微卫星标记的开发959throughput sequencing to the development of Pocillopora damicomis microsatellite.Artificial bleaching was conducted to get the less contaminated coral genomic DNA.Then,we obtained 2786microsatellite loci after next generation sequencing.This study examined6polymorphic markers in Sanya P.damicornis population.Results revealed that effective allele number of these markers ranged from 2.198to5.000per locus,the observed heterozygosity value is from0.531to0.875,the expected heterozygosity value ranged from0.512to0.816.The Shannon indexranged from0.856to 1.829and the polymorphic information content(PIC)of these loci changed from0.441to0.777.There are4high polymorphic loci and2moderate polymorphic locus.Only LB11loci was deviated from Hardy-Weinberg equilibrium after Bonferroni correction but no significant linkage disequilibria was detected between the comparisons of these loci.Key words:Pocillopora damicornis;artificial bleaching;microsatellite;polymorphic;high-throughput sequencing0弓I言造礁石珊瑚是低等无脊椎动物,生活于热带浅海底,在动物学上属于刺胞动物门珊瑚虫纲六放珊瑚亚纲石珊瑚目,种类繁多⑷。
2013年度上海海洋大学大学生创新计划项目立项公示名单
詹晖东(1139124)
周应祺
教授
43
高频GPS系统监测钱塘江潮对杭州湾两岸的影响
蔚铁成
1120124
吴景刚(1120123)
周扬(1226131)
黄波宁(1220128)
徐利存(1120132)
朱卫东
沈蔚
讲师
副教授
44
大学生翻转课堂创新与实践
谢慧
1226103
乔敏(1226102)
田野(1226126)
杨志金(1329526)
沈蔚
朱瑞芳
副教授
讲师
45
基于ARCGIS的海图浏览器设计
李佩原
1126228
黄志鹏(1156127)
赖忠云(1191241)
汪璐丹(1069118)
郭立新
朱瑞芳
副教授
讲师
46
基于SAR数据的地物信息提取——以浙江衢山岛为例
金旭晨
1126125
周扬(1226131)
陈绍坚(1311216)
许丹
讲师
15
用多个核基因标记对刀鲚、湖鲚进行物种界定
孙佳丽
1213209
王文祥(1213224)
夏瑾(1213132)
胡景(1311227)
李晨虹
教授
16
海水酸化和温度对厚壳贻贝早期发育及能量收支的影响
黄茜枝
1210111
刘佳敏(1210109)
李红伊(1210101)
胡佳佳(1391410)
苏腾(1129236)
闫鹏坤(1129219)
管卫兵
汪振华
副教授
讲师
41
基于渔获统计的中西太平洋岛国渔业资源可持续利用评价
T细胞条件性敲除Spi1基因小鼠的繁育及鉴定
Fabricationandperformanceevaluationofhigh modulusandhigh strengthsilkfibroinguidedboneregenerationmembraneLiaoXiaoyu1,FangHui1,YangFeiyu1,ZouDuohong1,2(1College&HospitalofStomatology,AnhuiMedicalUniversity,KeyLaboratoryofOralDiseasesResearchofAnhuiProvince,Hefei 230032;2ShanghaiNinthPeople'sHospital,ShanghaiJiaoTongUniversitySchoolofMedicine,ShanghaiJiaoTongUniversitySchoolofStomatology,ShanghaiKeyLaboratoryofStomatology,Shanghai 200011)Abstract Objective TodevelopahighmodulusandhighstrengthbiodegradablesilkfibroinGBRmembranetoaddresstheissueofmaintainingthespaceforboneregenerationintherepairofosseousdefects.Methods Afterpurifyingsilkfibroinprotein,membranematerialswerepreparedusingevaporation hotpressingmethod.Thephysi calandchemicalpropertiesandbiologicalperformanceofthemembraneswereevaluatedusingstretchingtests,invitrosimulations,andcellco culturingmethods.Results AsilkfibroinGBRmembranewassuccessfullyfabrica ted,resultinginasimulateddegradationrateof35 3%after12hinvitro.Thewet stateelasticmodulusreached45MPa,whilethetensilestrengthreached8 39MPa.Furthermore,thecellsurvivalratewasnearly100%after7days.Conclusion ThebiodegradableGBRmembraneproducedinthisstudypossesseshighmodulusandstrength,aswellasexcellentbiocompatibility,offeringapromiseasafoundationforaddressingthebonedefectre pairandbonespacemaintenance.Keywords bonedefectrepair;maintainingthespaceforboneregeneration;guidedboneregenerationmem brane;silkfibroin;evaporation heatpressingmethod;osteogenicdifferentiation;tensilestrength网络出版时间:2024-04-1213:52:17 网络出版地址:https://link.cnki.net/urlid/34.1065.R.20240410.1008.006T细胞条件性敲除Spi1基因小鼠的繁育及鉴定王卉卉,朱向玲,吴旭铭,张慧茹,周园园,王安琪,刘 崇,涂佳杰摘要 目的 繁育T细胞条件性敲除Spi1基因的小鼠并对其进行鉴定,为进一步探索Spi1编码蛋白PU 1的作用提供研究基础。
