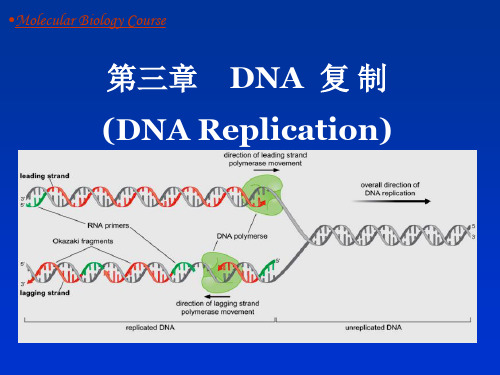
第3章 DNA的复制
分子生物学 第3章 DNA复制
DNA helicase (DNA解旋酶)
利用ATP供能,解开DNA双链, 可随复制叉 的伸展向前移动
大肠杆菌中解旋酶的种类
种 类
DnaA DnaB DnaC
功 能
辨认起始点,并结合到复制起始部位 解开DNA双链 运送和协同DnaB
single-stranded binding protein (SSB, 单链结合蛋白)
是一类调节DNA分子的超螺旋水平,可改变DNA拓扑性 质的酶。对DNA分子的作用是既能水解、又能连接磷酸 二酯键。 • 拓扑异构酶 I: 切开DNA双链中的一股,使DNA在解链旋 转中不打结,DNA变为松弛状态再封闭切口。 同转录有 关 • 拓扑异构酶 II: 能切断DNA双链,使螺旋松弛。在ATP参 与下,松弛的DNA进入负超螺旋,再连接断端。同复制
3´→5´外切酶活性: 切除错配的核苷酸
5'
3' C T T C A G G A G A A G T C C G G C G 5'
3'
DNA ligase
连接DNA链3-OH末端和相邻DNA链5-P末端,形成磷 反应需要ATP。
酸二酯键,从而把两段相邻的DNA链连接成完整的链。
二、 DNA复制的过程
E. Coli DNA在15N-标记的营养液中生
长多代,使DNA双链充分标记
将15N-标记
细胞在
14N中
细胞在
14N中复
细胞在
14N中复
的E.Coli 加入14N 培 养液中
万有引力
复制1 次
制第2次
制第3次
单林娜 制作
11
DNA半保留复制的生物学意义:
DNA的半保留复制表明DNA在代谢上的稳定性,
D第3章 第3节DNA的复制2014-5-8

DNA分子的两条链为模板
细胞中游离的4种脱氧核苷酸(ATCG)
酶
能量
解旋酶、DNA聚合酶
ATP
三、DNA复制的过程
5、过程: 解旋
碱基配对 形成子代DNA (合成子链)
P54
解旋: 解旋酶催化(氢键断裂)
模板 同时进行(边解旋边复制)
2:与复制有关的碱基计算
亲代DNA分子
复制1次
2
复制2次
复制3次
22
23
… …
复制n次
2n
与复制有关的碱基计算
①一个DNA连续复制n次后,共有多少个DNA?多少条脱氧核苷酸链? 母链多少条?子链多少条? 解析:所用知识为“半保留复制”和“碱基互补配对原则”,并图示 分析。 A T
TA GC CG
P54
复制 以母链为模板在DNA聚合酶的 : 催下,利用游离的脱氧核苷酸 进行碱基互补配对 形成子代 DNA: 组成 母链(旧链) 子链(新链)
三、DNA复制的过程
5、过程: 解旋
碱基配对 形成子代DNA (合成子链)
P54
6、特点: ①边解旋边复制 ②半保留复制
★原则:碱基互补配对原则
7、结果:1个DNA分子 两个完全相同的DNA分子 8、精确复制的原因: ⑴独特的双螺旋结构为复制提供了精确的模板
连续第一次复制
A T TA GC CG A T TA GC CG
根据半保留复制和碱基互补配对原则 解: n DNA分子数= 2
n+1 脱氧核苷酸链数= 2
连续第二次复制
A T TA GC CG A T TA GC CG A T TA GC CG A T TA GC CG
高中生物人教(2019)必修2课件第3章第3节 DNA的复制

2.DNA 复制的过程
能量 解旋酶
螺旋 母链
脱氧核苷酸 DNA 聚合酶
碱基互补配对
模板链
双螺旋结构
3.DNA 复制的特点 (1)过程:_边__解__旋__边__复__制___,多起点复制。 (2)方式:半保留复制。 4.准确复制的原因 (1)DNA 独特的双螺旋结构,为复制提供了精确的模板。 (2)通过_碱__基__互__补__配__对___,保证了复制能够准确地进行。 5.DNA 复制的意义 将遗传信息从亲代细胞传递给子代细胞,从而保持了_遗__传__信__息___的连续性。
