乙酰辅酶A羧化酶(ACC)活性检测试剂盒说明书可见分光光度法
NEFA游离脂肪酸测定试剂盒(ACS-ACOD法)产品说明书

游离脂肪酸测定试剂盒(ACS-ACOD 法)说明书【产品名称】通用名称:游离脂肪酸测定试剂盒(ACS-ACOD 法)英文名称:Nonestesterified fatty acid Assay Kit (Enzymic Method)(NEFA )【包装规格】R1:2⨯60ml 、R2:2⨯20ml ;R1:2⨯45ml 、R2:2⨯15ml ;R1:1⨯45ml 、R2:1⨯15ml ;校准品(选配):1⨯2ml (1个水平)。
【预期用途】用于体外定量测定人血清中游离脂肪酸的含量。
临床上主要用于高血脂症、冠心病和动脉粥样硬化的辅助诊断。
【检验原理】游离脂肪酸和辅酶A 在乙酰辅酶A 合成酶(ACS )的作用下反应生成乙酰辅酶A 。
乙酰辅酶A 在乙酰辅酶A 氧化酶(ACOD )的作用下生成H 2O 2,随后通过Trinder ’s 底物在过氧化物酶(POD )的作用下生成有色物质。
【主要组成成分】由试剂R1、R2和校准品组成。
试剂R1:乙酰辅酶A 合成酶(ACS )0.8KU/L 、MgCl 2(氯化镁)5mmol/L 、吐温-20 0.1%;试剂R2:乙酰辅酶A 氧化酶(ACOD)20KU/L 、过氧化物酶( POD )30KU/L 、吐温-20 0.1%;校准品:含十六(烷)酸水溶液。
校准品可以溯源至北京华宇亿康校准品,注:校准品浓度见每批瓶标示。
不同批号试剂盒中各组分不可以互换。
【储存条件及有效期】试剂和校准品在2℃~8℃避光条件下保存可以稳定365天。
试剂和校准品开瓶后2℃~8℃可稳定15天。
备注:生产日期及失效日期见外盒或瓶标签。
【适用仪器】日立7180、奥林巴斯AU680、贝克曼LX-20/DXC800、迈瑞BS-380、朕江T900全自动生化分析仪。
【样本要求】1、空腹静脉采血,样本为新鲜的血清样本。
2、样本采集后立即离心分离,并在当日检测,如当日不能检测,冷藏2℃~8℃下可稳定48h ;避免反复冻融。
辅酶 II NADP(H)含量检测试剂盒说明书
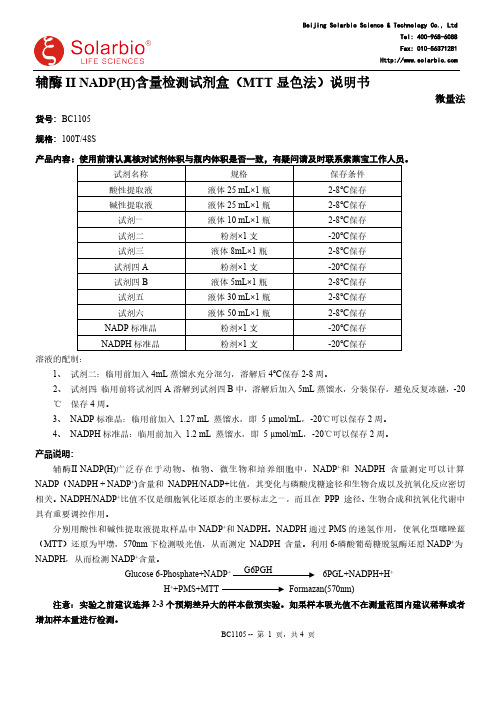
辅酶II NADP(H)含量检测试剂盒(微量法货号:BC1105规格:100T/48S产品内容:使用前请认真核对试剂体积与瓶内体积是否一致,有疑问请及时联系索莱宝工作人员。
试剂名称规格保存条件酸性提取液液体25 mL×1瓶2-8℃保存碱性提取液液体25 mL×1瓶2-8℃保存试剂一液体10 mL×1瓶2-8℃保存试剂二粉剂×1支-20℃保存试剂三液体8mL×1瓶2-8℃保存试剂四A粉剂×1支-20℃保存试剂四B液体5mL×1瓶2-8℃保存试剂五液体30 mL×1瓶2-8℃保存试剂六液体50 mL×1瓶2-8℃保存NADP标准品粉剂×1支-20℃保存NADPH标准品粉剂×1支-20℃保存溶液的配制:1、试剂二:临用前加入4mL蒸馏水充分混匀,溶解后4℃保存2-8周。
2、试剂四:临用前将试剂四A溶解到试剂四B中,溶解后加入5mL蒸馏水,分装保存,避免反复冻融,-20℃保存4周。
3、NADP标准品:临用前加入1.27 mL 蒸馏水,即5 µmol/mL,-20℃可以保存2周。
4、NADPH标准品:临用前加入1.2 mL 蒸馏水,即5 µmol/mL,-20℃可以保存2周。
产品说明:辅酶Ⅱ NADP(H)广泛存在于动物、植物、微生物和培养细胞中,NADP+和NADPH 含量测定可以计算NADP(NADPH + NADP+)含量和NADPH/NADP+比值,其变化与磷酸戊糖途径和生物合成以及抗氧化反应密切相关。
NADPH/NADP+比值不仅是细胞氧化还原态的主要标志之一,而且在PPP 途径、生物合成和抗氧化代谢中具有重要调控作用。
分别用酸性和碱性提取液提取样品中NADP+和NADPH。
NADPH通过PMS的递氢作用,使氧化型噻唑蓝(MTT)还原为甲瓒,570nm下检测吸光值,从而测定NADPH 含量。
乙酰辅酶 a 羧化酶基因 -回复

乙酰辅酶a 羧化酶基因-回复乙酰辅酶A 羧化酶基因是人类基因组中的一种酶基因,它在人体中起着重要的调节作用。
本文将会一步一步回答与该基因相关的问题,并解释其在人体中的重要功能。
第一步:什么是乙酰辅酶A 羧化酶基因?乙酰辅酶A 羧化酶基因是人体中一种编码羧化酶(carboxylase)的基因。
羧化酶是一类酶,它在体内参与乙酰辅酶A 代谢过程中的反应。
乙酰辅酶A 是一种核苷酸,广泛存在于各个细胞中,并在许多生物化学反应中发挥重要作用。
第二步:乙酰辅酶A 的主要功能是什么?乙酰辅酶A 在细胞能量代谢中起着重要的作用。
它作为多种酶的底物参与各种代谢反应,包括脂肪酸合成、胆固醇合成和脂质代谢等。
此外,乙酰辅酶A 也参与氨基酸和有机酸代谢,并在糖原合成和糖原分解中起到调节作用。
第三步:乙酰辅酶A 羧化酶基因突变会导致什么结果?乙酰辅酶A 羧化酶基因的突变可能会导致乙酰辅酶A 代谢途径发生紊乱。
这可能导致一系列的代谢紊乱,并进一步导致一些疾病。
例如,某些突变可能导致脂肪酸过氧化和酮体生成的异常,可能引发脂代谢障碍和某些代谢性疾病。
第四步:乙酰辅酶A 羧化酶基因和疾病之间有何关联?乙酰辅酶A 羧化酶基因的突变与一些疾病的发生和发展密切相关。
例如,某些突变可能导致乙酰辅酶A 羧化酶活性降低,从而引发婴儿中的生命威胁型代谢性疾病-短链脂肪酸羧化酶缺乏症。
这种疾病主要影响婴儿的能量代谢和脑功能。
然而,并非所有的突变都会导致疾病,因为人体通常会有多个代谢途径来保持代谢平衡。
第五步:如何识别乙酰辅酶A 羧化酶基因的突变?识别乙酰辅酶A 羧化酶基因的突变通常需要进行基因测序。
通过对该基因进行测序,可以检测出其中的突变情况。
此外,也可以进行相关的生化分析,以评估乙酰辅酶A 代谢通路的功能。
第六步:如何处理乙酰辅酶A 羧化酶基因突变引起的疾病?对于突变引起的疾病,治疗方法通常会因疾病的不同而有所不同。
一般而言,治疗的目标是通过调整患者的饮食习惯和用药治疗来维持身体的代谢平衡。
acc 代谢产物

acc 代谢产物
乙酰辅酶A羧化酶(ACC)是脂肪酸合成的关键酶,能够催化乙酰辅酶A(Acetyl CoA)羧化为丙二酸单酰辅酶A(Malonyl CoA)。
丙二酸单酰辅酶A是脂肪酸合成的中间产物,同时也是脂肪酸氧化的关键代谢产物。
此外,丙二酸单酰辅酶A还是胆固醇和其他脂质合成的重要前体。