基于Illumina MiSeq高通量测序技术分析不同保质期鸡蛋干的微生物群落多样性
24㊀2020Vol.46No.24(Total 420)DOI:10.13995/ki.11-1802/ts.024560引用格式:宋思家,黄子彧,林莹莹,等.基于Illumina MiSeq 高通量测序技术分析不同保质期鸡蛋干的微生物群落多样性[J].食品与发酵工业,2020,46(24):24-30.SONG Sijia,HUANG Ziyu,LIN Yingying,et al.Microbial community diversity in egg curds with different shelf life revealed by Illumina MiSeq sequencing[J].Food and Fermentation Industries,2020,46(24):24-30.基于Illumina MiSeq 高通量测序技术分析不同保质期鸡蛋干的微生物群落多样性宋思家1,黄子彧2,林莹莹1,郭慧媛1∗1(中国农业大学食品科学与营养工程学院,精准营养与食品质量重点实验室,教育部功能乳品重点实验室,北京,100083)2(中国农业大学食品质量安全北京实验室,北京,100083)摘㊀要㊀为探究造成鸡蛋干腐败变质的核心优势微生物及造成鸡蛋干保质期差异的微生物种类,为鸡蛋干的防腐保鲜提供理论依据,采用Illumina MiSeq 高通量测序技术对保质期不同的3组腐败变质鸡蛋干的菌群组成进行分析㊂结果表明,3组鸡蛋干样品在门水平的优势菌群均为厚壁菌门,该菌门在15d 组㊁22d 组和29d 组的丰度占比分别为96.73%㊁87.55%和70.94%;在属水平,15d 组㊁22d 组和29d 组中相对丰度最高的菌属均为芽孢杆菌属,其在各组中的占比分别为33.14%㊁47.38%和68.85%,因此,芽孢杆菌属是鸡蛋干的核心致腐微生物㊂此外,造成鸡蛋干保质期差异的菌属包括乳杆菌属和狭义梭菌属,它们可加速鸡蛋干的腐败变质,是鸡蛋干保质期的限制因素,应在杀菌防腐环节重点关注㊂关键词㊀鸡蛋干;Illumina MiSeq 测序;微生物多样性;芽孢杆菌属;乳杆菌属;狭义梭菌属Microbial community diversity in egg curds with different shelf liferevealed by Illumina MiSeq sequencingSONG Sijia 1,HUANG Ziyu 2,LIN Yingying 1,GUO Huiyuan 1∗1(Key Laboratory of Precision Nutrition and Food Quality,Key Laboratory of Functional Dairy (Ministry of Education),College of Food Science and Nutritional Engineering,China Agricultural University,Beijing 100083,China)2(Beijing Laboratory for FoodQuality and Safety,China Agricultural University,Beijing 100083,China)ABSTRACT ㊀To investigate the dominant bacteria that caused the deterioration of egg curd and the mi-crobial species that caused the difference in shelf life of egg curd,Illumina MiSeq sequencing was ap-plied to analyze the microbial community diversity in three groups of spoiled egg curd with different shelf life.The results indicated that Firmicutes were the dominant phylum in these three groups of egg curd,and the abundances of Firmicutes in the 15d group,22d group and 29d group were 96.73%,87.55%and 70.94%,respectively.At the genus level,the most abundant bacteria in the 15d group,22d group,and 29d group were Bacillus ,which accounted for 33.14%,47.38%,and 68.85%in each group,respectively.Therefore,Bacillus was the most dominant spoilage bacteria in egg curd.In addi-tion,the dominant spoilage bacteria also included Lactobacillus and Clostridium _sensu _stricto _13,which can accelerate the deterioration of egg curd.They were the main bacteria that caused the shelf life differ-ence of egg curd and should be paid close attention to in the sterilization process.Key words ㊀egg curd;Illumina MiSeq sequencing;microbial community diversity;Bacillus ;Lactobacil-lus ;Clostridium _sensu _stricto _13第一作者:博士研究生(郭慧媛副教授为通讯作者,E-mail:guohuiyuan@)㊀㊀基金项目:国家重点研发计划课题(2018YFD0400305)收稿日期:2020-05-26,改回日期:2020-09-02㊀㊀鸡蛋干是中国市面上常见的休闲蛋制品,因其营养丰富㊁风味独特而深受消费者喜爱㊂依据GB 