哺乳动物细胞表达系统
cho细胞表达重组蛋白方案

CHO (Chinese Hamster Ovary) 细胞是常用的哺乳动物细胞系统,用于表达重组蛋白的研究和生产。
以下是一般性的CHO 细胞表达重组蛋白的方案:
1. 购买表达载体:选择适合的表达载体,可以是质粒或病毒载体。
载体应包含适当的启动子、选择标记等。
2. 转染CHO 细胞:将表达载体导入CHO 细胞中。
转染方法可以选择经典的化学或电穿孔法,也可以选择使用特定的转染试剂或转染仪器。
3. 选择稳定转染株:在转染后,使用适当的选择剂(如抗生素) 处理细胞,以选择稳定表达重组蛋白的细胞株。
可通过单克隆分离等方法筛选和扩增单一细胞克隆。
4. 细胞培养条件优化:优化培养基配方和细胞培养条件,包括温度、pH 值、培养基组分等,以提高重组蛋白的产量和纯度。
5. 表达蛋白的诱导:使用适当的诱导剂或方法,例如添加诱导剂(如甲酪酸) 到培养基中,以启动重组蛋白的表达。
6. 重组蛋白的纯化和分析:通过细胞破碎和不同的纯化步骤(如亲和层析、离子交换层析、凝胶过滤等)从培养基或细胞提取物中纯化目标重组蛋白,并使用适当的分析方法验证表达的蛋白的纯度和功能。
在每个步骤中,需要根据具体的重组蛋白和研究目的进行优化和调整。
此外,合理的培养细胞和操作操作也至关重要,以确保产量和纯度的理想达到。
这些方案的细节将根据具体的实验目的和需要进行个体化定制。
蛋白体外表达与纯化

蛋白体外表达与纯化随着后基因组时代的到来,蛋白质组成为科学研究的热点。
蛋白质作为生命机体的主要活动的承担者,其体外表达与纯化在研究相应基因的功能上有重要意义。
蛋白体外表达系统按其表达宿主可分为原核表达系统,真核表达系统和哺乳动物细胞表达系统。
一:原核表达系统原核表达系统的宿主菌主要以大肠杆菌为代表,大肠杆菌表达体系是目前应用最广泛的外源基因表达体系,这也是外源基因表达的首选体系。
该表达体系的优点:遗传学和生理学背景清楚;容易培养;外源基因经常可以高效表达及操作简单、周期短、成本低等。
其不足之处是不能进行典型真核细胞所具有的复杂的翻译后修饰;广泛的二硫键的形成及外源蛋白组装成蛋白复合体的能力也受到限制;另外外源基因产物在大肠杆菌中易形成不溶的包涵体;有时由于真核mRNA的结构特性及密码子使用频率与大肠杆菌的差异,而的不到足够的产物。
二:真核表达系统真核表达系统的宿主菌主要以酵母表达系统为代表,酵母基因表达系统的载体通常既能在酵母中进行复制也能在大肠杆菌中进行复制,形成所谓酵母菌――大肠杆菌穿梭载体。
因以大肠制备质粒DNA较方便,通常利用大肠杆菌系统构建酵母载体以简化手续,缩短时间。
作为基因表达系统的宿主应该具备以下条件:安全无毒,不致病;遗传背景较清楚,容易进行遗传操作;容易进行载体DNA的导入;培养条件简单;有良好的蛋白分泌能力;有类似高等真核生物的蛋白翻译后修饰功能。
三:哺乳动物细胞表达系统由于本专业不涉及哺乳动物细胞表达系统的应用,故此不赘述。
表达载体的种类及相应的分离纯化方法作为表达载体必须具备以下特征:稳定的遗传复制、传代能力,无选择压力下能存在于宿主细胞内;具有显性的筛选标记;启动子的转录是可调控的;启动子的转录的mRNA能够在适当的位置终止;具有外源基因插入的多克隆位点。