A.酶①和酶②均作用于氢键 B.该过程的模板链是 a、b 链 C.该过程可发生在细胞有丝分裂间期 D.DNA 复制的特点是半保留复制 解析:由题图可知,酶①(解旋酶)使氢键断裂,而酶②(DNA 聚合酶)催化形成磷酸 二酯键,A 错误。答案:A
2.真核细胞中 DNA 复制如下图所示,下列表述错误的是
结果如下图所示。
(1)Ⅰ代细菌 DNA 分子的结构是怎样的? 提示:Ⅰ代细菌 DNA 分子中只有一条链被 15N 标记。 (2)Ⅲ代细菌 DNA 分子的平均相对分子质量是多少? 提示:Ⅲ代细菌 DNA 分子的平均相对分子质是为(7a+b)/8。
1.关于 DNA 分子复制的早期推测 在 DNA 分子复制的早期研究中,科学家们提出了三个模型:全保留复制模型、弥 散复制模型和半保留复制模型。 比较如下: 全保留复制:亲代 DNA 分子两条链不变,子代 DNA 分子的两条链都是新合成的。 半保留复制:新合成的每个 DNA 分子中,都保留了原来 DNA 分子中的一条链。 弥散复制:亲代 DNA 分子的两条链分散成短片段,与新合成的子代 DNA 分子的 两条链分散成的短片段混杂在一起,不能分出亲代 DNA 单链。
《DNA的复制》PPT课件

子代DNA:
母链(旧链) 组成
子链(新链)
边
半
解
保
旋
留
复
复
制பைடு நூலகம்
制
多
起
点
复
制
具有100个碱基对的1个DNA分子片断, 内含40个胸腺嘧啶,如果连续复制两次, 则需要游离的胞嘧啶脱氧核苷酸的数目为 ( 180 )个。
某DNA分子共含有含氮碱基1400个,其中一 条链上A+T/C+G= 2:5,问该DNA分子连续复制 2次,共需游离的胸腺嘧啶脱氧核苷酸的数目是
7、准确复制原因 ①DNA双链提供精确的模板
②碱基互补配对原则
8、复制的生物学意义:P54
【智慧眼——寻找规律】
规律2:亲代DNA分子经 n 次复制后, 所需某种游离的脱氧核苷酸数为:
R =a (2 n-1) 其中 a 表示亲代DNA含有的某种
碱基数,n 表示复制的次数。
规律3:碱基总数=失去H2O数+2
来的科学研究发现,小鼠体内的HMGIC基因与肥胖 直接相关。具有HMGIC基因缺陷的实验鼠与作为对 照的小鼠,吃同样多的高脂肪食物,一段时间后, 对照组的小鼠变得十分肥胖,而具有HMGIC基因缺 陷的实验鼠体重仍然保持正常。
没有HMGIC基因,就没有肥胖的表现,有HMGIC基因就有 肥胖表现。说明基因能控制生物的性状(功能单位)。
1三、概、念D:NA分子复制的过程(P54)
2、场所:细胞核(主要)、线粒体、叶绿体
3、时期:有丝分裂间期、减数分裂第一次分裂的间期
模板:DNA的两条母链
4、条件
原料:游离的脱氧核苷酸(A、G、C、T) 能量:ATP
酶:DNA解旋酶、DNA聚合酶等
高中生物必修二第三章第3节 DNA分子的复制

活动任务----演绎推理:
请依据两种假说分别演绎推理15N标记的DNA在14N的培养 基中培养1代前后的DNA,并分别预测两种假说第0代和第1代 DNA密度梯度离心后的结果,并画在离心管相应的位置上。
实验结果:
大肠杆菌在含15NH4Cl的 培养液中生长若干代
转移到含14NH4Cl 的培养液中
15N/15N DNA
A.每条染色体的两条单体都有被标记
B.每条染色体中都只有一条单体被标记
C.只有半数的染色体中一条单体被标记
D.每条染色体的两条单体都不被标记
4、用P标记了玉米体细胞(含20条染色体)的 DNA分子双链,再将这些细胞转入不含32P的 培养基中培养,在第二次细胞分裂的中期、后 期,一个细胞中的染色体总条数和被32P标记 的染色体条数分别是
2.半保留复制
新合成的DNA 分子一半新的, 一半旧的
3.分散复制
新合成的DNA分子新的和旧 的都有
1956年,两位年轻的美国分子生物学家梅塞尔森和斯塔 尔合作开展关于DNA复制的实验研究,实验结果于1958 年正式发表。
关键问题1: 肉眼看不见的DNA分子,用什么方法区分
亲代和子代的DNA单链?