2749 2015‘食品安全国家标准蛋与蛋制品“内容,在保质期内的鸡蛋干在感官方面应具有产品正常的色泽㊁滋味㊁气味,无异味,且具有产品正常的形状㊁形态㊁无酸败㊁霉变等;在微生物限量方面,其菌落总数应<104CFU/g,大肠菌群应<10CFU/g[1]㊂目前在实际生产过程中,鸡蛋干的保质期为9个月㊂鸡蛋干的生产原料以全蛋液或蛋清液为主,在加工制作过程中极易受到微生物污染而导致产品腐败变质[2]㊂相关调查研究表明,预包装鸡蛋干即便经过高压蒸汽灭菌后也可能出现胀包㊁汤汁浑浊㊁有异味的变质现象,尤其在夏秋高温季节,鸡蛋干发生腐败变质的概率更高[3]㊂最近数十年,鸡蛋干的市场在不断扩大,因此对鸡蛋干的防腐保鲜开展研究愈显重要㊂而明确鸡蛋干的优势致腐微生物,是防腐的首要关键㊂然而目前对于鸡蛋干的腐败微生物群落多样性的报道十分有限,贺燕等[3]从腐败鸡蛋干中分离鉴定得到2株芽孢杆菌,袁先铃等[4]在腐败鸡蛋干中检测到彭氏变形菌和奇异变形菌㊂因此,开展鸡蛋干中腐败微生物群落多样性的研究具有重要意义㊂对于微生物群落多样性的研究,长期以来主要是基于‘伯杰细菌鉴定手册“,对微生物菌落进行分离㊁纯化,进而通过形态学观察和一系列复杂的生理生化实验进行鉴定㊂这种传统方法不但耗费时间和精力,而且无法对微生物进行精确鉴定,不能检测到微生物群落多样性的真正概貌[5-6]㊂因为有些微生物的培养条件非常苛刻,甚至不可培养,常规方法难以检测到不可培养的微生物㊂近年来,在微生物群落多样性研究中已经越来越倾向于采用Illumina MiSeq高通量测序技术[7-10]㊂高通量测序技术也称 下一代测序技术 ,可同时测序数百万个核酸分子,根据16S rDNA基因序列分析特定样品中微生物群落的构成情况[11-12]㊂目前高通量测序技术已成为监测食品贮藏过程中微生物群落演替㊁分析发酵及腐败食品微生物群落结构等食品微生物多样性研究的首选方法[11]㊂本研究以保质期不同的腐败变质鸡蛋干为研究对象,基于Illumina Miseq测序平台对其16S rDNA V3~V4进行扩增子测序分析,研究其微生物群落结构组成和相对丰度,探究造成鸡蛋干腐败变质的核心优势微生物以及造成鸡蛋干保质期差异的微生物种类,为靶向抑制鸡蛋干的腐败变质提供理论依据㊂1㊀材料与方法1.1㊀材料与试剂鸡蛋,北京德青源农业科技股份有限公司;细菌基因组DNA提取试剂盒,美国Omega科技公司;Axy Prep DNA凝胶回收试剂盒,美国Axygen公司;2ˑTaq PCR MasterMix㊁DNA Marker,天根生化科技(北京)有限公司㊂1.2㊀仪器与设备5418R高速离心机,德国Eppendorf公司;拍击式均质机,法国Interscience公司;RS-400型真空包装机,北京日上公司;T100Thermal Cycler聚合酶链式反应(polymerase chain reaction,PCR)仪㊁PowerPac Basic型电泳仪,美国Bio-Rad公司;ChampGel5000增强型全自动凝胶成像仪,北京赛智创业科技有限公司;Illumina MiSeq测序仪,美国Illumina公司㊂1.3㊀试验方法1.3.1㊀样品制备鸡蛋干的制作工艺参照参考文献[13],具体为取预处理蛋液于模具中,水浴加热使其中心温度达到80ħ,并维持20min,待其冷却收缩后脱模㊂将鸡蛋干样品真空包装后,进行90ħ㊁30min灭菌,于25ħ保存直至变质㊂预实验发现,对于使用不同批次原料㊁于不同的时间点制作的鸡蛋干样品,即使其加工工艺完全一致,样品的保质期也会有较大差异㊂为探究不同保质期的鸡蛋干样品中菌群结构的差异,采用上述加工工艺,分别使用不同批次原料,于不同的时间点制作了多批样品,从其中选取贮藏15d㊁22d和29d腐败变质的3批鸡蛋干产品,取样保存于-80ħ冰箱待测㊂将3批鸡蛋干样品分别编号为15d㊁22d 和29d,每组样品设置3个平行㊂1.3.2㊀细菌总DNA的提取参考高乾坤等[14]的方法,对样品进行处理后,采用美国Omega公司的细菌基因组DNA提取试剂盒,提取细菌总DNA,提取步骤参照试剂盒产品说明书,并进行1%(质量分数)的琼脂糖凝胶电泳,以检测提取效果㊂1.3.3㊀PCR扩增及高通量测序以检验合格的基因组DNA为模板,扩增细菌16S rDNA基因V3~V4区㊂正向引物为338F(5ᶄ-ACTCCTACGGGAGGCAGCA-3ᶄ),反向引物为806R (5ᶄ-GGACTACHVGGGTWTCTAAT-3ᶄ)㊂所得PCR产物经2%(质量分数)琼脂糖凝胶电泳检测扩增效果,2020年第46卷第24期(总第420期)25㊀26㊀2020Vol.46No.24(Total 420)然后将样品进行纯化并定量检测㊂将检测合格的样品送至上海美吉生物医药科技有限公司进行Illumina MiSeq 高通量测序㊂1.4㊀数据分析对MiSeq 测序得到的双端序列数据进行处理,首先将成对的reads 拼接成一条序列,同时对reads 的质量和拼接的效果进行质控过滤㊁得到有效序列,并校正序列方向[15]㊂基于操作分类单元(operational taxonomic units,OTU)聚类分析结果,研究腐败变质鸡蛋干样品中微生物的多样性,通过Alpha 多样性检测单样本测序深度,并分析微生物群落的丰度和多样性[16]㊂基于分类学信息,在各个分类水平上进行群落结构统计,综合讨论保质期不同的鸡蛋干样品中微生物群落结构组成和相对丰度㊂2㊀结果与分析2.1㊀测序数据统计与OTU 分析采用Illumina Miseq 高通量测序获得了3种不同保质期鸡蛋干样品的原始序列数据后,经Fastp 及FLASH 软件优化数据后得到了9个样本共529384条有效序列,序列平均长度为409.70~429.92bp,主要集中在401~440bp(图1-a)㊂稀释曲线可反映在高通量测序过程中的样品取样深度,因此是评价测序数据量是否全面覆盖微生物群落的常用指标[14]㊂为了探究鸡蛋干样品的测序数据量是否科学,从测序获得的所有原始数据中随机抽取数据,并统计其物种多样性指数,得到稀释曲线(图1-b 和图1-c)㊂结果表明,随着数据量的增加,OTU 曲线和Shannon-Wiener 曲线的趋势已经逐渐平缓,即再增加数据量a -样品序列长度分布图;b -OTU 曲线;c -Shannon-Wiener 曲线图1㊀样品序列长度分布图㊁OTU 曲线和Shannon-Wiener 曲线Fig.1㊀Length distribution profile of trimed sequences,OTU curves and Shannon-Wiener curves对于发现新物种也无显著影响㊂样本的OTU 覆盖度已经基本饱和,更多的数据量对发现新OTU 的边际贡献很小,这个结果说明测序数据量足够大,可以反映样本中绝大多数的微生物多样性信息[17]㊂2.2㊀Alpha 多样性分析根据97%相似性水平下的OTU 信息,采用Alpha 多样性指标的Shannon㊁Simpson㊁ACE㊁Chao1及Cov-erage 