在脂肪酸合成过程中,丙二酸单酰辅酶A可以通过缩合反应生成长链脂肪酸,也可以作为不饱和脂肪酸的合成原料。
另外,丙二酸单酰辅酶A还是胆固醇合成的中间产物,可以转化为甲羟戊酸,进而合成胆固醇和其他固醇类物质。
总的来说,丙二酸单酰辅酶A是脂肪酸和胆固醇等脂质合成的重要代谢产物,其合成和代谢途径受到ACC 等多种酶的调节。
了解其合成和代谢过程有助于深入理解脂质代谢的调控机制,对于预防和治疗脂质代谢相关疾病(如肥胖、糖尿病、高血脂等)具有一定的指导意义。
EMS_诱变创制水稻抗乙酰辅酶A_羧化酶抑制剂类除草剂种质

江苏农业学报(JiangsuJ.ofAgr.Sci.)ꎬ2023ꎬ39(2):305 ̄312http://jsnyxb.jaas.ac.cn江㊀群ꎬ凌溪铁ꎬ唐兆成ꎬ等.EMS诱变创制水稻抗乙酰辅酶A羧化酶抑制剂类除草剂种质[J].江苏农业学报ꎬ2023ꎬ39(2):305 ̄312.doi:10.3969/j.issn.1000 ̄4440.2023.02.001EMS诱变创制水稻抗乙酰辅酶A羧化酶抑制剂类除草剂种质江㊀群1ꎬ㊀凌溪铁2ꎬ㊀唐兆成2ꎬ㊀周珍珍2ꎬ㊀张保龙1ꎬ2(1.海南大学热带作物学院/三亚南繁研究院ꎬ海南海口570228ꎻ2.江苏省农业科学院种质资源与生物技术研究所/江苏省农业生物学重点实验室ꎬ江苏南京210014)收稿日期:2022 ̄11 ̄25基金项目:江苏省农业科技自主创新基金项目[CX(21)2041]作者简介:江㊀群(1998-)ꎬ女ꎬ四川宜宾人ꎬ硕士研究生ꎬ主要从事水稻抗除草剂育种研究ꎮ(E ̄mail)1692579264@qq.com通讯作者:张保龙ꎬ(E ̄mail)zhbl2248@hotmail.comꎻ周珍珍ꎬ(E ̄mail)zhenzhenzhounj@163.com㊀㊀摘要:㊀创制非转基因抗除草剂水稻种质资源对于稻田杂草防控具有重要价值ꎮ本研究以甲基磺酸乙酯(EMS)水溶液诱变镇糯19水稻种子ꎬ获得1株能稳定遗传的可耐受乙酰辅酶A羧化酶(ACCase)抑制剂类除草剂高效盖草能的M3代水稻幼苗(突变体)ꎮ分别扩增镇糯19野生型和突变体的基因组DNA并进行测序和序列比对ꎬ发现突变体ACCase基因的开放阅读框(ORF)的第5374位碱基发生了点突变ꎬ导致编码的第1792位氨基酸由异亮氨酸突变为亮氨酸ꎮ镇糯19野生型和突变体分蘖盛期大田喷施3种田间推荐剂量的ACCase抑制剂类除草剂后农艺性状调查结果表明突变体对高效盖草能㊁精喹禾灵和唑啉草酯抗性明显高于野生型ꎮ本研究获得了能稳定遗传的非转基因抗AC ̄Case抑制剂类水稻新种质ꎬ具有一定的应用价值ꎬ为抗除草剂水稻育种提供了种质资源ꎮ关键词:㊀水稻ꎻ抗除草剂种质ꎻ甲基磺酸乙酯(EMS)ꎻ乙酰辅酶A羧化酶中图分类号:㊀S335.3㊀㊀㊀文献标识码:㊀A㊀㊀㊀文章编号:㊀1000 ̄4440(2023)02 ̄0305 ̄08EMSmutagenesistocreatericeanti ̄acetyl ̄CoAcarboxylaseinhibitor ̄her ̄bicidegermplasmJIANGQun1ꎬ㊀LINGXi ̄tie2ꎬ㊀TANGZhao ̄cheng2ꎬ㊀ZHOUZhen ̄zhen2ꎬ㊀ZHANGBao ̄long1ꎬ2(1.CollegeofTropicalCrops/SanyaNanfanResearchInstituteꎬHainanUniversityꎬHaikou570228ꎬChinaꎻ2.InstituteofGermplasmResourcesandBio ̄technology/ProvincialKeyLaboratoryofAgrobiologyꎬJiangsuAcademyofAgriculturalSciencesꎬNanjing210014ꎬChina)㊀㊀Abstract:㊀Cultivatingnon ̄transgenicherbicide ̄resistantricegermplasmresourcesisofgreatvalueforweedcontrolinricefields.InthisstudyꎬZhennuo19riceseedsweremutagenizedbyethylmethylsulfonate(EMS)solutionꎬandaM3generationofriceseedlingswithstableinheritanceandtolerancetoacetyl ̄CoAcarboxylase(ACCase)inhibitorherbicideswereobtained.ThegenomicDNAsofwild ̄typeandthemutantwereamplifiedandsequencedrespectively.Itwasfoundthattherewasapointmutationatthe5374thbaseoftheopenreadingframeoftheresistantriceACCasegeneꎬresultinginamutationoftheencoded1792thaminoacidfromisoleucinetoleucine.ThreekindsofACCaseinhibitorherbicidesweresprayedinthefieldandtheagronomictraitswereanalyzed.Theresultsshowedthattheresistanceofthemutanttohaloxy ̄fop ̄R ̄methylꎬquizalofop ̄P ̄ethylandpinoxadenwassignificantlyhigherthanthatofwildtype.Inthisstudyꎬanewnon ̄transgenicricegermplasmwithACCaseinhibitorresistancewasobtainedꎬwhichhadcertainapplicationvalueandcouldprovidegermplasmresourcesforherbicide ̄resistantricebreeding.Keywords:㊀riceꎻherbicide ̄resistantgermplasmꎻethylmethylsulfonate(EMS)ꎻacetylCoAcarboxylase503㊀㊀水稻是中国三大粮食作物之一ꎬ培育高产稳产的优质水稻是解决粮食问题的关键ꎮ稻田杂草严重影响水稻的产量和品质ꎬ杂草导致中国稻谷每年亏损率超过15%ꎬ部分地区甚至超过50%[1]ꎮ化学除草是当今世界使用最多的稻田除草方法ꎮ然而ꎬ过度使用除草剂不仅会导致杂草对除草剂产生抗性ꎬ还会对作物产生药害㊁降低水稻产量和品质ꎬ严重时甚至造成水稻颗粒无收[2]ꎮ因此ꎬ培育抗除草剂的水稻品种可以经济有效地解决稻田的杂草防除问题ꎮ乙酰辅酶A羧化酶(ACCase)是植物初级代谢中脂肪酸合成的关键酶之一ꎬ其主要功能是将乙酰辅酶A羧化为丙二酰辅酶Aꎮ该反应是脂肪酸合成的第一步ꎬ也是限速的关键步骤[3]ꎮ脂肪酸不仅是功能物质甘油三脂的组成成分ꎬ还能转化为作为细胞膜组成成分的磷脂[4]ꎮ自1958年发现乙酰辅酶A羧化酶可作为除草剂的作用靶标后ꎬ针对该靶标已开发了三大类除草剂并商品化应用ꎬ分别是芳氧苯氧基丙酸酯类(APP)[5]㊁环己烯酮类(CHD)[6]和新苯基吡唑啉类(DEN)[7 ̄8]ꎮ其中ꎬAPP类除草剂包括高效氟吡甲禾灵(Haloxyfop ̄R ̄methylꎬ又称高效盖草能)㊁精喹禾灵(Quizalofop ̄P ̄ethyl)㊁精恶唑禾草灵(Fenoxaprop ̄P ̄ethylꎬ又称骠马)㊁恶唑酰草胺(Metamifop)和氰氟草酯(Cyhalofop ̄butyl)等ꎮCHD类除草剂包括烯禾啶(Sethoxydim)㊁噻草酮(Cy ̄cloxydim)和环苯草酮(Profoxydim)等ꎻDEN类除草剂有唑啉草酯(Pinoxaden)ꎮ乙酰辅酶A羧化酶抑制剂类除草剂主要被用于控制禾本科杂草ꎬ具有高效㊁低毒㊁对后茬作物安全等特点[9]ꎮ目前ꎬ水稻生产中登记并许可使用的乙酰辅酶A羧化酶抑制剂类除草剂仅有氰氟草酯㊁恶唑酰草胺和环苯草酮ꎬ这极大限制了水稻生产中杂草的防治ꎮ因此培育抗乙酰辅酶A羧化酶抑制剂类除草剂水稻ꎬ不仅可以拓宽稻田除草剂的选择和使用范围ꎬ还可有效控制稻田杂草的发生与危害ꎮ化学诱变是培育和筛选抗性除草剂作物种质资源的重要方法ꎮEMS是非常有效且负面影响小的化学诱变剂ꎬ被广泛应用于构建优良性状的水稻突变体[10 ̄12]ꎮ顾佳清等利用EMS处理粳稻品种中花11ꎬ从诱变的水稻群体中筛选出高产的突变体ꎬ经过后代的纯化ꎬ得到了一个可以直接推广应用的水稻突变新品系申化一号[13]ꎮ陈忠明等通过EMS处理籼稻9311ꎬ筛选出了大粒的突变体M316和长穗突变体9311eR[14 ̄15]ꎮ本课题组用EMS诱变处理包括9311在内的多个水稻品种ꎬ成功筛选到多个抗咪唑啉酮类除草剂的突变体ꎬ进一步鉴定结果表明突变均发生在编码乙酰乳酸合成酶(ALS)靶标基因上[16]ꎮ本研究通过EMS诱变糯稻品种镇糯19构建突变群体ꎬ用APP类除草剂高效盖草能去筛选诱变处理后的M2代幼苗ꎬ获得能稳定遗传的抗性植株ꎬ并对抗性植株的ACCase基因位点突变㊁氨基酸序列变异进行鉴定ꎬ最后就3种不同ACCase抑制剂类除草剂对获得的抗除草剂材料农艺性状影响进行分析ꎬ旨在为水稻抗除草剂育种提供依据和材料ꎮ1㊀材料与方法1.1㊀材料与试剂供试水稻材料镇糯19由江苏丘陵地区镇江农业科学研究所提供ꎮ试验所用除草剂的种类及相关信息见表1ꎮ表1㊀本试验所用除草剂Table1㊀Herbicidesusedinthisstudy名称㊀类别来源推荐田间施用剂量(g/hm2ꎬa.i.)高效盖草能APP江苏中旗科技股份有限公司64.8精喹禾灵APP天津中农立华农用化学品有限公司60.0唑啉草酯DEN瑞士先正达作物保护有限公司45.0㊀㊀生物试剂甲基磺酸乙酯(EMS)购自美国Sigma ̄Aldrich公司ꎬ2ˑRapidTaqMasterMix㊁PhantaMaxSuper ̄FidelityDNAPolymerase聚合酶购自南京诺唯赞生物科技有限公司ꎬCTAB购自北京鼎国昌盛生物技术有限责任公司ꎮ1.2㊀镇糯19水稻种子的EMS诱变及抗ACCase抑制剂类除草剂突变体的筛选㊀㊀镇糯19种子(M1代)清水浸泡2h后ꎬ用质量浓度5 0mg/ml的EMS水溶液浸种处理14hꎬ硫代硫酸钠中和30min后ꎬ将种子捞出并用清水冲洗5~6遍ꎮ将诱变处理后的种子播种于大田ꎬM1代植株成熟后ꎬ种子混收(M2代)作为突变群体库ꎮ从突变群体库中取M2代种子播种于大棚苗床ꎬ待水稻幼苗长至3~4叶期时喷施64 8g/hm2ꎬa.i.高效盖草603江苏农业学报㊀2023年第39卷第2期能ꎬ施药后21d观察记录水稻表型并将正常生长的水稻苗移栽至盆钵内ꎬ单株收获种子得到突变体种子(M3代)ꎬM3代种子播种后得到M3代幼苗ꎮ1.3㊀抗除草剂突变体ACCase基因的PCR鉴定和碱基序列分析㊀㊀从国家水稻数据中心数据库(https://www.rice ̄data.cn/)获得水稻ACCase基因(OsACCꎬ序列号为LOC_Os05g22940)的碱基序列ꎮ根据OsACC基因的保守序列使用SnapGene6.0.2软件进行特异性引物设计ꎬ共设计了8对引物ꎬ分别是OsACC ̄F1~Os ̄ACC ̄F8和OsACC ̄R1~OsACC ̄R8(表2)ꎮ表2㊀本试验所用引物Table2㊀Primersusedinthisstudy引物名称㊀序列(5ᶄң3ᶄ)PCR产物长度(bp)OsACC ̄F1GTCAGATTTCACACATCTGGG1422OsACC ̄R1CAGGGGCACAAATAATGTACTOsACC ̄F2AAAAAGCTGCGTGAAGTATGC1614OsACC ̄R2TCTCGACTGTGAAGTGCTGCOsACC ̄F3CCCTATTGAAGACATCCTGATTG1597OsACC ̄R3AACAGAAATGGCATGATGGAOsACC ̄F4CAAACGTAGACTACACAGTTGAC1641OsACC ̄R4TGTTTGGCACCATTATGAGAAOsACC ̄F5TTGACAAGGTAAACATCATGTCC1635OsACC ̄R5AAAAGGTCATTGAAAAATTCACGOsACC ̄F6TCTATCCAAATCCTGCTGCC1631OsACC ̄R6AATGGCCAGTTCTAATTGCGOsACC ̄F7AGTTTTCTTCGGGCCAGATT1634OsACC ̄R7GGCTGGTCAAGACGCTGTATOsACC ̄F8CATGGAAGTGCTGCTATTGCCAG1866OsACC ̄R8CAGACTTGCACTTTCATCTGGCA㊀㊀采用CTAB法[14]提取水稻的基因组DNAꎬ取M3代三叶一心期的叶片0 