核酶的发现与应用
9核糖体1.比较Ⅰ型内含子与Ⅱ型内含子在结构与剪接上有什么不同?答:I

9核糖体1. 比较Ⅰ型内含子与Ⅱ型内含子在结构与剪接上有什么不同?答:I型内含子转录后可以形成9个由碱基配对形成的特定二级结构,分别命名为P1至P9,P1和P7是保守的。
I型内含子具有自我剪接的功能,在剪接反应中,要有一种鸟嘌呤核苷(含有游离的3'-OH)G-OH。
G首先结合到内含子的5'端,当线性的内含子成为环状时,其3'端可以距离5'端15个核苷酸以外,从而将原来的5'端和15个碱基(或以上)的节段(包括G)切除出去。
这种自我剪接,是由RNA的特定序列的核酸内切酶的活性所催化。
II型内含子主要存在于线粒体中的一类内含子,它的剪接位点类似于核编码结构基因的内含子,并同样遵从GT--AG规律。
剪接机理同核内含子的剪接相似,也要形成一个套索的中间体,通过形成5'-2'键将要剪接的位点靠近到一起。
但是,II型内含子的剪接又不完全与核内含子的剪接相同,它具有自我剪接的功能,不需要剪接体和snRNA的参与,也不需要ATP供能。
从结构上看,II型内含子的6个结构域可形成发夹环, 结构域5与6之间只间隔3个碱基,结构域6参与转酯作用。
2. 上游启动子与内部启动子差异答:DNA上的启动子区是转录起始前RNA聚合酶识别的一段碱基序列。
大多数基因的启动子区在编码区的上游,不过,对于由RNA聚合酶III转录的基因,启动子通常在转录起始点的下游,例如5S核糖体RNA 的基因,RNA聚合酶结合在转录起始位点的下游即DNA编码区内。
3. 反义RNA与核酶的区别答:反义RNA是指与mRNA互补的RNA分子, 也包括与其他RNA互补的RNA分子。
由于核糖体不能翻译双链的RNA,所以反义RNA与mRNA特异性的互补结合, 即抑制了该mRNA的翻译。
通过反义RNA 控制mRNA的翻译是原核生物基因表达调控的一种方式,最早是在E.coli 的产肠杆菌素的Col E1中发现的,许多实验证明在真核生物中也存在反义RNA。
近几年来通过人工合成反义RNA的基因, 并将其导入细胞内转录成反义RNA, 即能抑制某特定基因的表达,阻断该基因的功能, 有助于了解该基因对细胞生长和分化的作用。
第六章核糖体和核酶

1. 发现核糖体及核糖体功能鉴定的两个关键技术是什么?答:核糖体最早是Albert Claude于1930s后期用暗视野显微镜观察细胞的匀浆物时发现的,当时称为微体(Microsomes),直到1950s中期,George Palade在电子显微镜下观察到这种颗粒的存在。
当时George Palade和他的同事研究了多种生物的细胞, 发现细胞质中有类似的颗粒存在, 尤其在进行蛋白质合成的细胞中特别多。
后来Philip Siekevitz 用亚细胞组份分离技术分离了这种颗粒, 并发现这些颗粒总是伴随内质网微粒体一起沉积。
化学分析揭示, 这种微粒富含核苷酸, 随之命名为ribosome,主要成分是核糖体RNA(rRNA),约占60%、蛋白质(r蛋白质)约占40%。
核糖体的蛋白质合成功能是通过放射性标记实验发现的。
将细胞与放射性标记的氨基酸短暂接触后进行匀浆,然后分级分离,发现在微粒体部分有大量新合成的放射性标记的蛋白质。