( )个;第4次复制时需要游离的胞嘧 啶脱氧核苷酸的数目为 ( )个
五、DNA复制与细胞分裂的关系:
进行第一次有丝分裂:
进行第二次有丝分裂:
1. 蚕豆根尖细胞在含3H标记的胸腺嘧啶脱氧核苷酸 培养基中完成一个细胞周期,然后在不含放射性标记 的培养基中继续分裂至中期,其染色体的放射性标记 分布情况是( )
A.中期20和20、后期40和20 B.中期20和10、后期40和20 C.中期20和20、后期40和10 D.中期20和10、后期40和10
第三章 DNA的复制

(1)端粒和端粒酶的发现
1978 年 , Blackburn 发现四膜虫大核中 rDNA 小分 子 末 端 的 端 粒 结 构 为 370520bp 的 (GGGGTT)n 重复片段。
加尾实验 1984
加尾实验 1985
四膜虫抽提液
酵母 末端重复序列
端 粒 酶 的 鉴 定
1985
端粒酶的分离纯化
TA
母代DNA 子代DNA
半保留复制的意义
按半保留复制方式,亲代DNA所含的信 息以极高的准确度传递给子代DNA分子,子 代保留了亲代的全部遗传信息 ,体现了遗 传的保守性。
遗传的保守性,是物种稳定性的分子基 础,但不是绝对的。
3.1.2 复制叉和复制体
复制叉:发生复制的 位点,或者称为生 长点。
后随链:背向复制叉,一段亲本DNA链先暴露 出来才能以相反方向合成DNA小片段,然后 这些小片段DNA连接形成完整的后随链。
冈崎的实验—脉冲标记实验
lig-突变体
冈崎的实验—脉冲追踪实验
3.1.5复制的起点、方向
复制起点(origin of replication,ori)
原核生物复制起始位点区特点
Dolly 1996-2003
端粒酶和永生
3.3 DNA复制的终止
ColE I
3.4 DNA复制的调控
质 粒 的 复 制 调 控
真核生物的DNA复制的调控
GLN1 GLN2 GLN3
cyclin
p34
MPF
cdc6,cdc8, cdc9,cdc21
3.2.2 多复制子复制的非一致性
每个复制子发动复制的先后时序有很大区别: 同一染色体上不同复制子之间 不同类型细胞之间
复制子的多少与DNA复制的速度有关 基因组的复制完成与细胞、组织及发育状态有 关。
高中生物必修二 学习笔记 第3章 第3节 DNA的复制

第3节DNA的复制[学习目标] 1.运用假说—演绎法探究DNA的复制方式,概述DNA通过半保留的方式进行复制。
2.理解DNA的准确复制是遗传信息稳定传递的基础。
一、对DNA复制的推测及DNA半保留复制的实验证据1.对DNA复制的推测(1)半保留复制①提出者:______________。
②观点:DNA复制方式为____________。
③内容:DNA复制时,DNA双螺旋解开,互补的碱基之间的________断裂,解开的两条单链分别作为复制的模板,游离的____________根据____________原则,通过形成________,结合到作为模板的单链上。
④结果:新合成的每个DNA分子中,都保留了原来DNA分子中的________。
(2)全保留复制:指DNA复制以DNA双链为模板,子代DNA的双链都是________的。
2.DNA半保留复制的实验证据(1)实验方法:____________技术和____________技术。
(2)实验原理:只含15N的DNA密度____,只含14N的DNA密度____,一条链含14N、一条链含15N的双链DNA密度_______________________________________________。
因此,利用______技术可以在试管中区分含有不同N元素的DNA。
(3)探究DNA的复制方式①提出问题:DNA以什么方式复制?②作出假设:DNA以__________________________________________________________方式复制。