5g放在带有1颗小钢珠的2ml离心管中ꎬ放到液氮中冷冻至叶片组织变脆ꎬ再将离心管放到频率为60Hz的组织研磨机研磨2minꎬ然后加入400μlCTAB提取液ꎬ离心管65ħ水浴30min后ꎬ在通风橱中加入400μl氯仿ꎬ充分混匀至提取液呈乳绿色ꎬ12000r/min离心10minꎬ在离心期间标记好管号ꎬ将600μl无水乙醇加入到已经标记好的1 5ml离心管中ꎬ移液枪吸取上清液300μl加到已经准备好的离心管中ꎬ上下颠倒混匀再沉淀1h以上ꎬ12000r/min离心10minꎬ倒掉上清液ꎬ开盖ꎬ室温下风干12h至离心管底部有明显的白色DNA沉淀ꎬ风干后加入灭菌蒸馏水200μlꎬ于-20ħ保存ꎮ以M3代的基因组DNA为模板ꎬ采用2ˑRapidTaqMasterMix或PhantaMaxSuper ̄FidelityDNAPolymerase聚合酶扩增OsACC基因的8个片段ꎮ用1%琼脂糖凝胶进行电泳检测ꎮ将条带大小正确的PCR产物送南京擎科生物科技有限公司进行测序ꎻ使用SnapGene6.0.2软件分析测序结果ꎬ明确野生型和突变体的OsACC基因碱基序列差异性ꎮ1.4㊀喷施除草剂后水稻农艺性状调查2022年在江苏省农业科学院试验基地进行镇糯19野生型和突变株系对3种乙酰辅酶A羧化酶抑制剂类除草剂耐受性试验ꎮ5月中旬播种ꎬ6月中旬插秧ꎮ试验设分别喷施高效盖草能㊁精喹禾灵㊁唑啉草酯及清水对照4个处理ꎬ每处理2.5mˑ4 0mꎮ移栽行距为0 25mꎬ株距为0 15mꎮ按照常规大田生产进行浇水和施肥等田间管理ꎮ镇糯19野生型和突变体幼苗移栽大田27d后ꎬ进行高效盖草能㊁精喹禾灵㊁唑啉草酯及清水(CK)的喷施处理ꎮ除草剂的用量见表1ꎮ各处理选择连续的20株ꎬ在水稻喷施除草剂前以及喷施除草剂后30d㊁90d进行茎蘖数㊁株高㊁主茎旗叶长度等农艺性状调查ꎮ其中ꎬ喷药后90dꎬ水稻已进入成熟期ꎬ统计的茎蘖数为成穗数ꎮ1.5㊀数据处理与统计分析采用MicrosoftExcel2019进行数据处理ꎬ用GraphPadPrism8.0.1软件进行统计分析ꎮ2㊀结果与分析2.1㊀抗除草剂突变体筛选高效盖草能是一种内吸传导型除草剂ꎬEMS诱变的镇糯19M2代水稻幼苗在3~4叶期喷施高效盖草能7d后ꎬ绝大部分水稻幼苗叶片颜色变成浅绿ꎻ喷施高效盖草能21d后ꎬ敏感植株叶片几乎完全失去绿色㊁部分已经枯死ꎻ具有抗性的植株能继续正常生长ꎮ经大量筛选后ꎬ最终获得1株具有高效盖草能抗性的M2单株(图1)ꎬ成熟后收获单株种子ꎬ得到M3代抗性突变体ꎮ2.2㊀OsACC突变位点已知高效盖草能的作用靶标是ACCaseꎬ植物对703江㊀群等:EMS诱变创制水稻抗乙酰辅酶A羧化酶抑制剂类除草剂种质图1㊀喷施高效盖草能后筛选到的M2代抗性水稻植株Fig.1㊀M2generationresistantriceplantscreenedafterspra ̄yingwith64.8ga.i./hm2haloxyfop ̄R ̄methyl高效盖草能的抗性主要源于ACCase基因的突变[17 ̄19]ꎮ为了确定突变体中靶标基因是否发生突变ꎬ我们用了8对引物对野生型(镇糯19 ̄WT)和抗性M3单株(镇糯19 ̄1792)的基因进行扩增ꎬ全部都获得了与预期大小相符合的条带(图2)ꎮ㊀㊀上述PCR扩增的产物经测序和碱基序列比对ꎬ发现相对于野生型OsACC的ORFꎬ突变体OsACC基因中存在一个点突变ꎬ其开放阅读框(ORF)的第5374位碱基由A突变成Tꎬ从而引起编码的第1792位氨基酸由异亮氨酸(Ile)突变为亮氨酸(Leu)(图3A)ꎮOsACC蛋白的全长有2327个氨基酸ꎬ将Os ̄ACC蛋白全长氨基酸序列在NCBI的ConservedDo ̄main数据库(https://www.ncbi.nlm.nih.gov/Struc ̄ture/cdd/wrpsb.cgi)进行保守结构域分析ꎬ发现其包含了4个结构域(Domain):生物素羧化酶(BC)㊁生物素羧基载体蛋白(BCCP)㊁乙酰辅酶A羧化酶中心(ACCcentral)和羧基转移酶(CT)(图3B)ꎮ进一步的氨基酸序列分析结果表明ꎬ突变体中第1792位氨基酸的突变位于CT结构域ꎬ该突变类型与已报道的大穗看麦娘(Alopecurusmyosuroides)的抗性位点突变类型是一致的ꎬ对应于其ACCase氨基酸序列第1781位点ꎻ突变类型也相同ꎬ均由Ile突变为Leu(图3B和3C)[17]ꎮ因此ꎬ突变体抗除草剂功能的获得是由OsACC氨基酸序列第1792位氨基酸由异亮氨酸突变为亮氨酸引起的ꎮ2.3㊀突变体的农艺性状在分别喷施高效盖草能㊁精喹禾灵和唑啉草酯14d后ꎬ野生型植株生长均受到了显著影响ꎬ大部分植株叶片出现枯黄症状ꎮ突变体植株在分别喷施以上3种除草剂后ꎬ叶片仍然是绿色且可以正常生长ꎬ表明突变体对这3种除草剂均具有抗性(图4)ꎮ㊀㊀分蘖期分别喷施3种不同除草剂后ꎬ野生型和突变体株高㊁分蘖数及旗叶长度的变化如图5所示ꎮ结果显示ꎬ在喷施清水处理的情况下ꎬ野生型和突变体植株的株高在处理前(0dꎬ即幼苗移栽到大田27d)基本没有差异ꎬ但在处理后30d和90dꎬ突变体的株高显著低于野生型的株高(图5A)ꎻ两者在处理前㊁后的单株茎蘖数均无明显差异(图5E)ꎮ在分别喷施3种不同除草剂前(0d)ꎬ野生型和突变体植株的株高和单株分蘖数都没有明显差异ꎬ但是在喷施处理后ꎬ两者受除草剂的影响表现出明显差异(图5B~图5D㊁图5F~图5H)ꎮ其中ꎬ在喷施高效盖草能30d和90d后ꎬ突变体的株高均显著高于野生型(图5B)ꎬ单株茎蘖数也显著多于野生型(图5F)ꎮ野生型对精喹禾灵和唑啉草酯都非常敏感ꎬ喷施田间推荐剂量后水稻植株均死亡ꎬ因此未统计喷药后的株高和分蘖数ꎬ而突变体对这两种除草剂表现出较强的抗性ꎬ所有植株存活且能正常生长ꎬ株高随时间逐渐增加(图5C和5D)ꎮ突变体的单株茎蘖数在精喹禾灵处理后随时间呈先增后减趋势ꎬ但经唑啉草酯处理后变化不明显ꎬ未出现明显增加现象(图5G和5H)ꎮ喷施清水处理的突变体旗叶长度显著短于野生型ꎻ高效盖草能处理后ꎬ突变体的旗叶长度显著长于野生型(图5I)ꎮ由于野生型在喷施田间推荐剂量的精喹禾灵和唑啉草酯后植株已经枯死ꎬ因此未能进行旗叶长度统计ꎮ综合以上结果ꎬ在田间推荐剂量下ꎬ突变体对高效盖草能㊁精喹禾灵和唑啉草酯的抗性水平均高于野生型ꎮ3㊀讨论植物对除草剂的抗性机制包括非靶标和靶标抗性两大类ꎮ其中ꎬ非靶标抗性是由靶标基因以外的突变引起的ꎬ使植物对除草剂的吸收或转运率降低㊁螯合或代谢作用增强ꎻ靶标抗性是由除草剂的靶标基因发生突变引起的[20]ꎮ现在已发现的大部分植物抗ACCase抑制剂类除草剂的抗性机制是由于ACCase基因碱基突变引起氨基酸位点发生变异ꎬ这也是导致杂草抗药性产生的主要原因[21 ̄22]ꎮ截止803江苏农业学报㊀2023年第39卷第2期M表示DNAmarkerꎻ泳道1表示野生型ꎻ泳道2表示突变体ꎮF1~F8㊁R1~R8为引物ꎬ见表2ꎮ图2㊀镇糯19野生型和突变体中OsACC基因的PCR扩增结果Fig.2㊀PCRamplificationofOsACCinZhennuo19wild ̄typeandmutantA:突变体(镇糯19 ̄1792)中OsACC基因的Sanger测序色谱图ꎻB:OsACC蛋白结构域示意图ꎻC:野生型(镇糯19 ̄WT)和突变体(镇糯19 ̄1792)的羧基转移酶(CT)结构域氨基酸序列比对ꎮ图3㊀镇糯19突变体中突变基因OsACC及其编码氨基酸序列分析Fig.3㊀AnalysisofmutantgeneOsACCanditsencodedaminoacidsequenceinZhennuo19mutant903江㊀群等:EMS诱变创制水稻抗乙酰辅酶A羧化酶抑制剂类除草剂种质镇糯19 ̄WT㊁镇糯19 ̄1792分别表示镇糯19野生型和突变体ꎻGCN㊁JK㊁ZL和H2O分别表示喷施高效盖草能㊁精喹禾灵㊁唑啉草酯及清水处理ꎮ图4㊀镇糯19野生型和突变体田间喷施不同除草剂后的表型Fig.4㊀PhenotypesofZhennuo19wild ̄typeandmutantaftersprayingwithdifferentherbicidesinthefield目前ꎬ杂草中已报道了十几种ACCase氨基酸置换与其抗药性相关ꎬ分别对应于大穗看麦娘ACCase的7个氨基酸位点(均位于CT结构域内):第1781位㊁第1999位㊁第2027位㊁第2041位㊁第2078位㊁第2088位和第2096位[22 ̄25]ꎮ在以上这些突变中ꎬ以第1781位氨基酸由Leu突变成Ile最为普遍ꎬ对三大类不同的ACCase抑制剂类除草剂都表现出高抗性ꎬ却没有适合度代价(Fitnesscost)[26 ̄28]ꎮ本研究通过筛选EMS诱变的镇糯19水稻突变体ꎬ鉴定到了1个能稳定遗传的抗除草剂突变体ꎮ对突变体进行了基因鉴定ꎬ确定其编码靶标蛋白OsACC的第1792位氨基酸由Leu突变成Ileꎮ该突变类型与已报道的突变类型一致ꎬ对应于大穗看麦娘ACCase第1781位氨基酸突变ꎮ这是该突变类型使水稻获得多种ACCase抑制剂类除草剂抗性的首次报道ꎮEMS是最常见的化学诱变剂ꎬ在植物的诱变育种中被广泛应用[29]ꎮ本试验通过EMS诱变镇糯19种子ꎬ筛选到了抗ACCase抑制剂类除草剂的水稻植株ꎬ突变体能耐受田间推荐剂量的高效盖草能㊁精喹禾灵和唑啉草酯ꎮ其中ꎬ喷施了田间推荐剂量的唑啉草酯后ꎬ镇糯19野生型植株在处理30d后几乎全部死亡ꎻ喷施了田间推荐剂量的精喹禾灵后ꎬ野生型的植株在喷施30d后全部死亡ꎻ而突变体在分别喷施3种除草剂后ꎬ均未出现死亡现象ꎬ基本可以正常生长ꎮ所获得的抗性突变体对高效盖草能㊁精喹禾灵㊁唑啉草酯的抗性水平均明显强于野生型ꎮ突变体和野生型的最小致死剂量或50%抑制浓度(GR50)㊁OsACC酶活性的差异尚有待进一步明确ꎮ大豆㊁棉花和玉米等转基因作物已在全球范围内进行了商品化生产ꎬ产生了巨大的社会效益和经济效益ꎮ目前为止ꎬ中国虽然有多种转基因作物已经被正式批准商品化生产ꎬ但进行大面积种植的仅013江苏农业学报㊀2023年第39卷第2期H2O㊁GCN㊁JK㊁ZL分别表示喷施清水㊁高效盖草能㊁精喹禾灵㊁ꎬ∗∗表示在0.01水平上极显著ꎮND表示没有数据ꎬns表示没有显著差异ꎮ图5㊀不同除草剂处理下的水稻株高㊁分蘖数和旗叶长度Fig.5㊀Plantheightꎬtillernumberandflagleaflengthofriceunderdifferentherbicidetreatments有番木瓜和棉花ꎮ2009年ꎬ农业部颁发了中国拥有自主知识产权的转Bt基因抗虫水稻生产应用安全证书ꎬ但目前中国尚未批准转基因水稻的商业化生产ꎮ因此ꎬ培育非转基因的抗除草剂水稻品种具有重要价值ꎮ上世纪90年代晚期ꎬ美国路易斯安那州州立大学稻米研究中心通过EMS诱变技术育成了一系列耐咪唑啉酮类除草剂(ALS抑制剂类除草剂)的非转基因水稻品种ꎮ2002年ꎬ巴斯夫公司开发了非转基因抗咪唑啉酮类除草剂的水稻品种Clearf ̄ield在美国进行了商业化推广ꎬ解决了水稻种植的杂草稻危害问题[30]ꎮ2018年ꎬ巴斯夫又在美国上市了非转基因水稻品种Provisiaꎬ可以抗精喹禾灵ꎬ拟与抗咪唑啉酮类除草剂水稻品种Clearfield进行轮作并交替使用两种不同作用机理的除草剂ꎬ实现对杂草稻和其他一年生杂草的可持续性防控[31]ꎮ本研究通过EMS诱变筛选到的抗ACCase抑制剂类除草剂突变体ꎬ具有与抗除草剂精喹禾灵水稻品种Provisia类似的抗除草剂性状ꎬ可为中国非转基因抗除草剂水稻育种提供重要材料ꎮ4㊀结论本研究通过EMS诱变筛选获得了可稳定遗传的抗ACCase抑制剂类除草剂的水稻突变体材料ꎬ可耐受3种不同田间推荐剂量的除草剂ꎬ具有一定的生产应用价值ꎮ野生型在喷施田间推荐剂量的高效盖草能㊁精喹禾灵㊁唑啉草酯后ꎬ株高和分蘖均受到严重抑制甚至死亡ꎬ但突变体基本能正常生长ꎮ突变体中OsACC突变基因编码蛋白质的第1792位氨基酸由Ile变成Leuꎬ使其对ACCase抑制剂类除草剂的耐受性显著提高ꎮ在当前中国转基因水稻尚未放开㊁公众对转基因作物品种存在疑虑的大背景下ꎬ本研究获得的非转基因抗除草剂材料具有良好的应用前景ꎮ113江㊀群等:EMS诱变创制水稻抗乙酰辅酶A羧化酶抑制剂类除草剂种质参考文献:[1]㊀董立尧ꎬ高㊀原ꎬ房加鹏ꎬ等.我国水稻田杂草抗药性研究进展[J].植物保护ꎬ2018ꎬ44(5):69 ̄76.[2]㊀程艳勤.浅析除草剂对水稻的危害及治理[J].农技服务ꎬ2016ꎬ33(6):109 ̄114.[3]㊀KONISHITKUJꎬSHINOHARAKꎬYAMADAKꎬetal.Acetyl ̄CoAcarboxylaseinhigherplants:mostplantsotherthangramineaehaveboththeprokaryoticandtheeukaryoticformsofthisenzyme[J].PlantandCellPhysiologyꎬ1996ꎬ37(2):117 ̄122. [4]㊀王㊀爽ꎬ张荣全ꎬ叶㊀非.乙酰辅酶A羧化酶抑制剂的研究进展[J].农药科学与管理ꎬ2003(10):26 ̄32.[5]㊀蔡靖萱.扬州市小麦田菵草对ACCase抑制剂的抗性研究[D].