后将微粒体部分进一步分离,得到核糖体和膜微粒,这一实验结果表明核糖体与蛋白质合成有关。
两个关键技术是亚细胞组份分离技术和放射性标记技术。
2•说明人体单倍体染色体组中四种rRNA基因的组成、排列方式和拷贝数。
答:在人基因组的四种rRNA基因中,18S、5.8S和28S rRNA基因是串联在一起的,每个基因被间隔区隔开,5S的rRNA基因则是编码在另一条染色体上。
前3个基因组成一组,分布在人的13、14、15、21、22 等5条染色体上。
在间期核中,所有这5条染色体rRNA基因区域,转录时聚集在一起,形成一个核仁。
在人体单倍体染色体组中,每组rRNA基因有200个拷贝。
每一拷贝为一个rDNA 转录单位。
这 3 个基因是纵向串联排列在核仁组织者的DNA 上。
真核细胞核糖体的5S rRNA基因则是独立存在于一个或几个染色体上,拷贝数达几千个。
在人的细胞中,该基因的拷贝有24000 个之多,它们串联排列在 1 号染色体接近末端处。
人教版高考生物新教材各种酶的最全归纳及拓展
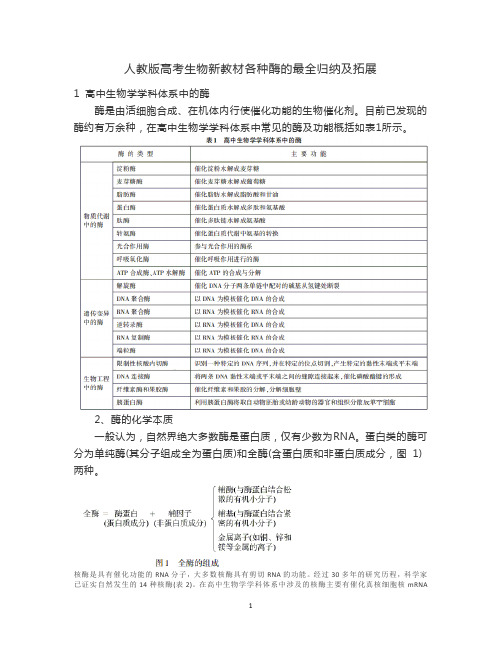
人教版高考生物新教材各种酶的最全归纳及拓展1 高中生物学学科体系中的酶酶是由活细胞合成、在机体内行使催化功能的生物催化剂。
目前已发现的酶约有万余种,在高中生物学学科体系中常见的酶及功能概括如表1所示。
2、酶的化学本质一般认为,自然界绝大多数酶是蛋白质,仅有少数为RNA。
蛋白类的酶可分为单纯酶(其分子组成全为蛋白质)和全酶(含蛋白质和非蛋白质成分,图1)两种。
核酶是具有催化功能的RNA分子,大多数核酶具有剪切RNA的功能。
经过30多年的研究历程,科学家已证实自然发生的14种核酶(表2)。
在高中生物学学科体系中涉及的核酶主要有催化真核细胞核mRNA前体剪接的剪接体和催化蛋白质生物合成的核糖体。
3酶作用的机制酶的作用机制是通过降低生化反应的活化能来提高反应速率。
目前该机制一般用中间产物学说来解释,其核心是酶在催化过程中首先与底物结合形成酶-底物中间复合物,发生化学反应后再分解成酶和产物,酶在反应前后数量和性质均不变。
4酶的作用特点酶的作用具有高效性,与无机催化剂的反应相比,酶促反应的速率一般要高1010~1012倍,甚至更高(表3)。
酶的作用具有专一性,酶对底物的选择具有严格的专一性,即一种酶只能作用于一种或一类底物,使其发生特定类型的化学反应,并产生特定的产物。
酶的催化活性依赖其空间结构的完整,一旦变性则会失去催化能力。
高温、高压、极端pH和重金属盐等都容易使酶失去催化活性。
故酶促反应要求在比较温和的条件下进行,如常温、常压等。
核酶在发挥作用时与上述起催化作用的蛋白质具有相似的特征,也有专一性,高效性和对温度、pH敏感等。
5关于酶专一性的假说酶作用的专一性源于酶在催化时存在活性中心与底物结合的过程。
酶的活性中心又称活性部位,是指酶分子中能直接同底物结合并起催化反应的空间部位(图2)。
5.1“锁钥”学说人教版高中生物学教材必修1“降低化学反应活化能的酶”一节课后习题中展示了酶作用专一性的“锁钥”学说。
生物化学I 第三章 酶学