③演绎推理(预期实验结果)离心后应出现____条DNA带;a.重带(密度最大):两条链都为______标记的亲代双链DNA。
b.中带(密度居中):一条链为14N标记,另一条链为15N标记的子代双链DNA。
c.轻带(密度最小):两条链都为______标记的子代双链DNA。
④实验验证实验结果条带数量在试管中位置DNA含N情况亲代靠近试管底部15N/15N-DNA 第一代位置居中第二代一条带位置居中,一条带位置靠上⑤实验结论:DNA的复制是以__________的方式进行的。
3.3 DNA的复制

聚焦三:DNA复制中的变异
碱基互补配对差错导致基因突变
聚焦四:DNA复制中的有关计算
15N
14N
【分析】
亲代DNA分子
复制1次
2=21
4 =22 8 =23
复制2次
复制3次
无论DNA复制多少次,含有原来母链的DNA分子 永远只有两条
思维拓展
1)复制n次后DNA个数:
Ⅱ 2n(n=复制次数) Ⅲ Ⅰ
1.概念: 是指以亲代DNA为模板 合成子代DNA的过程。 2.时间: 细胞有丝分裂的间期和 减数第一次分裂的间期 3.场所: 细胞核(主要)
模板: DNA分子的两条链。
4.条件
原料: 细胞中游离的4种脱氧核苷酸。 能量: ATP。 酶: DNA解旋酶,DNA聚合酶等。
适宜的环境
5.过程
解旋:
DNA利用ATP,在解旋酶的作用下, 把两条螺旋的双链解开。 以解开的每一条母链为模板,在 DNA聚合酶等的作用下,将游离的4种 脱氧核苷酸按碱基互补配对原则,各 自合成与母链互补的一段子链。
亲代DNA分子
问题2:如果要在实验中直观地区别、“标识” 母链或子链,可以采取什么办法?
同位素(具放射性)标记的方法
亲代DNA分子
问题3:如果用同位素(
放射性)进行标记,用 什么元素? 可用C、H、O、 N、P等元素 问题4:如果亲代DNA是15N的,放在14N的环境 中进行培养,则亲代、子一代、子二代DNA分 别含有哪种N元素?
复制前
亲代DNA分子
复制后
子一代: 15N/14N-DNA (全部) 子二代: 15N/14N-DNA(1/2) 14N/14N-DNA(1/2) 问题5: 要验证上述预测,就要分别观察亲代和子 代的情况,但实验中,复制后的DNA分子混合在一起 的,不易分离。怎么解决这个问题?
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
DNA聚合酶催化的链延长反应
3´ 5´ 5´
3´
3´
3´
5´
3´
5´ 5´
模板链
RNA引物
5´
子链
3´
DNA聚合酶(DNA Polymerase)
共同性质
[1] 以脱氧核苷酸三磷酸 (dNTP) 为前体催化合成 DNA;
[2]需要模板和引物的存在; [3]不能起始合成新的DNA链; [4]催化dNTP加到生长中的DNA链的3'-OH末端; [5]催化DNA合成的方向是5'→3'。
思考
楚雄师范学院化学与生命科学系 范树国
遗传信息传递的 中心法则
生物的遗传信息以密码的形式储 存在DNA分子上,表现为特定的核苷 酸排列顺序。在细胞分裂的过程中, 通过DNA复制把亲代细胞所含的遗传 信息忠实地传递给两个子代细胞。在 子代细胞的生长发育过程中,这些遗 传信息通过转录传递给 RNA ,再由 RNA通过翻译转变成相应的蛋白质多 肽链上的氨基酸排列顺序,由蛋白质 执行各种各样的生物学功能,使后代 表现出与亲代相似的遗传特征。后来 人们又发现,在宿主细胞中一些RNA 病毒能以自己的RNA为模板复制出新 的病毒RNA,还有一些RNA病毒能以 其RNA为模板合成DNA,称为逆转录 这是中心法则的补充。
第三章 DNA的复制
本章重点介绍遗传中心法则和 DNA 的半 保留复制以及逆转录的过程和机理,对DNA 的损伤和修复、突变和重组作一般介绍。