在原核表达系统中常用的表达载体有:PET-载体系列,用这类载体表达出的外源蛋白在N端或C端或两端均具有his tag。
哺乳动物表达系统原理

哺乳动物表达系统原理引言:哺乳动物表达系统是指哺乳动物通过语言、声音、姿势和化学信号等方式进行交流和传递信息的机制。
这种表达系统在哺乳动物种群中起着至关重要的作用,它有助于个体之间的合作、繁殖和求偶行为的进行。
本文将介绍哺乳动物表达系统的原理及其在不同物种中的表现。
一、语言交流:语言是人类最为重要的交流方式之一,但在其他哺乳动物中并不普遍存在。
然而,一些哺乳动物如海豚、鲸鱼和某些灵长类动物,通过发出特定的声音来传递信息。
这些声音可以包括呼吸声、鸣叫声、鸣叫声等。
它们通过调整声音的频率、音量和时长来传递不同的意义,从而实现交流。
这种语言交流对于维持个体之间的社交关系、警戒和求偶行为至关重要。
二、声音表达:除了语言交流外,声音也是哺乳动物表达系统中的重要组成部分。
许多哺乳动物通过发出特定的声音来传达不同的信息。
例如,大象通过发出嘶吼声、波动声和低音鼻音来进行交流。
这些声音可以传达警告、求偶和社交的信息。
类似地,狗通过吠叫声来表达不同的心情和需求,如警戒、寻求注意和表达快乐。
三、姿势和动作:除了声音外,姿势和动作也是哺乳动物表达系统中的重要组成部分。
例如,狗通过摇尾巴、耳朵的姿势和身体的动作来表达不同的情绪和意图。
摇尾巴可以表示高兴、友好或兴奋,而低垂的耳朵和身体的低姿态则可能表示害怕或不安。
类似地,猫通过抓、踩和搔抓等动作来表达不同的意思,如攻击、抚摸和警告。
四、化学信号:除了声音、姿势和动作,化学信号也是哺乳动物表达系统中的重要组成部分。
许多哺乳动物通过释放具有特定化学成分的信息素来传递信息。
例如,狗通过尿液中的信息素来标记自己的领地和状态,以及吸引异性。
类似地,许多灵长类动物通过分泌具有特定化学成分的汗液来进行社交和求偶行为。
五、不同物种中的表现:不同的哺乳动物在表达系统中的表现也存在差异。
例如,灵长类动物如黑猩猩和猴子可以通过复杂的声音、姿势和面部表情来进行交流。
而大象则以低音的鸣叫声和身体姿势来进行交流。
哺乳动物细胞表达系统
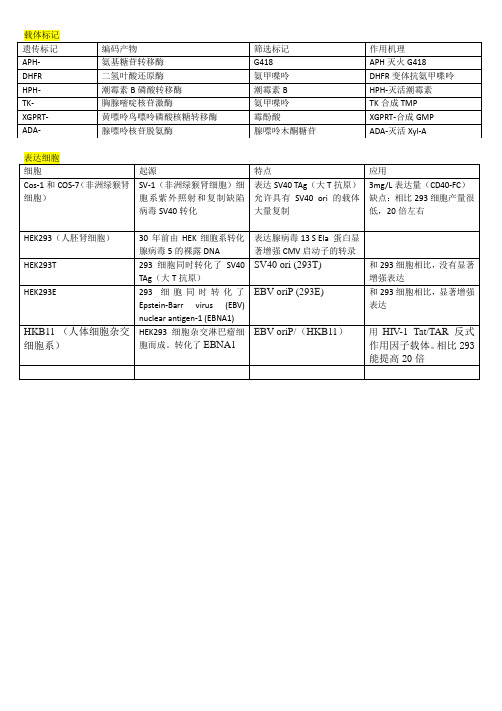
30年前由HEK细胞系转化腺病毒5的裸露DNA
表达腺病毒13 S Ela 蛋白显著增强CMV启动子的转录
HEK293T
293细胞同时转化了SV40 TAg(大T抗原)
SV40 ori (293T)
和293细胞相比,没有显著增强表达
HEK293E
293细胞同时转化了Epstein-Barr virus (EBV) nuclear antigen-1 (EBNA1)
EBV oriP(293E)
和293细胞相比,显著增强表达
HKB11(人体细胞杂交细胞系)
HEK293细胞杂交淋巴瘤细胞而成。转化了EBNA1
EBV oriP/(HKB11)
用HIV-1 Tat/TAR反式作用因子载体。相比293能提高20倍
遗传标记
编码产物
筛选标记
作用机理
APH-
氨基糖苷转移酶
G418
APH灭火G418
DHFR
二氢叶酸还原酶
氨甲喋呤
