靶向哺乳动物细胞线粒体的核酸转运
mirna作用原理

mirna作用原理一、引言mirna(microRNA)是一类长度约为20~24个核苷酸的非编码RNA分子,是细胞内调控基因表达的重要因素。
在哺乳动物中,mirna可通过与mRNA靶标相互作用,调控基因转录、剪接和蛋白翻译等过程。
二、mirna的合成与成熟mirna的合成经历多个步骤,包括转录、成熟和靶标识别等过程。
具体步骤如下:2.1 转录mirna基因位于基因组的非编码区域,与mRNA的转录有所不同。
mirna的转录通常由RNA聚合酶II进行,但其转录起始位点可被辅助序列、转录因子和启动子等调控。
转录后的初始转录物被称为pri-mirna。
2.2 剪接和修饰pri-mirna在细胞核内由核酸酶Drosha切割生成前体mirna(pre-mirna)。
pre-mirna包含一个带有长的发夹结构,该结构可保护mirna序列免受核酸酶RNase的降解。
随后,pre-mirna被转运至胞质。
2.3 Dicer酶的介入在胞质中,pre-mirna被RNase III类酶Dicer切割,生成成熟的双链mirna (mature mirna)。
Dicer酶通过识别pre-mirna的发夹结构,切割出成熟mirna 序列。
2.4 修饰与装配成熟mirna的两个链的两端均会发生修饰,包括磷酸化和甲基化。
此外,成熟mirna还与Argonaute(AGO)蛋白相结合,形成RNA诱导沉默复合物(RISC),以实现靶向调控。
三、mirna的作用机制mirna通过与mRNA靶标相互作用,调控基因表达。
其作用机制主要包括激活RNase 活性和抑制翻译两种方式。
3.1 miRNA结合靶标mRNAmirna通过与mRNA的3’非翻译区(3’ UTR)相互作用,形成RNA复合物。
该相互作用主要通过mirna的5’末端6-7个碱基与mRNA的靶标区域互补配对。
3.2 激活RNase活性当mirna与mRNA结合时,mirna可以作为引导RNA,将RNA诱导沉默复合物(RISC)靠近目标RNA。
备战2024年高考生物易错题(新高考专用)易错点17 生物工程的“四个”不相等(含答案

备战2024年高考生物易错题(新高考专用)易错点17 生物工程的“四个”不相等(含答案易错点17 生物工程的“四个”不相等目录01 易错陷阱(四大陷阱)02 举一反三【易错点提醒一】脱毒苗≠抗毒苗【易错点提醒二】杂交瘤细胞≠分泌所需抗体【易错点提醒三】卵子受精≠受精完成【易错点提醒四】受体细胞有抗性≠导入重组DNA03 易错题通关易错陷阱1:利用植物组织培养技术培养植物分生区可获得抗毒苗【分析】植物组织培养技术培养植物分生区获得的是脱毒苗,没有抗病毒的能力不是抗毒苗(基因工程可得到)。
易错陷阱2:第一次筛选得到的杂交瘤细胞即可分泌所需的抗体【分析】经过克隆化培养和抗体检测筛选后得到的杂交瘤细胞才能分泌所需抗体。
易错陷阱3:卵子受精就代表受精作用完成【分析】形成两个极体代表卵子受精,常以观察到两个极体或者雌、雄原核作为受精的标志。
易错陷阱4:受体细胞表现抗性就表示导入重组DNA【分析】受体细胞表现抗性也有可能导入空载体。
【易错点提醒一】脱毒苗≠抗毒苗【例1】甘薯由于具有增强免疫功能、防癌抗癌、抗衰老、防止动脉硬化等作用,故越来越被人们喜爱。
但甘薯属于无性繁殖作物,同时又是同源六倍体,具有自交不育和杂交不亲和性,这使甘薯生产和育种存在诸多常规方法难于解决的问题。