DNA 是绝大多数生物体遗传信息的载体,继 1953年Watson & Crick提出DNA双螺旋结构模型 后, 1958 年, Crick 提出了‚中心法则‛ (Central dogma)揭示了遗传信息的传递规律。
楚雄师范学院化学与生命科学系 范树国
单链DNA结合蛋白(SSBP)
螺旋酶沿复制叉方向向前推进产生了一段单链区,但是这种单链 DNA不会长久存在,会很快重新配对形成双链 DNA或被核酸酶降解 。在细胞内有大量单链DNA结合蛋白能很快地和单链DNA结合,防 止其重新配对形成双链 DNA或被核酸酶降解。一个SSBP四聚体结 合于单链DNA上可以促进其他SSBP四聚体现相邻的单链DNA结合, 这个过程称为协同结合 (cooperative binding),SSBP结合到单 链DNA上后,使其呈伸展状态,没有弯曲和结节,有利于单链DNA 作为模板。SSBP 可以重复使用,当新生的DNA链合成到某一位置 时,该处的SSBP便会脱落,并被重复利用。
楚雄师范学院化学与生命科学系
范树国
[2]3'→5'外切酶活性──校对作用
这种酶活性的主要功能是从3'→5'方向识别和切除 不配对的DNA生长链末端的核苷酸。
楚雄师范学院化学与生命科学系
范树国
[3]5'→3'外切酶活性──切除修复作用 从5’→3’方向水解 DNA生长链前方的 DNA链,主 要产生5’-脱氧核苷酸。这种酶活性只对DNA上配对 部分(双链)磷酸二酯键有切割活力作用。每次能切除 10个核苷酸,而且DNA的聚合作用能刺激5'→3'外切 酶活力达 10 倍以上。因此,这种酶活性在 DNA 损伤 的修复中可能起着重要作用。对冈崎片段5'末端DNA 引物的去除依赖此种外切酶活性。
螺旋酶(helicase)可以促使DNA在复制叉处打 开双链。螺旋酶可以和单链 DNA 结合,并且与 ATP 结合,利用 ATP 分解成 ADP 时产生的能量沿 DNA 链向前运动促使 DNA 双链打开。目前,大肠 杆菌中发现有两种螺旋酶参与这个过程,一种 称为螺旋酶Ⅱ或螺旋酶Ⅲ,与前导链的模板DNA 结合沿 5 '→ 3' 方向运动 ; 第二种称为 Rep 蛋白, 和前导链的模板DNA结合沿3'→5'方向运动。
DNA PolⅠ在空间结构上近似球体,直径约6.5 nm。 在酶的纵轴上有一个约 20A 的深沟 (cleft) ,带有正电 荷,这是该酶的活性中心位置,在此位置上至少有 6 个结合位点: [1] 模板DNA结合位点; [2] DNA生长链 或引物结合位点; [3] 引物末端结合位点,用以专一引 物或DNA生长链的3'-OH; [4] 脱氧核苷三磷酸结合位 点; [5] 5'→3'外切酶活性位点,用以结合生长链前方 的5'-端脱氧核苷酸并切除之; [6] 3'→5'外切酶活性位 点,用以结合和切除生长链上未配对的
楚雄师范学院化学与生命科学系
范树国
(3)该酶对作用底物的选择性较强,一般只能将2
-脱氧核苷酸掺入到DNA链中。
(4)该酶不是复制的主要聚合酶,因为此酶缺陷的
大肠杆菌突变株的DNA复制都正常。可能在DNA
的损伤修复中该酶起到一定的作用。
楚雄师范学院化学与生命科学系
范树国
大肠杆菌DNA聚合酶Ⅰ
这是1956年由Arthur Kornberg 首先发现的 DNA 聚合 酶,又称Kornberg酶。此酶研 究得清楚而且代表了其他 DNA 聚合酶的基本特点。
DNA的损伤与修复
DNA突变 DNA的遗传重组
楚雄师范学院化学与生命科学系
范树国
DNA生物合成有两种方式
1. DNA的复制(DNA指导下的DNA合成)
DNA体内复制:原核、真核生物的染色体、细菌质 粒(环状,双链)、真核细胞器DNA(线粒体、叶 绿体)、病毒(双链,环状) DNA的体外复制:分子克隆。