DHFR变体抗氨甲喋呤
HPH-
潮霉素B磷酸转移酶
潮霉素B
HPH-灭活潮霉素
TK-
胸腺嘧啶核苷激酶
氨甲喋呤
TK合成TMP
XGPRT-
黄嘌呤鸟嘌呤磷酸核糖转移酶
霉酚酸
XGPRT-合成GMP
ADA-
腺嘌呤核苷脱氨酶
腺嘌呤木酮糖苷
ADA-灭活Xyl-A
载体标记
表达细胞
细胞
Hale Waihona Puke 起源特点应用Cos-1和COS-7(非洲绿猴肾细胞)
SV-1(非洲绿猴肾细胞)细胞系紫外照射和复制缺陷病毒SV40转化
表达SV40 TAg(大T抗原)允许具有SV40 ori 的载体大量复制
哺乳动物细胞表达系统原理

哺乳动物细胞表达系统原理
哺乳动物细胞表达系统是一种用于生产重组蛋白和基因治疗的有效工具。
其原理主要基于哺乳动物细胞具有促使蛋白正确折叠和实现复杂修饰的功能,使得表达的蛋白更接近天然状态。
哺乳动物细胞表达系统有两种主要方式:瞬时转染表达和稳定转染表达(稳定细胞系构建)。
瞬时转染表达是指在短时间内表达出一定量的蛋白。
外源基因不整合到宿主染色体上,随着细胞的生长不断丢失,表达小量蛋白的过程。
这种方法简捷,实验周期短。
稳定转染/稳定细胞株筛选则是指将构建好的质粒线性化,在导入到培养好
的哺乳动物细胞内,通过一定的转染方法实现质粒与细胞的融合。
线性化的质粒进入到宿主细胞后,与细胞自身的基因组进行整合,同时随着细胞的生长繁殖而生长,过程中在经过一系列的筛选鉴定,排除未正确融合的重组子,或不能稳定表达下去的重组子,最终筛选出可以稳定表达的细胞株。
以上内容仅供参考,建议查阅哺乳动物细胞表达系统相关书籍获取更全面和准确的信息。
哺乳动物细胞表达系统

据文献报道,在不断提高的选择压力下,dhfr及侧翼序列 能扩增至上千个拷贝,大大增加目的基因的表达水平。
真核表达载体-启动子
外源基因在哺乳动物细胞中的表达与多种因素有关,主要 是启动子和增强子的强弱以及它们之间的搭配。
常用的高等哺乳动物受体细胞
迄今为止,用于医疗用品(药物、抗体、诊断试剂)大规 模生产的高等哺乳动物受体细胞主要还是中国仓鼠卵巢细 胞(CHO),其优势有如下几个方面: 遗传背景清楚,生理代谢稳定 与人的亲缘关系接近,外源蛋白修饰准确 基因转移和载体表达系统完善 耐受剪切力,便于大规模培养 被美国FDA确认为安全的基因工程受体细胞
Pei等用该细胞株表达分泌型的基质金属蛋白酶MMPI3,发现 高表达的阳性细胞克隆可占转染细胞的5%~l0%,其中一个 克隆的表达量可占细胞上清总蛋白的l5%~20%,在细胞单层 贴壁培养情况下表达量达10 mg/L。
对细胞株选择性地进行遗传改造
BHK/vl6细胞株是稳定表达单纯疱疹病毒(HSV)VP16蛋白 的BHK细胞,由于VP16的转录激活作用,载体中的HSV 早期启动子在该工程细胞中有很高的活性。
Clontech公司开发的Tet-off系统中的启动子则由CMV启动 子的核心序列和7个Tet阻遏蛋白结合位点组成。
这些启动子在诱导前后活性可相差4个数量级。
真核表达载体-启动子