下列相关操作不合理的是()A.用于快速繁殖优良品种的植物组织培养技术,可以解决种苗用量大、成本高的问题B.利用茎尖分生组织培养脱毒苗,使植株具备抗病毒的能力,产品质量得到提高C.利用原生质体融合实现体细胞杂交,克服杂交不亲和,可以充分利用遗传资源D.利用培养的愈伤组织进行诱变育种,可以显著提高变异频率,有利于缩短育种年限【变式1-1】下列实例培育过程中应用到植物组织培养技术的是()①制造人工种子②培育抗盐转基因植物③培育抗病毒转基因烟草④培育单倍体A.②③④B.①③④C.①②④D.①②③④【变式1-2】辣椒素作为一种生物碱广泛用于食品保健、医药工业等领域。
肠道微生物组与健康:机制见解

肠道微生物组与健康:机制见解摘要:肠道微生物群现在被认为是有助于调节宿主健康的关键元素之一。
几乎所有的身体部位都被微生物定植,这表明与我们的器官存在不同类型的串扰。
由于分子工具和技术(即宏基因组学、代谢组学、脂质组学、宏转录组学)的发展,宿主和不同微生物之间发生的复杂相互作用正在逐步被破译。
如今,肠道微生物群偏差与许多疾病有关,包括肥胖、2 型糖尿病、肝脂肪变性、肠病(IBD)和几种类型的癌症。
因此,表明涉及免疫、能量、脂质和葡萄糖代谢的各种途径受到影响。
在这篇综述中,特别关注对该领域当前理解的批判性评估。
讨论了许多解释肠道细菌如何与保护或疾病发作有因果关系的分子机制。
我们检查了公认的代谢物(即短链脂肪酸、胆汁酸、三甲胺 N-氧化物),并将其扩展到最近确定的分子作用物(即内源性大麻素、生物活性脂质、酚衍生化合物、晚期糖基化终产物和肠联基因)及其特异性受体,如过氧化物酶体增殖物激活受体α (PPARα)和γ (PPARγ)、芳烃受体(AhR)和 G 蛋白偶联受体(即 GPR41、GPR43、GPR119、武田 G 蛋白偶联受体 5)。
总而言之,了解将肠道微生物与健康联系起来的复杂性和分子方面将有助于为已经开发的新疗法奠定基础。
人类肠道微生物组人类微生物组在这里被认为是微生物、它们的基因和产物的集合,它们从出生起就在我们体内定植并垂直转移。
虽然所有身体部位都被定植(图 1),但在肠道中发现的微生物数量最高,这已经得到了广泛的研究。
在这里,我们回顾了解决肠道微生物、其活性和介质分子如何促进我们健康的主要和最新发现。
图 1 根据不同身体部位的细菌总丰度。
不同器官中细菌数的边界,由细菌浓度和体积得出。
在健康受试者中,口腔和唾液微生物组包含数百万种微生物,这些微生物每天与我们的食物一起吞咽,但它们在肠道中的持久性受到许多因素的阻碍,包括胃的酸度、十二指肠内外胆汁酸(BA)的产生、消化酶和抗菌蛋白许多其他主要变量会影响进一步的下游微生物定植,例如 pH 值、氧浓度和氧化还原电位等化学参数、粘液、胆汁和抗体的生物产生,以及物理方面,包括肠道结构、蠕动和转运时间(图 1)。
靶向Bcl-2、Bcl-xl、Mcl-1、Bcl-w、A1反义核酸抑制消化系肿瘤细胞增殖效果的差异