扬州:扬州大学ꎬ2020.[6]㊀RENDINAARꎬFELTSJM.CyclohexanedioneHerbicidesarese ̄lectiveandpotentinhibitorsofacetyl ̄CoAcarboxylasefromgrasses[J].PlantPhysiolꎬ1988ꎬ86(4):983 ̄986.[7]㊀袁国徽ꎬ王恒智ꎬ赵㊀宁ꎬ等.耿氏硬草对乙酰辅酶A羧化酶类除草剂抗性水平及分子机制初探[J].农药学学报ꎬ2016ꎬ18(3):304 ̄310.[8]㊀董元海.新苯基吡唑啉类除草剂唑啉草酯的合成[D].武汉:武汉工程大学ꎬ2017.[9]㊀刘博宏ꎬ叶㊀非.芳氧苯氧基丙酸酯类除草剂的应用进展[J].农药科学与管理ꎬ2011ꎬ32(2):20 ̄25.[10]吕㊀军ꎬ刘㊀军ꎬ姜秀英ꎬ等.EMS诱导水稻 辽星1号ᶄ突变体的筛选与鉴定[J].分子植物育种ꎬ2022ꎬ20(12):4038 ̄4043. [11]董颖苹ꎬ连㊀勇ꎬ何庆才ꎬ等.植物化学诱变技术在育种中的运用及进展Ⅱ.突变体的筛选及分子检测[J].种子ꎬ2005ꎬ24(8):54 ̄58.[12]黄㊀静.水稻EMS诱变效率和品种内遗传多态性分析[D].福州:福建农林大学ꎬ2015.[13]顾佳清ꎬ张智奇ꎬ周㊀音ꎬ等.EMS诱导水稻中花11突变体的筛选和鉴定[J].上海农业学报ꎬ2005ꎬ21(1):7 ̄11.[14]陈忠明ꎬ王秀娥.水稻强优势恢复系9311粒重的诱变改良[J].分子植物育种ꎬ2005ꎬ3(3):353 ̄356.[15]陈忠明ꎬ王秀娥ꎬ胡兴雨ꎬ等.水稻长穗颈恢复系9311eR的诱变选育[J].江苏农业科学ꎬ2005(4):9 ̄11.[16]陈天子ꎬ余㊀月ꎬ凌溪铁ꎬ等.EMS诱变水稻创制抗咪唑啉酮除草剂种质[J].核农学报ꎬ2021ꎬ35(2):253 ̄261.[17]DÉLYECꎬCALMÈSÉꎬMATÉJICEKA.SNPmarkersforblack ̄grass(AlopecurusmyosuroidesHuds.)genotypesresistanttoacetylCoA ̄carboxylaseinhibitingherbicides[J].TheoreticalandAppliedGeneticsꎬ2002ꎬ104(6):1114 ̄1120.[18]DÉLYECꎬZHANGXꎬCHALOPINCꎬetal.Anisoleucineresi ̄duewithinthecarboxyl ̄transferasedomainofmultidomainacetyl ̄coenzymeAcarboxylaseisamajordeterminantofsensitivitytoary ̄loxyphenoxypropionatebutnottocyclohexanedioneinhibitors[J].PlantPhysiologyꎬ2003ꎬ132(3):1716 ̄1723.[19]DÉLYECꎬZHANGXꎬMICHELSꎬetal.Molecularbasesforsensitivitytoacetyl ̄coenzymeAcarboxylaseinhibitorsinblack ̄grass[J].PlantPhysiologyꎬ2005ꎬ137(3):794 ̄806.[20]POWLESSBꎬYUQ.EvolutioninAction:plantsresistanttoher ̄bicides[J].AnnualReviewofPlantBiologyꎬ2010ꎬ61(1):317 ̄347.[21]袁国徽ꎬ田志慧ꎬ高㊀原ꎬ等.上海市水稻田千金子对3种乙酰辅酶A羧化酶抑制剂的抗性现状及酶突变机制[J].农药学学报ꎬ2022ꎬ24(3):492 ̄500.[22]BECKIEHJꎬTARDIFFJ.Herbicidecrossresistanceinweeds[J].CropProtectionꎬ2012ꎬ35:15 ̄28.[23]DENGWꎬCAIJꎬZHANGJꎬetal.MolecularbasisofresistancetoACCase ̄inhibitingherbicidecyhalofop ̄butylinChinesespran ̄gletop(Leptochloachinensis(L.)Nees)fromChina[J].PesticBiochemPhysiolꎬ2019ꎬ158:143 ̄148.[24]PENGYꎬPANLꎬLIUDꎬetal.Confirmationandcharacterizationofcyhalofop ̄butyl ̄resistantChinesesprangletop(Leptochloachinensis)populationsfromChina[J].WeedScienceꎬ2020ꎬ68(3):253 ̄259.[25]张㊀怡ꎬ陈丽萍ꎬ徐笔奇ꎬ等.浙江稻区千金子对氰氟草酯和噁唑酰草胺的抗药性及其分子机制研究[J].农药学学报ꎬ2020ꎬ22(3):447 ̄453.[26]VILAAIUBMMꎬNEVEPꎬPOWLESSB.ResistancecostofacytochromeP450herbicidemetabolismmechanismbutnotanAC ̄CasetargetsitemutationinamultipleresistantLoliumrigidumpopulation[J].NewPhytologistꎬ2005ꎬ167(3):787 ̄796. [27]MENCHARIYꎬCHAUVELBꎬDARMENCYHꎬetal.Fitnesscostsassociatedwiththreemutantacetyl ̄coenzymeAcarboxylaseallelesendowingherbicideresistanceinblack ̄grassAlopecurusmy ̄osuroides[J].JournalofAppliedEcologyꎬ2008ꎬ45(3):939 ̄947. [28]WANGTꎬPICARDJCꎬTIANXꎬetal.Aherbicide ̄resistantACCase1781setariamutantshowshigherfitnessthanwildtype[J].Heredity(Edinb)ꎬ2010ꎬ105(4):394 ̄400.