指数对鸡蛋干样品微生物物种的丰富度和多样性进行评估[14]㊂如表1所示,3组样品的Coverage指数都达到了0.999以上,即样品文库的覆盖率足够高,有未被检出序列的可能性较小,表明本次试验所建文库能够较准确地反映出待测鸡蛋干的微生物多样性情况㊂Chao1㊁ACE 指数用于计算菌群丰度,二者的值越大,说明样品的群落丰富度越高;Shannon㊁Simpson 指数用于计算菌群多样性指数,Shannon 的值越大,说明样品群落多样性越高,而Simpson 指数值越大,则说明其群落多样性越低[18]㊂22d 组的鸡蛋干样品ACE㊁Chao1及Shannon 指数最高,Simpson 值最低,这说明该组样品的细菌群落丰富度及多样性高于其他2组样品,可能由于该组样品的原料中及加工过程中污染的细菌种类最多;29d 组鸡蛋干样品的ACE㊁Chao1及Shannon 指数均为3组中最小,这说明了该组样品中细菌群落丰度及多样性最低,可能由于该组样品的原料中及加工过程中受到的污染程度较低㊂综上,经Alpha 多样性指数统计分析可知,鸡蛋干样品的物种多样性由高到低依次为22d 组㊁15d 组和29d 组㊂此结果显示,相比于污染的微生物物种的多样性,鸡蛋干所污染微生物的具体种类可能与鸡蛋干的保质期相关性更大,这与孙同辉等[11]的报道相一致㊂2.3㊀OTU 的Veen 分析基于样品总OTU 数作Veen 图,利用Venn 图可以展示多组样品间共有和每组样品特有的OTU 数目,并直观展示样品间OTU 的重叠情况[19]㊂如图22020年第46卷第24期(总第420期)27㊀所示,15d 组样品与22d 组样品共有的OTU 数107个,15d 组与29d 组共有的OTU 数131个,22d 组与29d 组共有的OTU 数92个,3组样本共有的OTU 数为81个,占各样本OTU 数比例较小,而各样本特有的OTU 数在113~259,表明各样本之间的菌群结构差异性较大㊂表1㊀Alpha 多样性指数Table 1㊀Alpha diversity indexes样品名称Shannon 值Simpson 值ACE 值Chao1值Coverage 值15d 0.370.91120.69122.720.999722d 1.640.52138.52139.080.999829d0.340.8585.1682.550.9997图2㊀不同保质期样品中微生物物种间OUT 分析图Fig.2㊀OUT analysis diagram of microbial species insamples with different shelf life2.4㊀不同保质期鸡蛋干样品细菌群落结构分析2.4.1㊀基于门水平的细菌菌群结构分析在门水平上,3组鸡蛋干样品的群落组成如图3所示㊂15d 组的鸡蛋干样品总细菌组成以厚壁菌门(Firmicutes)和变形菌门(Proteobacteria)为主,相对丰度分别为96.73%和1.96%㊂除此之外还包括少量的放线菌门(Actinobacteria )和拟杆菌门(Bacte-roidetes)微生物㊂而22d 组的鸡蛋干样品菌群结构以厚壁菌门㊁变形菌门和拟杆菌门为主,相对丰度分别为87.55%㊁9.41%和1.32%,同时还含有少量的放线菌门微生物㊂29d 组的鸡蛋干样品中微生物以厚壁菌门(70.94%)㊁放线菌门(27.81%)和变形菌门(1.09%)为主,还含有少量的拟杆菌门微生物㊂以上结果表明,在3组不同保质期的样品中,厚壁菌门的丰度均为最高,可见厚壁菌门是腐败变质鸡蛋干中的优势微生物㊂近年来许多研究表明,厚壁菌门微生物是高蛋白食品中的优势腐败菌,在乳制品㊁肉制品及水产品中都十分常见[18-21],这与本研究的结果一致㊂图3㊀基于门分类水平上的样品菌群分布图Fig.3㊀Bacterial distribution pattern at the phylum level2.4.2㊀基于科水平的细菌菌群结构分析基于科分类水平的3组鸡蛋干样品的微生物群落结构如图4所示㊂测序结果表明,15d 组的鸡蛋干样品中类芽孢杆菌科(Paenibacillaceae)所占比例最大,为47.43%,其次为芽孢杆菌科(Bacillaceae)和梭菌科(Clostridiaceae_1),所占比例分别为33.14%和14.96%,还有乳杆菌科(Lactobacillaceae)㊁伯克氏菌科(Burkholderiaceae)㊁链球菌科(Streptococcaceae)㊁假单胞菌科(Pseudomonadaceae)等相对丰度较低的菌种,说明鸡蛋干在制作和包装过程中污染微生物的种类复杂,导致样品菌落结构呈现多样性㊂22d 组的鸡蛋干样品菌群结构以芽孢杆菌科㊁乳杆菌科㊁链球菌科㊁伯克氏菌科为主,相对丰度分别为47.39%㊁29.11%和3.76%㊁3.29%,除此之外还包括少量的红杆菌科(Rhodobacteraceae)㊁假单胞菌科㊁类芽孢杆菌科微生物㊂29d 组鸡蛋干样品中的菌群在科水平上主要有芽孢杆菌科(68.85%)和小单孢菌科(Mi-cromonosporaceae,27.60%),还含有少量的假单胞菌科和伯克氏菌科微生物㊂以上结果表明,不同保质期鸡蛋干样品所含微生物种类和数量有明显差异,如相比于其他2组,29d 组样品中却未检出乳杆菌科微生物,可能由于该组样品的原料中及加工过程中污染的乳杆菌科较少㊂29d 组鸡蛋干样品的保质期最长,其微生物多样性低于其余2组,说明样品中污染的微生物种类和数量与鸡蛋干的保质期密切相关㊂22d 组样品微生物多样性最高,但此组保质期不是最短,可能由于样品中含有丰度较高的乳杆菌科微生物[22-23],对其他微生物产生拮抗作用㊁抑制其生长繁殖,相对延长了鸡蛋干保质期㊂此外,在15d 组样品中检测出梭菌科微生物,而其他2组中均未检出,梭菌科中有些菌种为厌氧菌,可在真空包装鸡蛋干中迅速生长繁殖,这可能是本组鸡蛋干保质期远小于其他2组的原因㊂3组不同保质期的样品中,均含有芽孢28㊀2020Vol.46No.24(Total 420)杆菌科微生物,且丰度较高,表明芽孢杆菌科是腐败变质鸡蛋干中的核心优势微生物㊂图4㊀基于科分类水平上的样品菌群分布图Fig.4㊀Bacterial distribution pattern at the family level2.4.3㊀基于属水平的细菌菌群结构分析选取在鸡蛋干样品丰度排名前15的属,根据其在每组鸡蛋干样品中的丰度信息聚类绘制成热图(图5-a)㊂热图可通过颜色变化直观地反映出各组鸡蛋干样品群落组成的相似性和差异性,更便于发现在各组样品中分布较多或较少的菌属[10]㊂图中颜色越偏砖红色,表明样品中所含该菌属比例越高,颜色越偏深蓝色,比例则越低㊂如图5-a 所示,在属水平,22d 组和29d 组聚为一类,15d 组单独聚为一类,说明22d 组和29d 组菌群整体结构相似度较高,与15d 组相似度较低,表明鸡蛋干样品的保质期与其菌群结构相关性较大㊂从图5-b 各样品菌群柱状分布图中可以看出,在属水平上,15d 组优势菌群主要是芽孢杆菌属(Bacillus ,33.14%)㊁短芽孢杆菌属(Brevibacillus ,30.96%)㊁类芽孢杆菌属(Paenibacillus ,16.46%)和狭义梭菌属(Clostridium _sensu _stricto _13,14.86%)㊂在优势菌群中,芽孢杆菌属所占比例最大,它属于需氧或兼性厌氧菌,此属中大部分细菌为腐生菌,广泛分布于自然环境中㊂该菌属可形成芽孢,对不良环境条件有很强的抵抗力,不易杀灭㊂贺燕等[3]在腐败鸡蛋干中也有检测到芽孢杆菌属微生物㊂短芽孢杆菌属为需氧或兼性厌氧菌,近年来相关报道多集中于该属工程菌株的研究,鲜见有研究发现其在食品腐败中的作用[24-25]㊂类芽孢杆菌属为需氧或兼性厌氧菌,近年来许多研究表明,其为多种乳制品和即食食品的腐败微生物[26-27]㊂狭义梭菌属广泛分布于土壤㊁下水污泥㊁人和其他哺乳动物的肠道等环境中,是乳制品和肉制品中常见的腐败微生物[20,28-29]㊂22d 组优势菌群主要是芽孢杆菌属(47.38%)㊁乳杆菌属(Lactobacillus ,28.26%)和链球菌属(Strep-tococcus ,3.57%)㊂乳杆菌属为兼性厌氧菌,广泛分布于自然界中,污染乳制品㊁肉制品㊁水产品等高蛋白食品后可引起腐败变质㊂链球菌属为兼性厌氧菌,该属中有少数种为腐生菌,污染食品后可引起腐败变质[29]㊂a -样品菌群相对丰度热图;b -样品菌群相对丰度柱形图图5㊀基于属分类水平上的样品菌群相对丰度热图和柱形图Fig.5㊀Heat map and stacked-column of relative abundances ofbacterial communities at the genus level29d 组优势菌属主要是芽孢杆菌属(68.85%)和小单孢菌属(Micromonospora ,27.60%)㊂小单孢菌属为好氧或微好氧的腐生菌,是土壤和水域的常见微生物群体[30],其在真空包装鸡蛋干中占比达到27.60%,可能由于鸡蛋干中初始污染小单孢菌属较多,且微好氧微生物能在较低的氧分压下生长,鸡蛋干虽然经过真空包装,但仍无法将样品中氧气完全排出,因此小单孢菌属中部分微好氧菌种可以生长㊂以上结果表明,在3组不同保质期的样品中,芽孢杆菌属的相对丰度均为最高,可见芽孢杆菌属是鸡蛋干的核心致腐微生物㊂保质期最长的29d组样品中优势菌属为芽孢杆菌属和小单孢菌属,其中小单孢菌属为好氧或微好氧菌,在真空包装鸡蛋干中生长缓慢,而芽孢杆菌属中部分菌种为兼性厌氧菌,可在真空包装条件下良好生长,因此芽孢杆菌属应为限制此组样品保质期的微生物㊂而相比于29d组,22d组样品中还含有较高丰度的乳杆菌属,这可能是造成本组鸡蛋干保质期<29d的原因㊂此外,狭义梭菌属仅在保质期为15d的鸡蛋干样品中检测到,它属于厌氧菌,因此在真空包装鸡蛋干中的生长速度大于非厌氧菌,可快速分解鸡蛋干中营养物质造成腐败变质,这可能是15d组鸡蛋干保质期远低于其他2组的原因㊂3㊀结论本研究以保质期不同的3组腐败变质鸡蛋干为研究对象,采用Illumina Miseq高通量测序技术分析样品中菌群组成情况㊂结果表明,3组腐败变质鸡蛋干的微生物的多样性以及丰度存在较大差异,而总体来看,厚壁菌门为腐败变质鸡蛋干的优势菌门,芽孢杆菌科为优势菌科㊂在属水平,芽孢杆菌属在3组样品中的相对丰度均为最高,是鸡蛋干的核心致腐微生物㊂此外,造成鸡蛋干保质期差异的菌属包括乳杆菌属和狭义梭菌属,它们可加速鸡蛋干的腐败变质,是鸡蛋干保质期的限制因素,应在杀菌防腐环节重点关注㊂高通量测序技术检测不同保质期鸡蛋干样品的微生物组成,不仅可以分析鸡蛋干腐败过程中的核心优势菌群,还可以高效快速地分析样品中微生物种类的差异,为今后鸡蛋干质量安全监测及靶向研究鸡蛋干防腐保鲜技术提供理论依据㊂参考文献[1]㊀GB2749 2015食品安全国家标准蛋与蛋制品[S].北京:中国标准出版社,2017.GB2749 2015National standards of food safety eggs and egg products[S].Beijing:China Standards Press,2017.[2]㊀薛艳蓉,茂文,赵瑞生,等.鸡蛋在储存过程中微生物及蛋品质的变化研究[J].畜牧与饲料科学,2017,38(8):67-69;72.XUE Y R,MAO W,ZHAO R S,et al.Dynamic changes of microbio-logical and qualitative features of chicken eggs during storage process [J].Animal Husbandry and Feed Science,2017,38(8):67-69;72.[3]㊀贺燕,谭纯良,李娜,等.复配防腐剂抑制鸡蛋干腐败菌效果研究[J].食品与机械,2019,35(11):159-164.HE Y,TAN C L,LI N,et al.Study on the effect of compound pre-servatives on inhibiting spoilage of egg curd[J].Food and Machinery, 2019,35(11):159-164.[4]㊀袁先铃,黄丹,潘明.腐败鸡蛋干中主要微生物的分离纯化及鉴定[J].食品研究与开发,2012,33(7):57-59.YUAN X L,HAUNG D,PAN M.Purification and identification of microbes in corruption dried eggcurd[J].Food Research and Devel-opment,2012,33(7):57-59.[5]㊀HANDELSMAN J.Metagenomics:Application of genomics to uncul-tured microorganisms[J].Microbiology and Molecular Biology Re-views,2004,68(4):669-685.[6]㊀吴燕燕,钱茜茜,李来好,等.基于Illumina MiSeq技术分析腌干鱼加工过程中微生物群落多样性[J].食品科学,2017,38(12):1-8.WU Y Y,QIAN Q Q,LI L H,et al.Microbial community diversity in dried-salted fish during processing revealed by Illumina MiSeq se-quencing[J].Food Science,2017,38(12):1-8.[7]㊀POOŁKA J,REBECCHI A,PISACANE V,et al.Bacterial diversityin typical Italian salami at different ripening stages as revealed by high-throughput sequencing of16S rRNA amplicons[J].Food Mi-crobiology,2015,46(4):342-356.[8]㊀ZHANG J B,LI Y,LIU X C,et al.Characterization of the microbialcomposition and quality of lightly salted grass carp(Ctenopharyng-odon idellus)fillets with vacuum or modified atmosphere packaging [J].International Journal of Food Microbiology,2019,293(6):87-93.