靶向Bcl-2、Bcl-xl、Mcl-1、Bcl-w、A1反义核酸抑制消化系肿瘤细胞增殖效果的差异蒋建伟;吴风云;何金花;廖晓莉;王威;吴志慧【摘要】目的比较靶向Bcl-2、Bcl-xl、Mcl-1、Bcl-w、A1反义核酸(antisense oligodeoxynucleotide,ASO)对消化系统肿瘤细胞(肝癌HepG2细胞、胃癌MGC-803细胞、结肠癌Lovo细胞)的增殖抑制作用和致凋亡作用的差异.方法分别合成靶向Bcl-2、Bcl-xl、Mcl-1、Bcl-w、A1的反义核酸和随机序列反义核酸(random oligodeoxynucleotide,RODN),采用脂质体LipofectamineTM2000转染细胞,WST法检测相同浓度的5种ASOs对肝癌HepG2细胞、胃癌MGC-803细胞、结肠癌Lovo细胞的增殖抑制作用和致凋亡作用.结果 Bcl-2、Bcl-xl、Mcl-1、Bcl-w、A1等5种ASOs中,不论是对细胞增殖抑制还是致凋亡作用,均以Bcl-xl ASO、Mcl-1 ASO的作用效果较好.结论在Bcl-2、Bcl-xl、Mcl-1、Bcl-w、A1等5种ASOs中,Bcl-xl ASO、Mcl-1 ASO可明显抑制消化系肿瘤细胞增殖,诱导细胞凋亡.【期刊名称】《中国药理学通报》【年(卷),期】2010(026)008【总页数】6页(P1093-1098)【关键词】反义核酸;Bcl-2;Bcl-xl;Mcl-1;增殖;凋亡;肿瘤细胞【作者】蒋建伟;吴风云;何金花;廖晓莉;王威;吴志慧【作者单位】暨南大学医学院生化教研室,广东,广州,510630;暨南大学医学院生化教研室,广东,广州,510630;浙江省温州市平阳县人民医院,浙江,温州,325400;暨南大学医学院生化教研室,广东,广州,510630;广东省广州市番禺区中心医院检验科,广东,广州,511440;暨南大学医学院生化教研室,广东,广州,510630;暨南大学医学院生化教研室,广东,广州,510630;暨南大学医学院生化教研室,广东,广州,510630【正文语种】中文【中图分类】R329.24;R329.25;R735.022线粒体相关的细胞凋亡途径通过Bcl-2家族蛋白的调节。
细胞内蛋白质定位和转运机制

细胞内蛋白质定位和转运机制细胞内蛋白质定位和转运机制是细胞内重要的生物学过程,它们维持了细胞的正常功能并参与了各种生物学活动。
本文将从细胞内蛋白质定位的基本原理、信号序列和定位机制以及蛋白质的转运机制等方面进行探讨。
一、细胞内蛋白质定位的基本原理细胞内蛋白质定位是指将蛋白质定向到细胞内特定的亚细胞结构或位置。
这一过程是通过特定的信号序列和机制实现的。
蛋白质定位的基本原理可以概括为两大类:靶向和扩散。
靶向是指蛋白质在合成过程中通过与一些特定的蛋白质或结构发生相互作用,从而被定向到细胞内的特定位置。
例如,细胞内的Golgi体是一个重要的分泌细胞器,某些蛋白质通过与Golgi体中的转运蛋白相互作用,从而被定位到Golgi体。
扩散是指蛋白质在合成过程中通过不断的扩散和分布,最终到达细胞内的特定位置。
这种定位机制主要依赖于蛋白质的物理和化学性质,以及细胞内各种蛋白质相互作用的平衡。
例如,细胞内水溶性蛋白质通过扩散和分布到达到达核内。
二、信号序列和定位机制在蛋白质的定位过程中,信号序列起到了非常重要的作用。
信号序列是蛋白质分子上的某一特定的氨基酸序列,它能够指导蛋白质被定位到特定的亚细胞结构或位置。
信号序列可以分为靶向信号序列和细胞内定位信号序列。
靶向信号序列通常位于蛋白质分子起始处,它能够与特定的蛋白质或结构发生相互作用,从而将蛋白质定向到细胞内的某个结构或位置。
细胞内定位信号序列通常位于蛋白质的内部,它能够改变蛋白质的物理和化学性质,从而影响蛋白质的定位。
蛋白质的定位机制可以分为几种类型:核定位、细胞质定位、内质网定位、线粒体定位、高尔基体定位等。
不同类型的定位机制通常与不同的信号序列和作用蛋白有关。