[29]SERRATXꎬESTEBANRꎬGUIBOURTNꎬetal.EMSmutagene ̄sisinmatureseed ̄derivedricecalliasanewmethodforrapidlyobtainingTILLINGmutantpopulations[J].PlantMethodsꎬ2014ꎬ10(1):5.[30]SHAXYꎬLINSCOMBESDꎬGROTHDE.Fieldevaluationofimidazolinone ̄tolerantClearfieldrice(OryzasativaL.)atnineLouisianalocations[J].CropScienceꎬ2007ꎬ47(3):1177 ̄1185. [31]CAMACHOJRꎬLINSCOMBESDꎬSANABRIAYꎬetal.Inherit ̄anceofProvisiariceresistancetoquizalofop ̄p ̄ethylunderlabora ̄toryandgreenhouseenvironments[J].Euphyticaꎬ2019ꎬ215(4):83.(责任编辑:石春林)213江苏农业学报㊀2023年第39卷第2期。
关键酶名词解释

关键酶名词解释关键酶是指在细胞代谢中发挥关键作用的酶。
酶(Enzyme)是一种催化生物化学反应的蛋白质,由于酶的催化作用极具特异性和高效性,因此酶在细胞代谢中起着至关重要的作用。
关键酶则是对维持生命特别重要的酶,其催化作用对于细胞的正常生存和功能发挥至关重要。
下面将对几种常见的关键酶进行详细解释。
1. 乙酰辅酶A羧化酶(Acetyl-CoA carboxylase):这是一种关键的酶,在细胞内催化乙酰辅酶A转化为丙酰辅酶A。
丙酰辅酶A是细胞合成脂肪酸和胆固醇的重要物质。
乙酰辅酶A羧化酶的活性直接影响脂肪酸和胆固醇的合成速率,调节细胞内脂肪代谢的平衡。
2. 乙醛脱氢酶(Alcohol dehydrogenase):这是一类关键酶,它催化乙醇转化为乙醛。
乙醛脱氢酶参与乙醇代谢的过程,将乙醇转化为乙醛,进而进一步转化为乙酸。
这个过程是人体中乙醇的主要代谢途径,也是酒精中毒的解毒过程的一部分。
3. DNA聚合酶(DNA polymerase):DNA聚合酶是细胞中复制和修复DNA过程中的关键酶类。
DNA聚合酶能够将DNA 模板链上的碱基序列准确地复制到合成链中,是DNA复制的关键酶。
细胞复制DNA时,DNA聚合酶能够保证复制过程中的准确性,从而确保新合成的DNA与原始模板DNA完全一致。
4. 丙酸脱氢酶(Pyruvate dehydrogenase):丙酸脱氢酶是关键的酶类之一,参与细胞呼吸过程中的关键环节。
丙酸脱氢酶能够将丙酮酸转化为乙酰辅酶A,为细胞供应继续产生能量所需的底物。
丙酸脱氢酶活性的调节与细胞的能量代谢紧密相关。
5. ATP酶(ATPase):ATP酶是将ATP(腺苷三磷酸)分解为ADP(腺苷二磷酸)和无机磷酸盐的关键酶。
细胞中的能量通常储存于ATP中,而ATP酶能够将ATP分解为ADP释放出储存的能量。
这个过程是细胞内能量供应的重要途径,也是调节细胞内ATP/ADP比例的重要手段。
以上仅为几个常见的关键酶的解释,关键酶的种类很多,每个关键酶都在重要的生物代谢过程中扮演着重要的角色。
乙酰辅酶a羧化酶系名词解释
乙酰辅酶a羧化酶系名词解释胞内or,胞外:是指在细胞膜以外。
例如,细胞膜外可能还有很多很小的内膜系统,其内不一定都有大分子的酶类。
此时可根据所知的底物的性质来确定它是否参与跨膜转运过程。
底物:即生物体内合成的某种化合物,能够被专一酶催化而转变成另一种物质。
此时我们说底物一般是指各种有机物。
酶:催化生物化学反应的特殊蛋白质。
蛋白质的一级结构(氨基酸顺序)决定了酶的活性中心,而其活性中心的空间结构则决定了酶的专一性。
肝胆胞外:分布于血液、肾脏、皮肤、肠道等的毛细血管中。
肝细胞外,在高尔基体的胞浆中也含有很多的线粒体,线粒体外围也含有很多的粗面内质网,有很多的电子传递链,电子流动的速度非常快,线粒体中的叶绿体进行光合作用。
胞内or,细胞器中:由于线粒体内很多区域的功能未知或研究尚少,因此这些区域被归为胞内or的部分。
比如,中心体、内质网、核糖体等。
细胞器:由膜围起来的细胞内部结构。
细胞器的作用:细胞器具有特定的结构和功能,并在发挥功能时与细胞内的其他部分协调配合。
细胞器通常包括细胞核、内质网、高尔基体、溶酶体、过氧化物酶体、线粒体、叶绿体、液泡等。
组成:辅酶A、乙酰辅酶A羧化酶、辅酶A脱氢酶、辅酶A脱氢酶脱氢酶:辅酶A脱氢酶是辅酶A和乙酰辅酶A脱氢形成丙酮酸的酶系,参与代谢和呼吸的调节。
乙酰辅酶A羧化酶:由三个亚基组成,有活性中心,结构和功能相似,但与辅酶A和乙酰辅酶A的反应部位和顺序不同。
酶原(Ct)基因:乙酰辅酶A羧化酶的基因。
转录本:转录单位以一定方式排列,保持一定顺序,并携带DNA序列的串联重复。
转录(translation)就是将转录本中的DNA序列翻译成蛋白质。
活性代谢产物:乙酰辅酶A羧化酶是从线粒体膜上的三羧酸循环受体接收丙酮酸,再将丙酮酸羧化成乙酰辅酶A,最后将乙酰辅酶A 还原成琥珀酰辅酶A,还原反应中脱下的氢以H结合形式供给三羧酸循环。
活性氧(reactive oxygen species, ROS)可以攻击细胞,造成正常细胞损伤,而且损伤持续时间长,危害大。
索莱宝 ABTS 自由基清除能力检测试剂盒说明书(可见分光光度法)
ABTS自由基清除能力检测试剂盒说明书可见分光光度法货号:BC4770规格:50T/24S产品内容:使用前请认真核对试剂体积与瓶内体积是否一致,有疑问请及时联系索莱宝工作人员。
试剂名称规格保存条件提取液液体60 mL×1瓶4℃保存试剂一液体80 mL×1瓶4℃保存试剂二粉剂×1瓶4℃保存试剂三液体20 μL×1支4℃保存试剂四液体1.5 mL×1支-20℃保存试剂五粉剂×1支4℃保存溶液的配制:1、试剂二:临用前加入3 mL蒸馏水,充分溶解;用不完的试剂可分装-20℃保存,-20℃可保存两周;2、试剂三工作液的配制:液体置于棕色试剂瓶中EP管内,临用前根据样本量按试剂三(μL):蒸馏水(mL)=1μL:12 mL的比例配成试剂三工作液,现用现配,用不完的试剂放于4℃保存;3、试剂四工作液的配制:可先将试剂四-20℃分装保存。
临用前根据样本量按试剂四:试剂一(V:V)=1:9的比例配成试剂四工作液,现配现用。
4、试剂五:粉剂置于试剂瓶内EP管中,含有5 mg维生素C。