在哺乳动物细胞中已发现存在大量在低氧环境中可诱导转录 的基因,如编码红细胞生成素(EPO)、转铁蛋白、血红素加 氧酶-1等的基因,它们都有一个共同的顺式作用元件 (CGTG ),有利于在5’或3’侧翼区的低氧诱导作用因子1(HIF-1)和低氧反应增强子(HRE)结合,激活靶基因的转录, 在低氧浓度下可使重组蛋白大量表达。
哺乳动物细胞表达系统的特点

哺乳动物细胞表达系统的特点
哺乳动物细胞表达系统是一种常用的重组蛋白表达系统,具有以下特点:
1. 表达的蛋白具有正确的翻译后修饰:哺乳动物细胞能够对表达的蛋白进行正确的翻译后修饰,如糖基化、磷酸化、乙酰化等,使表达的蛋白更接近天然蛋白的结构和功能。
2. 蛋白表达量较高:相对于其他表达系统,哺乳动物细胞表达系统能够产生较高水平的重组蛋白。
3. 适用于分泌型蛋白的表达:哺乳动物细胞具有完善的内质网和高尔基体等细胞器,可以将表达的蛋白分泌到细胞外,适用于分泌型蛋白的表达。
4. 产物易于纯化:哺乳动物细胞表达的重组蛋白通常具有较高的纯度,因为它们可以被分泌到细胞外,从而简化了纯化过程。
5. 适合治疗性蛋白的生产:由于哺乳动物细胞表达的蛋白具有与人体自身蛋白相似的结构和功能,因此适合用于生产治疗性蛋白,如单克隆抗体、细胞因子等。
不同蛋白质表达系统的比较

不同蛋白质表达系统的比较蛋白质是细胞内重要的生物大分子,可以发挥许多生命活动的关键作用。
不同的蛋白质表达系统可以用于生产不同类型的蛋白质,比较蛋白质表达系统的优缺点对于选择合适的表达系统具有重要的意义。
本文将介绍现有的几种主要的蛋白质表达系统,并对它们的特点进行比较。
第一种蛋白质表达系统是基于真核细胞的表达系统。
真核细胞是具有细胞核的细胞,其中包括了动物细胞、植物细胞和真菌细胞等。
这种表达系统利用真核细胞的转录和翻译机制来制造目标蛋白质。
在这种系统中,目标基因被插入到真核细胞的基因组中,然后通过RNA剪切和mRNA成熟等机制生成成熟mRNA,从而进行翻译,最终目标蛋白质被产生出来。
真核细胞表达系统的优点包括:能够产生生物活性和功能齐全的蛋白质。
这种系统还适用于产生大量的蛋白质,因此被广泛应用于产生多肽、抗体等药物。
但是,真核细胞表达系统的劣势在于工艺更加复杂,容易出现蛋白质不稳定、失去生物活性的问题。
此外,该工艺需要一定的时间来建立并优化系统。
第二个系统是基于细菌的表达系统。
细菌是单细胞生物,具有非常简单的结构和进化历史,是蛋白质表达方面的主要模型。
在这种表达系统中,表达载体中的目标基因被转化为蛋白质,并通过重组不同的DNA序列来实现该过程。
这种系统的优点在于简单、实时、具有可伸缩性和高效性。
制备蛋白质的成本也相对较低。
然而,这种表达系统的制约因素也很明显。
细菌系统不能表达合成二硫键、表达动力多肽等蛋白质的功能,使其应用领域相对狭窄。
此外,细胞酸碱值和产生蛋白质的环境等因素,都是影响蛋白质表达的关键因素。
第三种蛋白质表达方式是基于哺乳动物细胞的表达系统。
哺乳动物细胞表达系统主要用于大规模制备人用蛋白质。
这种表达系统利用哺乳动物细胞的转录和翻译机制来制造目标蛋白质。
在这种系统中,目标基因经过一系列的转染和筛选过程,被转移到哺乳动物细胞中,利用细胞所提供的设备进行蛋白质合成和修饰。
哺乳动物表达系统具有高表达量、具备更完整的蛋白质修饰等诸多优点。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