例如,核定位的信号序列通常富含正电荷氨基酸,而线粒体定位的信号序列则富含氨基酸序列(R-X-X-R)。
三、蛋白质的转运机制蛋白质定位到细胞内的特定位置后,往往需要通过转运机制到达目标位置。
蛋白质的转运可以分为受体介导转运和核孔复合物介导转运两种方式。
PHB2(Prohibitin 2)在线粒体中的研究进展

PHB2(Prohibitin 2)在线粒体中的研究进展董艳博;冯红【摘要】PHB2是真核细胞的一种在进化过程中高度保守的蛋白质,参与细胞的多种生命活动.有研究表明PHB2参与细胞周期的调节、转录调节、细胞增殖和凋亡、线粒体嵴形态的发生、信号转导等多种细胞过程.它广泛表达,分布于酵母、植物、蠕虫、苍蝇、哺乳动物等物种中,并且PHB2蛋白质是以复合物的形式定位于线粒体上.PHB2主要作为线粒体内膜的自噬受体,参与靶向线粒体的自噬和降解.研究综合了国内外的文献,对PHB2的功能,结构及表达进行阐述,为进一步研究PHB2蛋白质的功能提供了基础.【期刊名称】《天津科技》【年(卷),期】2018(045)001【总页数】4页(P36-39)【关键词】PHB2;线粒体;功能结构【作者】董艳博;冯红【作者单位】天津体育学院健康与运动科学系天津300381;天津体育学院健康与运动科学系天津300381【正文语种】中文【中图分类】Q4130 引言PHB首先作为细胞增殖的抑制剂而被发现,它的结构相当保守且普遍存在[1]。
其广泛分布于哺乳动物、原虫、植物、细菌以及真菌等生物体中。
人类基因组编码两种 PHB蛋白质,即 prohibitin 1(PHB1)和prohibitin 2(PHB2),phb1基因位于染色体17q21上,phb2基因位于染色体 12p13上[2]。
有研究表明,PHB1和PHB2的复合物是维持线粒体的功能所必需的,该复合物组成了一个环状大分子结构,在不同的细胞过程(如细胞周期进程和衰老)以及许多疾病(如肥胖,糖尿病和癌症等)中都发挥着重要作用(见图1)[3]。
1 PHB2的结构PHB2最初是由 Terashima及其同事们发现的,因与大鼠 B淋巴细胞的 IgM 受体(Immunoglobulin M receptor,免疫球蛋白质受体)相互作用而被命名为BAP37[4]。
PHB2蛋白质的氨基端有一段疏水区域,该区域将PHB2固定在膜上;羧基端含有一个保守的PHB结构域和一个可预测的螺旋区域,在酵母细胞中该区域与 PHB蛋白质复合物的合成密切相关[5]。
《细胞生物学》——细胞6章 蛋白质分选与膜泡运输雨课堂10-11
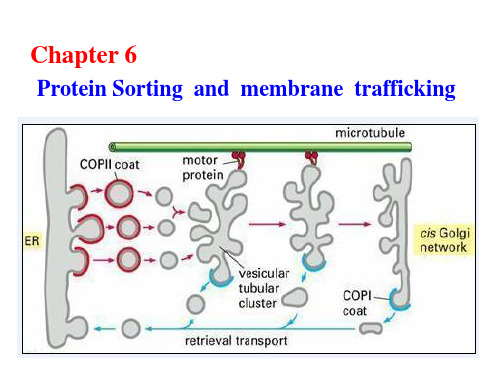
(一)蛋白质从细胞质基质输入到线粒体
1.线粒体蛋白从细胞质基质输入到线粒体基质: 两性的N端靶向信号序列(形成α螺旋构象); 分子伴侣胞质蛋白Hsc70和线粒体基质蛋白Hsc70; 从内外膜接触点的Tom(外膜移位子)和Tim(内膜移 位子)处输入。
2.线粒体蛋白以3种途径从细胞质基质输入到线粒体内膜:
转运到其功能发挥部位的过程。 蛋白质分选不仅保证了蛋白质的正确定位,也保证
了蛋白质的生物学活性
一、信号假说与蛋白质分选信号 二、蛋白质分选转运的基本途径与类型
三、蛋白质向线粒体、叶绿体和过氧化酶体 的分选
一、信号假说与蛋白质分选信号
20世纪60年代,乔治•帕拉德(George Palade)等发现,细胞 分泌的蛋白需要先进入内质网,再到高尔基体,然后分泌到 胞外。这个细胞分泌途径的重大发现,使他获得了1974年诺 贝尔生理学或医学奖