根据国际生化协会酶命名委员会的规定,每一个酶都用 四个打点隔开的数字编号,编号前冠以EC(酶学委员会缩 写),四个数字依次表示该酶应属的大类、亚类、亚亚类 及酶的顺序号,这种编码一种酶的四个数字即是酶的标码。
例如:EC1.1.1.27(乳酸脱氢酶) 酶
乳酸:NAD+氧化还原
u u u u
第一大类 氧化还原酶 第一亚类 —CHOH被氧化 第一亚亚类 氢受体为NAD+ 排序 顺序号为27
4. 1878年, Kü hne赋予酶统一的名称 “Enzyme”, 其意思为“在酵母中”。
Enzyme 酶
德国生物化学家
5. 1930~1936年,Northrop和Kunitz先后得到了胃蛋 白酶、胰蛋白酶和胰凝乳蛋白酶结晶,并用相应方法 证ቤተ መጻሕፍቲ ባይዱ酶是蛋白质。
为此, Northrop和Kunitz于1949年共同 获得诺贝尔奖。
(1)旋光异构专一性:
(2)顺反异构专一性:
例如:不同的酶有不同的活性中心,故对底物有严格的特异性。例如乳 酸脱氢酶是具有立体异构特异性的酶,它能催化乳酸脱氢生成丙酮酸 的可逆反应:
A、B、C分别为LDH活性中心的三个功能基团
消化道内几种蛋白酶的专一性
氨肽酶
(芳香) (硷性)
羧肽酶 羧肽酶
(丙)
Ser
His 活性中心重要基团: His57 , Asp102 , Ser195
Asp
(4)酶的活性中心与底物形状不是正好互补的。
(5)酶的活性中心是位于酶分子表面的一个裂 缝(Crevice)内。
(6)底物通过次级键较弱的作用力与酶分子结 合,这些次级键为:氢键、离子键(盐键)、 范德华力和疏水相互作用。 (7)酶的活性中心具有柔性或可运动性。
酶工程第6章-脱氧核酶2012

图; 9.转染48h; 10. 转染60h; 11. 转染60h后的阴性对
照; 12. 转染72h。
利用核酶或脱氧核酶抑制有害基因的基本原理
对医疗应用来说最主要的还是寻找那些具 有切割特定RNA顺序的核酶,从而可以在体 内抑制某些有害基因。
基因治疗的概念出现在二十几年前,现在已 经在临床上得到了实际应用。基因治疗最早的临 床研究是1990年Blaese 等进行的对腺苷脱氨酶 (ADA)缺乏症的治疗,随后在对遗传病、病毒侵染、 肿瘤等疾病的治疗中得到广泛的应用。中国也是 开展基因治疗比较早的国家,1991年薛京伦等开 展了血友病B基因治疗的临床实验,并取得比较理 想的效果。
结构稳定。生理条件下DNA比RNA稳定106 倍,DNA的磷酸二酯键比蛋白质的肽键抗 水解能力要高100倍。
成本低廉、易于合成和修饰。 脱氧核酶具有催化效率高和高度专一性等
特性。
脱氧核酶(Deoxyribozyme,DRz)的分类
分类依据:借鉴国际酶学委员会对蛋白酶 的分类方法,将DRz 分成4 类:水解酶、转移 酶、合成酶和氧化酶 具有水解酶活性的DRz 1、作用于RNA 的DRz 主要包括水解RNA的“10-23”、“8-17”DRz。
手枪形脱氧核酶自我剪切作用机理
切割点
10
5‘ 20
3‘ C A
茎I(结合部位)
40
茎II (催化部位) 30
手枪形结构脱氧核酶的自身切割位点在第14nt处,其3’端约27个碱基对自身切割活 性的发挥至关重要。
“二分”型结构脱氧核酶
核酶治疗HIV感染的研究进展

状 结构 核酶 。
1 核酶 的结构 1 1 锤 头状结 构 .