[9]㊀ZANG J H,XU Y S,XIA W S,et al.Dynamics and diversity of mi-crobial community succession during fermentation of Suan yu,a Chi-nese traditional fermented fish,determined by high throughput se-quencing[J].Food Research International,2018,111(9):565-573.[10]㊀戴玲瑛,齐曼婷,王立梅,等.虾酱中间产物细菌群落结构及多样性[J].食品科学,2018,39(22):173-178.DAI L Y,QI M T,WANG L M,et al.Bacterial community struc-ture and diversity in unsterilized shrimp sauce[J].Food Science,2018,39(22):173-178.[11]㊀孙同辉,陈洁,李雪琴,等.杀菌方式对年糕微生物多样性及品质的影响[J/OL].食品科学,2020./kc-ms/detail/11.2206.TS.20200109.0934.012.html.SUN T H,CHEN J,LI X Q,et al.Effect of sterilization method onmicrobial diversity and quality of rice cake[J/OL].Food Science,2020./kcms/detail/11.2206.TS.20200109.0934.012.html.[12]㊀SOON W W,HARIHARAN M,SNYDER M P.High-throughput se-quencing for biology and medicine[J].Molecular Systems Biology,2013,9(1):640.[13]㊀张伯男,樊晓盼,马俪珍,等.风味鸡蛋干的加工工艺研究[J].食品研究与开发,2018,39(11):101-105.ZHANG B N,FAN X P,MA L Z,et al.Study on processing tech-nology of flavor egg curd[J].Food Research and Development,2018,39(11):101-105.[14]㊀高乾坤,焦琳舒,杜贺超,等.高通量测序分析不同产地带鱼冷藏时微生物群落多样性[J].食品科学,2018,39(18):127-132.GAO Q K,JIAO L S,DU H C,et al.High-throughput sequencingdifferent waters during chilled storage[J].Food Science,2018,39(18):127-132.[15]㊀ANDREEVSKAYA M,JOHANSSON P,LAINE P,et al.Genome2020年第46卷第24期(总第420期)29㊀30㊀2020Vol.46No.24(Total 420)sequence and transcriptome analysis of meat-spoilage-associated lactic acid bacterium Lactococcus piscium MKFS47[J].Applied andEnvironmental Microbiology,2015,81(11):3800-3811.[16]㊀LU X M,LU P Z.Characterization of bacterial communities in sedi-ments receiving various wastewater effluents with high-throughput se-quencing analysis[J].Microbial Ecology,2014,67(3):612-623.[17]㊀江杨阳.小龙虾低温贮藏品质变化规律及其腐败菌鉴定[D].杭州:浙江大学,2019.JIANG Y Y.Studies on the quality changes of red swamp crayfish and its specific spoilage organisms during low temperature storage[D].Hangzhou:Zhejiang University,2019.[18]㊀马岩石,姜明,李慧,等.基于高通量测序技术分析东北豆酱的微生物多样性[J].食品工业科技,2020,41(12);100-105.MA Y S,JIANG M,LI H,et al.Analysis of microbial diversity of northeast soy sauce based on high-throughput sequencing technolo-gy[J].Science and Technology of Food Industry,2020,41(12):100-105.[19]㊀李春生,王悦齐,李来好,等.传统鱼露发酵过程中细菌群落演替及对其挥发性风味形成的影响分析[J].食品科学,2018,39(24):85-92.LI C S,WANG Y Q,LI L H,et al.Bacterial community succes-sion during fermentation of Chinese fish sauce and its effect on for-mation of volatile flavor components[J].Food Science,2018,39(24):85-92.[20]㊀毛海萍,袁开,金仁耀,等.基于传统分离培养和高通量测序分析市售咸鳓鱼中微生物多样性[J].食品研究与开发,2019,40(21):193-201.MAO H P,YUAN K,JIN R Y,et al.Study of microbial diversity in commercial salted Chinese herring using culture isolation and high-throughput sequencing[J].Food Research and Development,2019,40(21):193-201.[21]㊀李伟程,侯强川,于洁,等.传统发酵乳制品中微生物多样性研究[J].食品工业科技,2018,39(1):131-136.LI W C,HOU Q C,YU J,et al.Study on the diversity of microbi-al in traditional fermented dairy products[J].Science and Technol-ogy of Food Industry,2018,39(1):131-136.