[1]聚合作用 在引物 RNA3'-OH 末端,以 dNTP 为底物,按模板 DNA上的指令由DNA polⅠ逐个将核苷酸加上去, 就是 DNA polⅠ的聚合作用。酶的专一性主要表现 为新进入的脱氧核苷酸必须与模板 DNA配对时才有 催化作用。dNTP进入结合位点后,可能使酶的构象 发生变化,促进3'-OH与5'-PO4结合生成磷酸二酯键 。若是错误的核苷酸进入结合位点,则不能与模板 配对,无法改变酶的构象而被 3'-5'外切酶活性位点 所识别并切除之。
楚雄师范学院化学与生命科学系
范树国
大肠杆菌三种DNA聚合酶比较
比较项目 分子量 每个细胞的分子统计数
DNA DNA DNA 聚合酶Ⅰ 聚合酶Ⅱ 聚合酶Ⅲ
109,000 400 120,000 400,000 100 10-20
5´-3 ´聚合酶作用
3´-5 ´核酸外切酶作用 5´-3 ´核酸外切酶作用 转化率 功能
楚雄师范学院化学与生命科学系 范树国
DNA在细胞内往往以超螺旋状态存在, DNA 拓扑酶催化同一 DNA 分子不同超螺 旋状态之间的转变。DNA拓扑异构酶有 两类,大肠杆菌的ε蛋白(MW-110000) 就是一种典型的拓扑异构酶Ⅰ.它的作 用是暂时切断一条DNA链,形成酶-DNA 共价中间物而使超螺旋 DNA松弛化,然 后再将切断的单链 DNA连接起来,而不 需要任何辅助因子。而大肠杆菌中的 DNA旋转酶(DNA gyrase)则是典型的拓 扑异构酶Ⅱ,能将负超螺旋引入DNA分 子,该酶能暂时性地切断和重新连接 双链DNA,同时需要ATP水解为ADP以供 能。
DNA聚 合酶III DNA聚 合酶I
5´
3´
解旋酶 解链酶 引物酶和 引发体
SSB
RNA引物
3´ 5´ RNA
引物
3´
5´
大肠杆菌的引物酶也是 DNA 复制所必需的一种酶。 该酶由一条多肽链组成, MW为 60 KD ,每个细胞中 有50-100个分子由大肠杆菌的dnaG基因编码。引物 酶催化引物RNA分子的合成,但它和传统的RNA聚合 酶不一样。[1]引物酶对雷米封不敏感,而RNA聚合 酶则敏感 ;[2] 引物酶既可以利用核糖核苷酸,而 RNA 聚合酶只能利用核糖核苷酸 ;[3]引物酶只在复 制起点处合成 RNA 引物而引发 DNA 的复制,而RNA 聚 合酶则是启动 DNA 转录合成 RNA 从而将遗传信息由 DNA 传递到 RNA 。但在单链噬菌体 M13DNA 和质粒 Co1E1DNA 复制时,引物的合成是由 RNA 聚合酶催化 的。噬菌体 T7的Gene4 蛋白,T4 的Gene41和61蛋白 等也具有引物酶的活性和具有大肠杆菌引物酶相似 的功能。
复制
DNA
转录 逆转录 RNA 复制 蛋白质
翻译ห้องสมุดไป่ตู้
中心法则总结了生物体内遗传信息的流动规律,揭示遗传的分子基础,不仅使人 们对细胞的生长、发育、遗传、变异等生命现象有了更深刻的认识,而且以这方面的 理论和技术为基础发展了基因工程,给人类的生产和生活带来了深刻的革命。
目
第一节
录
DNA生物合成
第二节
第三节 第四节
大肠杆菌DNA聚合酶Ⅱ(DNApolⅡ)
1970年发现了DNA pol Ⅱ。此酶MW为120 KD,每 个细胞内约有100个酶分子,但活性只有DNA polⅠ 的5%。
(1) 聚合作用 : 该酶催化 DNA 的聚合,但是对模板 有特殊的要求。该酶的最适模板是双链 DNA 而中间 有空隙(gap)的单链DNA部份,而且该单链空隙部份 不长于100个核苷酸。对于较长的单链DNA模板区该 酶的聚合活性很低。 (2)该酶具有 3 '→5' 外切酶活性,但无 5 '→3' 外切 酶活性。
DNA的半保留复制可以说明 DNA在代谢上
的稳定性。经过多代复制, DNA 的多核苷
酸链仍可以保持完整,并存在于后代而不
被分解掉。
楚雄师范学院化学与生命科学系
范树国
DNA的半保留复制实验依据
1958年Meselson & stahl用同位素示踪标记加密度梯度离心技术实验,
证明了DNA是采取半保留的方式进行复制.
楚雄师范学院化学与生命科学系
范树国
复制中的大肠杆菌染色体放射自显影图 (Cairns实验)