• 多次跨膜蛋白:含有多个SA和多个STA的肽链将成为多次跨膜 蛋白。
• 跨内质网膜肽段的取向: 。
内质网膜整合蛋白的拓扑学类型
• STA:内部停止转移锚定序列 SA:内部信号锚定序列
• 线粒体、叶绿体和过氧化物酶体的蛋白质的信号序列 特称为导肽(leader peptide),其基本的特征是蛋白 质在细胞质基质中的游离核糖体上合成以后再转移到 这些细胞器中,因此称这种翻译后再转运的方式为后 翻译转运(post-translational translocation)。
在游离核糖体上起始合成由信号肽和与之结合的 SRP引导转移至糙面内质网 新生肽边合成边转运 糙面内质网腔或定位在ER膜上 经转运膜泡运 到高尔基体加工包装 分选至溶酶体、细胞质膜 或分泌到细胞外。 (2)后翻译转运(post-translational translocation)途径 在细胞质基质游离核糖体上合成以后 细胞核、 线粒体、叶绿体和过氧化物酶体,或者成为细胞质 基质的可溶性驻留蛋白和骨架蛋白。
非编码RNA的研究与应用

非编码RNA的研究与应用在近几十年的生命科学研究中,DNA和蛋白质一直是研究的重点,因为它们是生命体内最重要的两种分子。
但是,在过去的几十年中,随着RNA研究的深入,人们逐渐发现了RNA在生命过程中的重要性。
RNA通过转录和转录后修饰的方式,在细胞的生命活动中扮演着重要的角色。
近些年来,非编码RNA在RNA家族中引起了越来越多的关注。
非编码RNA的定义非编码RNA是指不参与编码蛋白质的RNA。
它们通常具有重要的生物学功能,包括转录调控、RNA剪切调控、蛋白质翻译调控以及RNA沉默等。
尽管非编码RNA在生命活动中扮演着重要的角色,但是长期以来它们一直被忽视或被认为只是某些细胞过程中附带存在的“垃圾”分子。
种类和功能非编码RNA通常可以分为两类:小的非编码RNA和长的非编码RNA。
小的非编码RNA的长度通常在20-30个核苷酸之间,包括RNA干扰RNA(siRNA)、微小RNA(miRNA)、piwi相关RNA(piRNA)和转运RNA(tRNA)等。
长的非编码RNA的长度通常超过200个核苷酸,包括长链非编码RNA(lncRNA)、circRNA(循环RNA)和snRNA(小核RNA)等。
siRNA是由双链RNA剪切后形成的小分子,它们主要通过靶向mRNA的特定区域,通过RNA序列匹配的方式降低转录和翻译活动。
miRNA是由小的非编码RNA构成的,它们在靶向mRNA以降低转录和翻译活动方面也发挥着重要的作用。
piRNA 主要存在于生殖细胞中,它们参与着细胞分裂和DNA修复等生命过程。
tRNA参与了蛋白质合成的过程,是细胞中蛋白质翻译的重要组成成分。
lncRNA是长链非编码RNA,它们在转录和表观遗传调控方面发挥着重要的作用。
circRNA是一类特殊的长链非编码RNA,它们通过形成循环结构维护其稳定性。
snRNA是小核RNA,在基因剪接、核糖体RNA组装以及mRNA剪切等生物过程中发挥重要作用。
非编码RNA的研究进展近年来,随着生物学研究的不断深入,越来越多的关于非编码RNA的研究也相继浮出水面。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