动物基因工程原理
特点与优势
哺乳动物细胞表达系统的优势在于能够指导蛋白 质的正确折叠,提供复杂的N型糖基化和准确的O型 糖基化等多种翻译后加工功能,因而表达产物在分 子结构、理化特性和生物学功能方面最接近于天然 的高等生物蛋白质分子。
应用领域
哺乳动物细胞表达系统已成为多种基因工程药物 的生产平台。
已用该细胞株获得了包括人组织纤溶酶原激活剂、生长因 子等在内的多种蛋白的高效稳定表达。
对细胞株选择性地进行遗传改造
基于腺病毒EIA蛋白可反式激活CMV启动子,Cockett等建 立了稳定表达EIA的CHO细胞株,在该细胞中重组前胶原 酶的产量与CHO细胞相比增加了l2倍;该细胞在多种抗体 的表达中亦取得满意的结果。
哺乳动物细胞表达系统在新基因的发现、蛋白质 的结构和功能研究中亦起了极为重要的作用。
研究现状
部分蛋白在哺乳动物细胞中的表达已从实验室研究迈向生 产或中试生产阶段。
已有许多重要的蛋白及糖蛋白利用哺乳动物细胞系统表达 和大量制备、生产。如人组织型血纤蛋白酶原激活因子、 凝血因子Ⅷ、干扰素、乙肝表面抗原、红血球生成激素、 人生长激素、人抗凝血素Ⅲ,集落刺激因子等。
启动子需包含两个识别序列:mRNA转录起始点和TATA 盒。TATA盒位于转录起始位点上游25--30bp处,是引导 RNA聚合酶在正确起始位点转录所必需的序列,即保证转 录的精确起始。其他上游启动子元件常位于TATA盒上游 100~200bp之间,其功能是调节转录的起始频率和提高转 录效率。
真核表达载体-启动子
高等哺乳动物受体细胞的条件
以高效表达外源基因为目标的高等哺乳动物受体细胞应具 备以下条件: 细胞系特征:丧失细胞接触抑制和锚地依赖特征,便于大 规模培养。 遗传稳定性:外源基因多次传代后不至于丢失,易于长期 保存。 合适的标记:便于转化株的筛选和维持。 生长快且齐:分裂周期短,生长均一,便于控制。 安全性能好:不合成分泌致病物质,不致癌。
真核表达载体-启动子
另一类杂合启动子是由活性很低的启动子与数个转录激活 因子结合位点串联而成,这些位点与转录激活因子的结合 受细胞外小分子药物的调控;这类启动子主要用作基因的 诱导表达。
如Invitrogen公司的ecdysone(脱皮激素)诱导表达系统, 其负责目的基因转录的启动子是由热休克蛋白启动子核心 序列(minimal promotor)和5个E/GRE元件组成;
高等哺乳动物受体细胞的遗传标记
遗传标记 编码产物
筛选药物
作用机理
APH-
氨基糖苷磷酸转移酶 G418
APH灭活G418
DHFR HPHTK-
二氢叶酸还原酶 潮霉素B磷酸转移酶 胸腺嘧啶核苷激酶
氨甲喋呤 潮霉素B 氨甲喋呤
DHFR变体抗氨 甲碟呤
HPH灭活潮霉素 B
TK合成TMP
XGPRTADA-
黄嘌呤鸟嘌呤磷酸核糖 霉酚酸 转移酶
因而可通过控制 tet 的浓度来控制基因表达的水平。
▪ 应用于研究基因不同水平的表达对细胞,个体发育的影响
有何不同。
作用机制示意图
四环素调控表达系统构成元件