一
锤头 状 核酶 困其底 物结 合后 呈现
种 锤 头状 结 构 而 得 名 。 见图 1 锤 状 核 酶 长 约 。
3 0个 核苷酸 。该 酶可 分 为 3个部 分 , 间 足 以 单 链 中 形式存 在 的 保 守 核 苷 酸 和 螺旋 Ⅲ组 成 的 催 化 核 心。 锤头 状 桉 酶 催 化 中 心 序 列 为 5’ uG GA c Au Guc c c uGA GA GA 3 。 两 侧 的 螺 旋 1 3为 核 酶 的 侧 G c A’ / 翼序列 , 根据底 物 序 列 不 同而 灵 活调 整 , 碱 基 配 可 以 对的方 式 与靶 R A 结 合 , 而 决 定核酶 作 用 的 特 异 N 从 性 。核 酶通 过侧翼 序 列识 别 底 物 GUA位 电, 后 催 然 化核 心切 割底物 R 3 端 的 G NA ’ UA三联 体 。
王福祥 孙 永 涛
人 免 疫 缺 陷 病 毒 ( ma mmu o e c n y Hu n i n d f i e ie vrsHI 是 引起 艾 滋病 的病 原 。HI 分 类 J属 于 i , v) u V 二 逆 转录 病毒科 慢病 毒亚科 。HI 有两 种类 型 : v V HI 1 HI 和 V一2 。其 中 HI 1 引起全 球 艾滋 病 流 行 V 是 的主要 病原 , 因此 , 目前 关 于 HI 1的 防治研 究 主 V 要 针对 HI V一1进行 的 。近年 来, 人们联 合使用 HI V 1蛋 白酶抑 制剂 , 核苷 类逆 转 录 酶抑 制剂 和 非 核 苷 类 逆转 录酶 抑 制剂 治 疗 HI V一1感 染 并 取 得 了令 人 鼓 舞 的进展 。然 而, V一1 染 仍 旧是 整个 人类 所 HI 感 要 面临 的难题 , 且新 近 又 出现 了耐 抗 HI 感 染 药 并 V 物 的病毒 株。 这就 需要 人们 努 力 寻 找一 项 新 的 治疗 策略 以 治疗 这 种 耐 药 HI 1感 染 。2 V O世 纪 9 O年 代初 人 们 发 现 核 酶 可 以 有 效 地 在 细 胞 培 养 中抑 制 HI 1的 繁殖 , 那 时 起通 过 基 因 治 疗 策 略, 酶 V 从 核 治疗 HI 1 染逐渐 进入 人类 临床 阶段 。 V 感 核酶 (i zme 是 一类具 有催 化 活 性 的 R roy ) B NA分 子, 通过碱基 配对特异地 与相应 的 R 可 NA 底物 结 合 , 特 异地 切割 靶 R 可 NA序 列 , 形象 地 称为 “ 子 被 分 剪 刀” 它 在抗病 毒 , 肿瘤等 领域 有 广泛的 应用 。本 , 抗 文对 核酶 的结构 , 作用 机 理 及 核 酶 在抗 HI 感 染 方 V 面 的应用 作一综述 。 在 18 9 2年, eh T C c R从 四 膜虫 r NA 前 体 的 加 R 工研 究 中首 先 发 现 r N 前 体 有 自我 剪接 作 用 , R A 即 RN A本身 也 有 催 化 作 用 , 提 出 了 r o y 并 i zme一 词 。 b
核酶和抗体酶

将不同的重链和轻链基因随机组合,克隆 到合适的表达载体中,在原核细胞表达不 同的抗体,形成一个抗体库,从这个抗体 库中,用抗原可以筛选到相应的抗体基因。
引入法
随着噬菌体抗体库技术的完善,可根据需 要构建适当序列的基因片断,绕过免疫学 方法,构建全新的抗体酶。 噬菌体展示技术将组建亿万种不同特异性 抗体可变区基因库和抗体在大肠杆菌中功 能性表达,与高效快速的筛选手段结合起 来,彻底改变了抗体酶生产的传统途径。
A. 酯酶的底 物–酯
B.酯的羧基碳原子 受到亲核攻击形成 四面体过渡态
C.设计的磷酸酯 类似物,作为抗原 去免疫实验动物
O –C –
磷酸酯类似物 免
(半抗原)
疫