靶向哺乳动物细胞线粒体的核酸转运付爱玲【摘要】Mitochondrial DNA (mtDNA)genome mutations and defects are the essential mechanism of a various of mitochondrial dysfunction associated with diseases. The studies of targeting de-livery nucleic acid into mammalian mitochondria can thoroughly correct mtDNA mutation, rescue mtDNA impairment and then reverse the progress of diseases. There’s obvious differences be-tween nucleic acid import pathway of mammalian mitochondria and gene transfection of nuclei. In this paper, the effective strat-egies of delivering DNA and RNA(tRNA,rRNA,mRNA and an-tisense RNA)into mitochondria have been reviewed, as well as the challenges and development.%线粒体 DNA(mitochondrial DNA, mtDNA)的遗传性突变和缺陷是多种线粒体功能失调相关疾病的根本原因。
靶向线粒体递送核酸,可从根本上纠正 mtDNA 突变、挽救 mtD-NA 损伤、阻断疾病进程。
哺乳细胞内线粒体的核酸转运途径与细胞核的基因转染大不相同。
该文综述了向哺乳动物细胞线粒体递送 DNA 和 RNA(tRNA、rRNA、mRNA 和反义RNA)的有效策略,并对其存在问题和发展趋势做一阐述。
【期刊名称】《中国药理学通报》【年(卷),期】2016(000)001【总页数】4页(P1-4)【关键词】线粒体转导;靶向序列;核酸转运;线粒体相关疾病;tRNA 载体;5s rRNA转导;穿梭蛋白【作者】付爱玲【作者单位】西南大学药学院,重庆 400716【正文语种】中文【中图分类】R-05;R329.24;R342.2;R342.3;R589.9;R977.6中国图书分类号: R-05; R329. 24; R342. 2; R342. 3; R589. 9; R977. 6线粒体是哺乳动物中唯一具有核外DNA的细胞器,它保留着自身完整的一套遗传系统,为氧化磷酸化相关复合物的生成提供必需的多肽和蛋白质。
哺乳动物线粒体DNA ( mitochondrial DNA,mtDNA)长度为16.5 kbp,共编码13个与氧化磷酸化有关的多肽和蛋白质、22个tRNA和2个rRNA,不包含基因间隔区和内含子[1]。
哺乳动物mtDNA突变显然对宿主细胞有明显的不良影响,并且大量的事实已证明线粒体基因组突变是引发多种疾病的主要原因[2]:在人类mtDNA中,已确定超过300个突变与疾病的发生有关( http: / /www.mitomap.org),如神经和肌肉退行性病变、心肌疾病、肿瘤、糖尿病和病理性衰老等[3]。
常见的突变包括编码蛋白的基因反义突变、tRNA和rRNA基因点突变、基因复制或删除等等[4]。
一般认为,当野生型和突变的mtDNA的比例达到一定的阈值时,就会出现临床症状[5]。