▪ 四环素阻遏子(Tet repressor protein,TetR) ▪ 四环素操纵子(Tet operator DNA Sequence, TetO) ▪ 单纯疱疹病毒Vp16转录活化区(Vp16 activation domain,AD) ▪ 四环素调控反式激活子(Tetracyeline-Controlled
对细胞株选择性地进行遗传改造
对细胞其它特性的遗传改造,包括细胞生长周期 的调控、细胞的抗凋亡能力、细胞贴壁能力、蛋 白糖基化的模式等方面,亦有相关报道,其实际 应用价值有待得到更多实验数据的支持。
真核表达载体
哺乳动物细胞表达载体的必要元件包括:一个高活性的启 动子、转录终止序列和一个有效的mRNA翻译信号。
IRES位于小RNA病毒RNA5’非编码区, 约为450bp。 IRES可与它连接的ORF同时翻译,构建双顺反子和 多顺反子真核表达载体。
分泌表达载体
可调控的哺乳动物细胞表达系统
四环素(tet)调控表达系统
▪ 加入或撤除四环素和四环素衍生物来调控基因表达。 ▪ 外源基因表达的水平与所加的 Tc 浓度在一定范围内相关,
启动子和增强子受细胞类型的限制,在不同的细 胞系中有很大不同,因此需根据宿主细胞的类型 选择不同的人巨细胞病毒早期启动 子(CMV-IE)、人延伸因子1-亚基启动子和Rous肉 瘤长末端重复序列;Invitrogen公司开发的pcDNA、 pEF和pRL三种系列载体即分别是以这三种启动子 驱动目的基因的表达。
哺乳动物细胞表达系统的表达水平从整个水平上看仍偏低, 一般处在杂交瘤细胞单克隆抗体蛋白产率的下限,即130μg/l08细胞/24小时。其限速步骤可能是在工程细胞中重 组蛋白的分泌效率较低。
一、稳定表达细胞系的筛选
构建目的基因的表达载体。 确定受体细胞。 确定筛选方法(抗生素的使用浓度)。 转染后48h,传代铺板 (1 : 100) ,加筛选药物。 3-4d换液1次,直至存活细胞长出岛状细胞簇,约2周。 克隆耐药细胞,检测目基因的存在与表达。
据报道红细胞生成素启动子在1%氧浓度比在21%氧浓度下活 性增强100倍。这又是一种新的提高表达量的作用机制。
真核表达载体-增强子
常用的增强子有Rous肉瘤病毒基因长末端重复序列和人巨 细胞病毒增强子。
James等利用糖皮质类固醇诱导的鼠乳房瘤病毒(MMTV)启 动子和鼠乳房瘤病毒长末端重复序列(MMTV-LTR),在 CHO细胞中表达分泌的碱性磷酸酶,产量为利用常规的 SV40和 CMV启动子的10倍,超过0.4mg/ml。
XGPRT和成 GMP
腺嘌呤核苷脱氨酶
腺嘌呤木酮糖苷 ADA-灭活Xyl-A
宿主细胞
常用的非淋巴细胞类有中国仓鼠卵巢(CHO)细胞、小仓鼠 肾(BHK)细胞、COS细胞、小鼠NSO胸腺瘤细胞和小鼠骨 髓瘤SP2/0细胞等。
不同宿主细胞表达的重组蛋白其稳定性和蛋白糖基化类型 不同,需根据要表达的目的蛋白选择最佳的宿主细胞。
常用的高等哺乳动物受体细胞
迄今为止,用于医疗用品(药物、抗体、诊断试剂)大规 模生产的高等哺乳动物受体细胞主要还是中国仓鼠卵巢细 胞(CHO),其优势有如下几个方面: 遗传背景清楚,生理代谢稳定 与人的亲缘关系接近,外源蛋白修饰准确 基因转移和载体表达系统完善 耐受剪切力,便于大规模培养 被美国FDA确认为安全的基因工程受体细胞