[22]㊀孙群,易华西,张冬,等.潜在益生菌的筛选及抑菌功能特性评价[J].食品科技,2019,44(12):1-5.SUN Q,YI H X,ZHANG D,et al.Screening of potential probiot-ics and evaluation of antibacterial function characteristics[J].FoodScience and Technology,2019,44(12):1-5.[23]㊀王玉文,刘英华.乳酸菌的生物拮抗作用[J].中国乳品工业,2008,36(2):55-57.WANG Y W,LIU Y H.State antagonism about lactic acid bacteria [J].China Dairy Industry,2008,36(2):55-57.[24]㊀赵乃昕,苑广盈.医学细菌名称及分类鉴定[M].济南:山东大学出版社,2013.ZHAO N X,YUAN G and Classification of Medical Bac-teria[M].Jinan:Shandong University Press,2013.[25]㊀邹亮,吴敬,陈晟.重组蔗糖异构酶在短短芽孢杆菌中的表达及发酵优化[J].食品与生物技术学报,2019,38(1):22-28.ZOU L,WU J,CHEN S.Expression and fermentation optimization of recombinant sucrose isomerase in Brevibacillus choshinensis [J].Journal of Food Science and Biotechnology,2019,38(1):22-28.[26]㊀HELMOND M,NIEROP GROOT M N,HERMIEN B V D V.Characterization of four Paenibacillus species isolated from pasteur-ized,chilled ready-to-eat meals[J].International Journal of FoodMicrobiology,2017,252(13):35-41.[27]㊀TRMCIC A,MARTIN N H,BOOR K J,et al.A standard bacterialisolate set for research on contemporary dairy spoilage[J].Journal of Dairy Science,2015,98(8):5806-5817.[28]㊀AKHTAR S,PAREDES-SABJA D,TORRES J A,et al.Strategyto inactivate Clostridium perfringens spores in meat products [J].Food Microbiology,2009,26(3):272-277.[29]㊀何国庆,贾英民,丁立孝.食品微生物学[M].北京:中国农业大学出版社,2009.HE G Q,JIA Y M,DING L X.Food Microbiology[M].Beijing:China Agricultural University Press,2009.[30]㊀杨苏声.细菌分类学[M].北京:中国农业大学出版社,1997.YANG S S.Bacterial Taxonomy [M ].Beijing:China AgriculturalUniversity Press,1997.。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
热带海洋学报 JOURNAL OF TROPICAL OCEANOGRAPHY 2010年 第29卷 第3期:47−54 http://jto.scsio.ac.cn; http://www.jto.ac.cn
收稿日期:2008-03-27; 修订日期:2008-07-21。刘学东编辑 基金项目:国家高技术研究发展计划(“863”)项目(2006AA10A409), 国家科技支撑计划项目(2006BAD01A13)和广东省科技计划项目 (2006B 20201063)联合资助. 作者简介:曲妮妮(1982—), 女, 山东省青岛市人,硕士研究生, 从事海洋生物遗传育种研究. E-mail: qunini0605@163. com 通讯作者:E-mail: pearlydh@163.com; pearlydh@pub.gz.gd.cn
海洋生物学 基于FIASCO技术的合浦珠母贝微卫星标记分离与筛选研究 曲妮妮1,2, 龚世园2, 黄桂菊1, 童金苟3, 喻达辉1 (1. 中国水产科学研究院南海水产研究所, 广东 广州 510300; 2. 华中农业大学水产学院, 湖北 武汉 430070; 3. 中国科学院水生生物研究所,湖北 武汉 430072)
摘要: 用FIASCO(Fast Isolation by AFLP of Sequences Containing Repeats)技术开展了合浦珠母贝Pinctada fucata基因组微卫星标记的分离与筛选研究。合浦珠母贝基因组DNA经限制性内切酶MseI酶切后与接头连接, 用生物素标记的(CA)15探针与其杂交, 然后用磁珠富集、洗脱获得单链目的片段, 经PCR扩增后形成双链, 最后进行克
隆转化, 构建微卫星富集文库。挑选克隆用探针引物(CA)15 和载体引物进行第2次筛选, 获得阳性克隆357个, 测
序结果表明, 297个克隆(83.2%)含有微卫星序列, 包括479个微卫星DNA结构域。其中完美型微卫星有370个(77.3%), 非完美型95个(19.8%), 复合型14个(2.9%)。合成引物49对, 有31对(63%)扩增出目的产物, 其中9 对在种群中(n=32)具有扩增多态性, 其多态信息含量PIC值在0.375―0.809之间, 平均为0.536; 等位基因数 在2―9个之间, 平均为4.889个; 观测杂合度介于0.200―0.600之间, 平均为0.415; 期望杂合度的变化范围为0.454―0.844, 平均为0.598。表明FIASCO技术适合于合浦珠母贝微卫星标记的分离与筛选。 关键词: FIASCO法; 合浦珠母贝Pinctada fucata; 微卫星标记; 分离与筛选
中图分类号: S91 文献标识码: A 文章编号: 1009-5470(2010)03-0047-08
Isolation and screening of microsatellite markers from the Chinese pearl oyster Pinctada fucata based on FIASCO
QU Ni-ni 1,2, GONG Shi-yuan 2, HUANG Gui-ju 1, TONG Jin-gou 3, YU Da-hui 1 (1. South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510300; 2. College of Fisheries, Huazhong Agricultural University, Wuhan 430070; 3. Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072)