靶向线粒体基因组的核酸转运是治疗线粒体疾病的一个有效方法。
线粒体进行DNA转染的主要困难是它的双层膜、体积小、数量大。
为避开线粒体转染的难题,人们曾尝试间接蛋白质转导这一替代方法:通过质粒或病毒转染细胞,使细胞核表达由于mtDNA致病突变而缺乏的蛋白质,再经线粒体靶向序列将表达的蛋白质运输到线粒体[6]。
尽管这一间接方法可能会得到令人鼓舞的结果,向线粒体递送核酸可从根本上纠正mtDNA突变、挽救mtDNA损伤、阻断疾病进程。
已建立的方法主要有线粒体转导、载体介导、靶向肽或靶向序列引导、穿梭蛋白介导核酸进入细胞线粒体;此外,将外源RNA插入rRNA或tRNA中,根据线粒体对胞质内RNA的摄取作用,可将外源的RNA转运至线粒体内( Fig 1)。
向线粒体内转运核酸仍处于当今研究的前沿。
1.1整个线粒体转导在正常状态下,哺乳动物和植物细胞的线粒体极少从胞质中直接摄取DNA,因此常规的细胞转染方法不能使DNA进入线粒体。
而单细胞生物酵母和莱茵衣藻不同,它们的线粒体可从胞质中摄取少量的DNA。
在早期的研究中,Clark等[7]将分离的线粒体( mtDNA中含有抗生素耐受性序列)与哺乳细胞共孵育,得到了抗生素耐受性的细胞,表明分离纯化的线粒体可直接被细胞内吞,并在胞质中发挥作用,从而使原本对抗生素敏感的细胞对抗生素产生了耐受性。
然而,在以后的25年内,一直没有人关注这个发现。
直至Katrangi 等[8]报道了将分离的正常线粒体与去除线粒体的人A549肺癌细胞一起孵育,正常线粒体被细胞所摄取,从而恢复了细胞的呼吸链。
在最近的报道中,为提高线粒体进入细胞的效率,将细胞穿透肽Pep-1连接到分离的野生型线粒体(活力约为83. 5%)表面,加入到肌阵挛性癫痫伴蓬毛样红纤维综合症病人的细胞中,结果显示在给予线粒体3 d后,细胞的线粒体功能得到了恢复,包括恢复了氧化磷酸化亚单位(复合物Ⅰ、Ⅲ和Ⅳ)的活性、线粒体膜电位和ATP的生成量,同时降低了活性氧自由基的产生[9]。
此外,将外源线粒体显微注射到细胞内,可导致外源线粒体快速取代细胞自身的线粒体。
这个方法可用于改善受精卵线粒体功能的研究。
由于哺乳动物的受精卵线粒体来源于母系,母系线粒体中的遗传突变将传递给下一代,因此线粒体替代的方法可能会对母系遗传疾病有一定的治疗作用。
研究表明,将分离的线粒体显微注射到小鼠受精卵中,在胚胎发育的第一阶段可检测到外源线粒体和mtDNA[10-11],然而,随后其水平大幅下降,其机制尚不清楚。
1.2重组DNA的线粒体转导在验证外源线粒体替代的同时,研究还发现了线性DNA可直接进入分离的线粒体。
在缺乏任何人工手段下,线状的裸DNA可主动进入分离的植物、动物和真菌线粒体中[12]。
此外,外源DNA进入线粒体后,可进行转录和表达,并且一些损伤的DNA在进入植物和动物线粒体后也可被修复。
已知细胞核基因组在细胞DNA转染时,对DNA的序列和长度有所限制,但分离的人线粒体对线性DNA的长度和序列无选择性。
环状DNA(质粒)可通过电穿孔的方式进入线粒体。
在生理条件下,电穿孔可引发哺乳动物、原生生物或植物中分离的线粒体对DNA的摄取。
Yoon等[13]将来源于大肠杆菌的DNA中增加了一个转导起始区( origin of transfer,oriT)序列,然后将其转染入分离的线粒体,当把线粒体与活细胞孵育后,重组的DNA可在活细胞的线粒体内进行表达。
此外,有人在无复制性的大肠杆菌中构建哺乳动物的全部mtDNA,将这种含有mtDNA的大肠杆菌介导至体外培养的哺乳动物细胞质中,使克隆的mtDNA与细胞本身的线粒体mtDNA对接。