对酯水解反应有 催化作用的单克
隆抗体
抗体酶用于有机酯的水解,过渡态类似 物磷酸盐和磷酸酯作为免疫原诱导产生 的单克隆抗体催化水解反应比未催化反 应快104倍。
L-19IVS
G- P
- OH +
15nt
P399nt
图 13- 四膜虫 35S RNA 内含子剪接 的转酯反应模型
L-19具有酶的主要特 征:专一性强,加快 反应速度,反应前后 酶分子保持不变
L-19 IVS所催化的水解反应和连接反应
异议
引入法
用基因工程方法改造和制备全新的抗体酶 是一种很有前途和发展潜力的抗体酶制备 方法。
将催化基因引入到特异抗体的抗原结合 位点上,使其获得催化功能。 也可以针对性地改变抗体结合区的某些 氨基酸序列,以获得高效的抗体酶。
引入法
对于已产生的单抗,分析抗体结合部位 的氨基酸顺序或对应的碱基顺序。 通过对抗体酶结合部位氨基酸对应的基 因序列进行定点突变,希望能在抗体结 合部位换上有催化作用的氨基酸。 改变抗体酶的催化效率。
核酶和抗体酶2

内含子
Ⅰ
型
5'外显子
5'
U pA
3'外显子
G pU
3'
内
第一次转酯反应
含
pG-OH
子
pGpA
的 5'
UOH
G pU
3'
自
我
第二次转酯反应
剪
5' pGpA
接
5'
U pU
3'
GOH 3'
I类内含子催化其他RNA分子 反应的几种类型
1、转核苷酸作用
2CpCpCpCpC CpCpCpCpCpC+CpCpCpC
• 这两个位点对于剪接是十分重要的,一旦发 生突变无论在体内还是在体外,会抑制剪接。
• 此法则几乎适合于所有真核生物的核基因, 这意味着它们切除内含子的机制是相同的, 但不适用于Ⅰ类内含子。
RNA的催化功能
• 核酶首先是美国 Colorado 大学Cech在研 究四膜虫rRNA剪接机制时发现的。
–一方面证明了四膜虫rRNA的剪接机制; –另一方面证明了L-19 分子的催化活性。
四膜虫rRNA内含子 ---Ⅰ型内含子
I型内含子的结构特点
1、拼接点序列为 5U··· ···G3
2、中部核心结构 3、内部引导序列 4、剪接通过转酯反应进行
引导序列
保守序列 G结合位点
剪接部位
Ⅰ型内含子二级结构通式
内部引导序列
• 内含子中可与外显子配对的序列称为内部引导序列 • 其作用是决定剪接的专一性。
+ 外显子1 外显子2 内含子1
•剪接产物通过凝胶电泳见到: rRNA前体 + + + + 核抽取物 - + - + GTP - + + -
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
U pA G pU 5'外显子3'外显子
内含子
核酶的研究进展
摘要: 80年代初,由美国科学家Cech 和Altman 发现了核酶,随着人类基因工程研究的深入,工作者和基础科学研究人员开始注意到核酶在各方面的应用潜力。
关键词: 概念 分类 剪接机制 反义核酸技术 医学上的应用 应用实例 技术问题
核酶的概念
核酶一词用于描述具有催化活性的RNA, 即化学本质是核糖核酸(RNA), 却具有酶的催化功能。
核酶的作用底物可以是不同的分子, 有些作用底物就是同一R NA 分子中的某些部位。
核酶的功能很多,有的能够切割RNA, 有的能够切割DN A, 有些还具有RNA 连接酶、磷酸酶等活性。
与蛋白质酶相比,核酶的催化效率较低,是一种较为原始的催化酶。
核酶的分类
剪接型核酶:这类核酶具有核酸内切酶和连接酶两种活性。
、作用机制:通过既剪又接的方式除去内含子 。
需要鸟苷酸或鸟苷及镁离子参与
剪接机制:
I 型内含子的结构特点: 1、拼接点序列为5’U··· ···G3’ 2、中部核心结构
3、内部引导序列
pGpA G pU 3'U 5'
OH
4、剪接通过转酯反应进行
剪切型核酶:这类核酶催化自身或者异体RNA的切割,相当于核酸内切酶。