宿主细胞
猴肾细胞(COS)是进行外源基因瞬时表达时用途最广的 宿主,其重组载件易于组建,便于使用,而且对插入DNA 的量或者采用基因组DNA序列的情况都没有什么限制,便 于通过检测表达情况来确证cDNA的阳性克隆,也利于快 速分析引入克隆化cDNA序列中的突变。
宿主细胞
近几年也陆续发现了几种新的有较大应用价值的细胞株:如 来源于MadIin-Darby犬肾的高分化内皮细胞株(MDCK)。
内含子
某些内含子有重要的基因功能调控序列,目的基因以基因 组形式克隆入载体能得到高表达。
为增加转录产物的稳定性,表达载体中一般含有天然的或 人工合成的内含子序列。
mRNA前体的剪切能够促进mRNA 从胞核向胞质的运输。 外显子和内含子接头 每个外显子和内含子接头区都有一
段高度保守的一致序列(consensus sequence),即内含子 5’末端大多数是GT开始,3’末端大多数是AG结束,称为 GT-AG法则,是普遍存在于真核基因中RNA的识别信号。
原核DNA序列,包括能在大肠杆菌中自身复制的复制子,便 于筛选含重组细菌的抗生素抗性基因,以及便于目的基因 插入的限制性酶切位点。
可视实验需要加入标志基因、内含子、内部核糖体进入位 点等。
真核表达载体
常用的标记基因有胸腺激酶(tk)基因、二氢叶酸还原酶 (dhfr)基因、新霉素(neo)抗性基因、氯霉素乙酰基转移酶 (cat)基因等
Pei等用该细胞株表达分泌型的基质金属蛋白酶MMPI3,发现 高表达的阳性细胞克隆可占转染细胞的5%~l0%,其中一个 克隆的表达量可占细胞上清总蛋白的l5%~20%,在细胞单层 贴壁培养情况下表达量达10 mg/L。
对细胞株选择性地进行遗传改造
BHK/vl6细胞株是稳定表达单纯疱疹病毒(HSV)VP16蛋白 的BHK细胞,由于VP16的转录激活作用,载体中的HSV 早期启动子在该工程细胞中有很高的活性。
真核表达载体-启动子
近年来又发现了一些新的强启动子,如:人 leukosialin基因启动子和鼠3-磷酸甘油激酶l (PGKI) 基因启动子,活性与CMV-IE相当。
人泛素蛋白(ubiquitin)C基因启动子不仅具有较高 的活性,而且比CMV-IE、PGK1等启动子有更广 泛的宿主细胞范围,几乎在转基因小鼠的所有组 织细胞中都具有较高活性。
真核表达载体-启动子
构建杂合的启动子是获得新启动子的一个重要途径。
比如由人ubiquitin C启动区序列与CMV增强子组成 的杂合启动子、由SV40早期启动子和人I型T淋巴 细胞病毒LTR中的增强子序列(R-U5片段)组成的 SR-α启动子的活性均与CMV-IE相当;
由鸡β-肌动蛋白启动子和CMV增强子序列构成的 杂合启动子不仅活性比CMV—IE高,而且具有更 为广谱的宿主细胞范围。Novagen公司的pBacMam 表达系统即是以该杂合启动子驱动目的基因的转录。
二、哺乳动物细胞表达系统
(一)宿主细胞 (二)表达载体的构建 (三)表达细胞株的基因共圹增和筛选方法 (四)可调控的哺乳动物细胞表达系统
高等哺乳动物受体细胞
高等哺乳动物细胞的生长特性 高等哺乳动物受体细胞的条件 高等哺乳动物受体细胞的遗传标记 常用的高等哺乳动物受体细胞