这种由大肠杆菌介导的mtDNA重组的方法也能得到设计的mtDNA[14]。
1.3线粒体靶向序列引导DNA进入向线粒体与整个线粒体转导的研究相反,通过线粒体靶向序列或靶向肽将DNA转运进入线粒体,这些研究开展较早、进展较快,并仍在持续进行。
已知由细胞核指导的线粒体前体蛋白在合成后,在N末端带有线粒体靶向序列。
该序列通过线粒体的蛋白导入途径,引导蛋白质进入线粒体。
常用的线粒体靶向序列有细胞色素氧化酶亚基8前体蛋白N末端的23个氨基酸( MSVLTPLLLRGLTGSARRLPVPR)等[15]。
线粒体靶向序列也可引导其他分子(核酸、纳米颗粒、脂质体)等进入线粒体。
将线粒体靶向肽的N末端与17个或322个碱基对的DNA回文序列连接,连接物也可进入分离的大鼠肝线粒体中。
在另外的体系中,线粒体靶向肽与肽核酸的一个片段连接,复合物可进入分离的哺乳动物细胞线粒体中。
近期研究表明,线粒体靶向肽与结合DNA的PEI偶联,多肽/PEI/ DNA复合物可定位在活细胞的线粒体[16]。
然而,该方法存在的问题是DNA与多肽的连接物不易合成,且易于水解,此外,线粒体靶向序列引导其他物质虽然可进入线粒体,但可能难以通透细胞膜,因此,需其他有效的途径弥补这个方法的缺陷。
为解决线粒体靶向序列与DNA的连接物可能难以进入细胞的问题,进一步开发了“转导载体”复合物。
该复合物包含了细胞穿透肽[17-18]、线粒体靶向肽和线粒体转录因子A( TFAM)或mtDNA结合因子。
其中后者可与DNA非共价结合,细胞穿透肽引导复合物进入细胞。
而线粒体靶向肽引导复合物进入线粒体。
这个复合物曾用于向帕金森病模型细胞的线粒体运送DNA,结果表明转导载体和DNA的共定位于线粒体,并阻断了线粒体损伤[19]。
此外,使用转导载体将正常的mtDNA运输进含有突变mtDNA的人类细胞,线粒体的功能可部分恢复。
如果通过转导载体将致病的mtDNA介导入人神经祖细胞,外源致病的mtDNA也可在线粒体中表达。
1.4纳米载体介导DNA进入线粒体设计的非阳离子脂质体载体MITO-porter能够用于向线粒体输送DNA[20]。
MITO-porter是一个内核含有运输的药物,外层分别为线粒体融合层和内吞体融合层的纳米颗粒。
MITO-porter经内吞进入细胞后,外层分别通过与内吞体膜和线粒体膜融合,以膜融合的方式携带药物进入活细胞线粒体基质中[21]。
有些双性载体也能够通过线粒体膜并聚集在线粒体内(如三苯基膦),这些载体可用于向线粒体内转运DNA。
双性载体可通透脂质双层膜进入线粒体中。
将PNA寡聚体与之共价连接后,在膜电势驱动下,双性载体将PNA携带进入培养的人细胞的线粒体。
此外,丝氨酸衍生的双性表面活性剂/DNA复合物也可经内吞机制进入细胞后,转染50%以上细胞的线粒体[22]。
尽管正常细胞的线粒体很少从胞质中摄取DNA,但在大多数物种中,线粒体可天然地从胞质中摄取RNAs,其中最常见的是tRNA。
这是由于线粒体的基因组并不编码完整的一套tRNA,因此线粒体就需分享胞质翻译系统中核编码的tRNAs。
不同物种能够转导进线粒体的tRNAs数量不同,例如有袋动物的线粒体仅可转导进一个tRNAs,而锥虫线粒体可转导进所有tRNAs[23]。
tRNAs进入线粒体的机制极可能是非特异tRNA导入复合体( tRNA import complex,RIC)途径。