这类RNA进行催化反应时只切不接。
类型:1) 自体催化剪切型2) 异体催化剪切型。
特点:在Mg 2+ 或其他二价金属离子存在下,在特定的位点,自我剪切,产生5‘-OH 和2’, 3‘-环磷酸二酯末端。
核酶的应用
核酶是在对多种植物病毒卫星RNA及类病毒RNA的自我剪接研究中发现的,数量较少,常见于rRNA的内含子。
核酶的具体作用主要有:
1.核苷酸转移作用。
2.水解反应,即磷酸二酯酶作用。
3.磷酸转移反应,类似磷酸转移酶作用。
4.脱磷酸作用,即酸性磷酸酶作用。
RNA内切反应,即RNA限制性内切酶作用。
核酸内切酶可以催化水解多核苷酸内部的磷酸二酯键。
有些核酸内切酶仅水解5′磷酸二酯键,把磷酸基团留在3′位置上,称为5′-内切酶;而有些仅水解3′-磷酸二酯键,把磷酸基团留在5′位置上,称为3′-内切酶。
能专一性地识别并水解双链DNA上的特异核苷酸顺序,称为限制性核酸内切酶。
当外源DNA侵入细菌后,限制性内切酶可将其水解切成片段,从而限制了外源DNA在细菌细胞内的表达,而细菌本身的D NA由于在该特异核苷酸顺序处被甲基化酶修饰,不被水解,从而得到保护。
限制性核酸内切酶可被分成三种类型。
Ⅰ型和Ⅲ型限制酶水解DNA需要消耗ATP,全酶中的部分亚基有通过在特殊碱基上补加甲基基团对DNA进行化学修饰的活性。
Ⅱ型限制酶水解DNA不需要ATP也不以甲基化或其它方式修饰DNA,能在所识别的特殊核苷酸顺序内或附近切割DNA。
因此,被广泛用于DNA分子克隆和序列测定。
核酶的具体应用实例
(一) 抗病治疗
随着对核酶的深入研究,已经认识到核酶在遗传病,肿瘤和病毒性疾病上的潜力。
位点切断RNA,使得它失去活性。
如果一个能专一识别HIV的RNA的核酶存
在于被病毒感染的细胞内,那么它就能建立抵抗入侵的第一防线。
甚至,HIV确
实进入到了细胞并进行了复制,RNA也可以在病毒生活史的不同阶段切断HIV
的RNA而不影响自身的RNA。
又如,白血病是造血系统的恶性肿瘤,目前尚缺
少有效的治疗方法。
核酶的发现,尤其是锤头状核酶,为白血病的基因治疗带来
了新的希望。
近些年,在国外的一些国家已经在小白鼠体内得到较好的效果。
(二)反义核酸技术
反义核酸反义核酸是指能与特定mRNA精确互补、特异阻断其翻译的RNA或D
NA分子。
利用反义核酸特异地封闭某些基因表达,使之低表达或不表达,这种技术即为反义核酸技术。
它包括反义RNA、反义DNA和核酶三大技术。
反义核酶作为一种基因下向调节作用因子,在抑制一些有害基因的表达和失控基因的过度表达上发挥着重要作用。
随着反义核酶技术的发展和成熟,已逐渐应用于抗某些人体寄生虫病的研究。
反义核酸目前有三种来源:一是利用固相亚磷酰胺法人工合成的短小反义寡聚核苷酸,这是反义核酸最普遍的应用方式,包括未修饰AON和硫代磷酸酯化(PS)、磷酸二酯化(PO)和甲基化等修饰AON二类,其中以PSAON应用最广泛;二是更具有实用价值的工人表达载体,包括单个基因和多个基因的联合反义表达载体,它是利用基因重组技术将靶基因序列反向持插入到载体的启动子和终止子之间,通过转录可源源不断产生反义RNA分子;三是天然存在的反义核酸分子,但目前分离纯化尚存在困难。
作用特点
1.高度特异性:反义核酸药物通过特异的碱基互补配对作用于靶RNA或DNA,犹如“生物导弹”。
2.高生物活性、丰富的信息量;反义核酸是一种携带特定遗传信息的信息体,碱
基排列顺序可千变万化,不可穷尽
3.高效性:直接阻止疾病基因的转录和翻译。
4.最优化的药物设计:反义核酸技术从本质上是应用基因的天然顺序信息,实际上是最合理的药物设计。
5.低毒、安全:反义核酸尚未发现其有显著毒性,尽管其在生物体内的存留时间有长有短,但最终都将被降解消除,这避免了如转基因疗法中外源基因整合到宿主染色体上的危险性