Abstract: Isolation and screening of microsatellite markers from Pinctada fucata were performed using the method of Fast
Isolation by AFLP of Sequences Containing Repeats (FIASCO). After restriction of genomic DNA with MseI, the digested fragments were ligated with adaptors and then hybridized with biotinylated (CA)15 probes. The tentative microsatellite DNA was isolated by streptavidin-coated magnetic beads from the hybridized mixture. After purification, the isolated microsatellite DNA was amplified using degenerated primer MseⅠ-N and then cloned into T-vector. After transforming, the microsatel-lite-enriched library was constructed. The second PCR screening was performed using the primer of (CA)15 probe and T vec-
tor’s primers, and 357 positive clones were obtained. Sequencing analysis showed that 297 clones (83.2%) contained microsa-tellite DNA, including 479 microsatellite domain. Among them, 370 microsatellites (77.3%) were perfect type, 95 (19.8%) imperfect type and 14 (2.9%) compound type. Forty nine pairs of primers were designed and 31 (63%) pairs were effective for PCR amplification. Nine loci showed polymorphism as tested by a population (n=32). PIC values for the nine loci ranged from 0.375 to 0.809 with an average of 0.536. The number of alleles ranged from 2 to 9 with an average of 4.889. The observed heterozygosity ranged from 0.200 to 0.600 with an average of 0.415, and the expected heterozygosity ranged from 0.454 to 0.844 with an average of 0.598. This study demonstrated that FIASCO is a useful technique of microsatellite isolation for Pinctada fucata. 48 热带海洋学报 Vol. 29, No. 3 / May, 2010 Key words: FIASCO method; Pinctada fucata; microsatellite marker; isolation and screening
自1989年微卫星标记报道以来[1–3], 由于其高度多态性和共显性特点, 得到广泛应用, 其分离技术也不断发展改进。早期的分离方法主要是小片段DNA克隆法[4], 该方法是将基因组DNA进行部分酶切, 分离小片段DNA构建文库, 然后进行测序筛选。由于这种方法耗时费力且筛选效率很低[4], 于是提出了构建微卫星富集文库的思路。Ostrander等[5]报道了引物延伸富集法, Karagyozov等[6]提出了滤膜富集法, Kijas等[7]提出了磁珠富集法, 其核心环节是酶切产物经PCR扩增后通过与生物素标记的探针杂交, 然后用磁珠富集, 获得大量富含微卫星序列的片段并构建文库供后续筛选。Lunt等[9]提出了基于RAPD-PCR的微卫星分离技术(PCR isolation of microsatellete arrays, PIMA)。Zane等[8]报道了基于AFLP技术的磁珠富集快速分离技术(Fast Isolation by AFLP of Sequences Containing Repeats, FIASCO), 即按照AFLP的方法进行酶切、连接、扩增后再利用磁珠杂交富集含有微卫星的DNA片段。FIASCO法因操作简单, 效率高, 是目前微卫星分离最快速的技术。匡刚桥等[10]用此法构建了鳜鱼的基因组微卫星富集文库, 得到18个多态标记; Liao等[11]用FIASCO法分离了铜鱼的微卫星标记, 得到9个多态位点。但目前尚未见用FIASCO法开展双壳贝类微卫星分离的研究报道。 合浦珠母贝(Pinctada fucata), 又叫马氏珠母贝, 是我国生产海水珍珠的重要贝类。但经过40多年的人工繁殖-养殖循环生产后出现了养殖性状退化的现象, 一些学者利用RAPD[12]、AFLP[13]等显性标记对其种质资源进行了研究, 然而利用共显性标记进行研究的报道很少, 其原因就是缺乏共显性标记。微卫星标记由于其多态性高, 是最好的共显性标记之一, 目前已广泛用于种质资源和遗传育种等研 究[14–15]。但目前双壳贝类的微卫星研究报道不多, 国内只有虾夷扇贝Pecten yesoensis [16]、栉孔扇贝Chlamys farrei[17]、长牡蛎Ostrea gigas[18]、翡翠贻贝Perna retzius[19]等, 国外也只有大珠母贝Pinctada maxima[20]、珠母贝P. margaritifera[21]、太平洋牡蛎Crassostrea gigas[22]、欧洲牡蛎Ostrea angulata [23] 、美洲牡蛎C. virginica[24]等。最近, 佟广香等[25]用磁珠富集法进行了马氏珠母贝的微卫星分离研究, 得到8个多态位点。本研究采用FIASCO技术进行了合浦珠母贝微卫星分离与筛选研究, 以期为合浦珠母贝微卫星标记的分离筛选提供快速、有效的技术手段。