反义核酸技术与化妆品发展
反义核酸化妆品是在生物技术药物研究的基础上最新发展起来的,与传统化妆品相比,具有高效、高选择性、安全环保的特点。
反义核酸技术在化妆品领域异军突起,代表了化妆品发展的一个重要方向。
反义技术是应用碱基配对的原理,以体内表达某种特定蛋白质的靶基因为基础,人工设计一段与之互补的基因片段封闭该靶基因,直接阻断该蛋白质的产生。
针对有害基因,突变基因,非正常基因及其过度表达的基因,科学家设计了反义核酸,使这些基因关闭或者低表达。
反义核酸是人工合成的DNA片段(简称寡核苷酸),它与待封闭基因的某一区段互补,能够抑制或封闭靶细胞基因的表达。
由于它与基因序列(称为正义链)碱基互补,或者说具有某种意义上的“镜象”关系,因此,这类寡核苷酸称为“反义核酸”。
人类基因组计划的进展为反义核酸技术的发展奠定了坚实基础。
近年来,国际著名的大型制药公司纷纷以各种方式介入反义技术研究,并且有多个反义核酸药物进入三期临床试验,这些都表明反义核酸技术及其产品的发展前景十分广阔。
反义核酸技术在药物研究方面的发展日臻成熟,随着基因组学的发展,在其他方面的应用也开始受到关注。
包括农业育种、功能基因研究、化妆品等方面都已经有了成功应用的例子。
反义技术是继基因克隆和重组技术之后在分子生物学领域中兴起的一门全新的基因工程技术。
采用反义技术开发新的生物医学美容产品,使生物医学美容从生理上完成人体的延缓衰老、抗皱、去痘、美白与健康,已经成为高科技化妆品研究的一个热点。
与疾病的产生原因相同,人体美的外在表现同样离不开基因。
基因通过其表达产物——蛋白质调节人体各种生理和生化过程,从而产生影响人体美的结果。
应用反义核酸技术筛选出表达影响人体美的蛋白质基因,设计一段与之互补的反义片段,将其封闭,从源头上抑制这种蛋白质的产生,借此达到美容效果。
目前,国外已有多个化妆品产品应用了反义技术,国内进行反义技术化妆品开发的企业还很少。
反义核酸技术系统应用于化妆品开发的特点设计简单——根据碱基互补原理,只要知道某种美损性皮肤问题的靶基因序列,就可设计相应的反义核酸DNA,进行系列化妆品产品的开发。
特异性高——反义核酸通常为2
0个碱基左右的寡核苷酸,它通过多点与靶基因结合,因而具有非常高的特异性。
靶点丰富——人类基因组计划研究成果为反义技术提供了丰富的靶点,可以不断开发出新的美容产品。
反义核酸技术化妆品的特点与传统化妆品相比,反义核酸技术化妆品突破了传统化妆品的界限。
安全性——反义核酸DNA与人皮肤亲和性和同源性高,对人体皮肤不产生毒、副作用。
参与皮肤细胞的生命活动——反义核酸化妆品直接参与人体皮肤细胞的生命活动,阻断致病mRNA翻译为致病蛋白质的过程,从本质上改善皮肤质量,达到美容目的。
促进健康基因生长,达到综合美容效果——明确了与美容相关的功能基因后,利用反义技术,对功能基因进行正向或负向调控,使皮肤细胞达到健康平衡的良好生理水平,达到综合美容效果。
(三)核酶在医学上的应用
1、核酶抗肝炎病毒的研究
目前人们已进行了核酶抗甲型肝炎病毒(HAV)、乙型肝炎病毒(HBV)、丙型肝炎病毒(HCV)以及HDV作用的研究。
人工设计核酶多为锤头状结构,少部分是采用发夹状核酶。
2、抗人类免疫缺陷病毒Ⅰ型(HIV-Ⅰ)核酶
1998年,美国加利福尼亚大学Wong-Staal等利用发夹核酶抑制HIV-Ⅰ基因表达,并在Ⅰ期临床实验中受到良好效果。
3、抗肿瘤治疗
核酶能在特定位点准确有效地识别和切割肿瘤细胞的mRNA,抑制肿瘤基因的表达,达到治疗肿瘤的目的。
五、核酶技术面临的问题
1、核酶催化切割反应的可逆性问题
2、催化效率低,如何提高催化效率
3、寻找合适载体将核酶高效、特异地导入靶细胞
4、使核酶在细胞内有调控地高效表达
5、增强核酶在细胞内的稳定性
6、对宿主的损伤问题有待进一步考察。