临用前加入2.8 mL提取液,充分振荡溶解;配成10 mmol/L维生素C溶液,用于阳性对照。
4℃可保存一周。
5、ABTS工作液的配制:临用前根据样本量按试剂一:试剂二:试剂三工作液(V:V:V)=76:5:4的比例配成ABTS工作液,现用现配,室温避光保存,务必在30分钟内使用。
产品说明:ABTS法可用于亲水性和亲脂性物质抗氧化能力测定,是使用最广泛的间接检测方法。
ABTS经氧化后生成稳定的蓝绿色阳离子ABTS自由基,在405 nm或734 nm处有最大吸收峰。
被测物质加入ABTS自由基溶液后,所含抗氧化成分能与ABTS自由基发生反应而使反应体系褪色,405 nm的吸光度下降,在一定范围内其吸光度的变化与自由基被清除的程度成正比。
本试剂盒中,通过测定吸光度下降的程度来反映样本清除ABTS自由基的能力。
乙酰辅酶A羧化酶在治疗肥胖中的潜在作用
在维持整个机体能量 和 调 控 脂 肪 代 谢 中 发 挥 着 必 不可少的作用 ( 因 此 % 对 它 的 研 究 也 必 然 会 为 肥 胖病在治疗方法上提供新的手段 (
RETAEC , 中 明 确 指 出 肥 胖 作 为 一 种 疾 病 应 引 起 人
们的广泛关注 ( 肥 胖 在 医 学 上 被 定 义 为 体 重 % 尤 其是脂肪组织的过分 增 加 % 而 导 致 不 利 于 身 体 健 康的一种状况 ( 体 重 是 由 遗 传 ) 环 境 和 心 理 因 素 相互作用来控制的 ( 这 些 因 素 相 互 作 用 的 紊 乱 最 终会改变能量的平衡状态而导致体重的改变(
388: 年 % UJ@FG 等 首 先 用 定 位 克 隆 的 方 法 将 小 鼠 的 肥 胖 基 因 $ KVESE GEFE % KV 基 因 ’ 和 人 类 的 同 源
序列鉴定出来 ( 随后 肥 胖 基 因 的 表 达 产 物 瘦 蛋 白 $BEWNAF ’ 的 分 子 结 构 % 生 理 作 用 成 为 研 究 热 点 之 一 ( 肥胖基因的鉴 定 进 一 步 表 明 % 遗 传 因 子 在 调 控肥胖病中的重要作 用 ( 对 肥 胖 基 因 和 瘦 蛋 白 的 研究也为治疗肥胖病提供了新思路 ( 近年来科学研究发现 % 丙二酰辅酶 ; 在 能 量 代谢中起着重要的调 控 作 用 ( 它 不 仅 可 以 调 控 脂 肪的降解与合成 % 同 时 还 可 以 作 为 一 种 信 号 分 子 通过调节下丘脑来对摄食和能量平衡进行调控 X30!Y( 因此 % 催化丙二酰辅酶 ; 生成的乙酰辅酶 ; 羧化 酶必然在整个能量代谢 过 程 中 扮 演 着 关 键 的 调 控 角色 ( 对乙酰辅酶 ; 羧化酶的研究表明了它确实
当体内需要能量的时 候 % 胰 高 血 糖 素 和 肾 上 腺 素 使 6$$ 磷酸化导致其失活 # 禁食 ( 运动 ( 激素可 以 激 活 依 赖 腺 苷 酸 单 磷 酸 的 蛋 白 激 酶 & 6’=> ’ %
脂肪酸合成过程关键酶
脂肪酸合成过程关键酶脂肪酸是一类重要的生物分子,它们在机体内起着能源供应、结构组成和信号传导等多种生理功能。
而脂肪酸的合成过程则是细胞内进行的一系列化学反应所组成的。
在脂肪酸合成过程中,关键酶起着至关重要的作用,本文将围绕关键酶展开讨论。
在哺乳动物细胞内,脂肪酸的合成主要发生在细胞质内的胞浆液滴上,这个过程被称为脂肪酸的胞浆液滴合成。
关键酶主要包括乙酰辅酶A羧化酶(ACC)、甘油3磷酸脱氢酶(G3PDH)、丙酮酸羧化酶(ACLY)、乙酰辅酶A羧化酶(ACACA)等。
乙酰辅酶A羧化酶(ACC)是脂肪酸合成过程中的第一个关键酶。
它催化乙酰辅酶A转化为酯化的乙酰辅酶A(Malonyl-CoA),并且是脂肪酸合成的速率限制酶。
乙酰辅酶A是脂肪酸合成过程的起始物质,而酯化的乙酰辅酶A则是脂肪酸链的延长单位。
甘油3磷酸脱氢酶(G3PDH)是脂肪酸合成过程中的另一个关键酶。
它催化甘油3磷酸(G3P)转化为丙酮酸,为脂肪酸合成提供了碳源。
丙酮酸是脂肪酸合成的中间产物,可以进一步被转化为乙酰辅酶A,从而参与合成反应。
丙酮酸羧化酶(ACLY)也是脂肪酸合成过程中的关键酶。
它催化丙酮酸转化为乙酰辅酶A,并且是脂肪酸合成的速率限制酶。
乙酰辅酶A是脂肪酸链的延长单位,它与酯化的乙酰辅酶A一起参与脂肪酸链的合成。
乙酰辅酶A羧化酶(ACACA)是脂肪酸合成过程中的最后一个关键酶。
它催化酯化的乙酰辅酶A转化为酯化的丙酰辅酶A,从而完成脂肪酸链的延长。
乙酰辅酶A羧化酶的活性与合成脂肪酸的速率密切相关。
乙酰辅酶A羧化酶(ACC)、甘油3磷酸脱氢酶(G3PDH)、丙酮酸羧化酶(ACLY)、乙酰辅酶A羧化酶(ACACA)是脂肪酸合成过程中的关键酶。
它们分别参与了脂肪酸合成的不同步骤,协同作用,完成了脂肪酸链的合成。
这些关键酶的活性和调控对于维持脂肪酸的平衡和正常代谢具有重要作用,对于相关疾病的发生和进展也具有重要意义。
深入研究这些关键酶的功能和调控机制,将有助于进一步理解脂肪酸合成的生物学过程,为相关疾病的预防和治疗提供新的思路和策略。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
乙酰辅酶A羧化酶(ACC)活性检测试剂盒说明书可见分光光度法
注意:正式测定之前选择2-3个预期差异大的样本做预测定。
货号:BC0410
规格:50T/24S
产品简介:
乙酰辅酶A羧化酶(Acetyl-CoA carboxylase,ACC)在生物体内催化乙酰辅酶A羧化生成丙二酰辅酶A,是脂肪酸和许多次生代谢产物合成的关键酶。
ACC的活性在一定程度上决定了脂肪酸的合成速度和含油量的高低。
ACC能够催化乙酰辅酶A、NaHCO
和ATP生成丙二酰辅酶A、ADP和无机磷,钼蓝与磷酸根生成660nm
3
有特征吸收峰的物质,通过钼酸铵定磷法测定无机磷的增加量来测定ACC活性。
试验中所需的仪器和试剂:
可见分光光度计、天平、低温离心机、1mL玻璃比色皿、可调式移液枪、研钵/匀浆器、漩涡震荡仪、水浴锅、蒸馏水、冰盒、EP管。
产品内容:
提取液一:液体60mL×1瓶,4℃保存;
提取液二:液体0.6mL×1支,-20℃避光保存;
试剂一:液体50mL×1瓶,4℃保存;
试剂二:粉剂×1瓶,-20℃保存。
临用前加入12.5mL试剂一,充分溶解待用;可分装后-20℃保存,避免反复冻融;
试剂三:粉剂×1瓶,-20℃保存。
临用前加入6mL双蒸水,充分溶解待用;可分装后-20℃保存,避免反复冻融;
试剂四:粉剂×1瓶,4℃保存。
临用前加入15mL双蒸水,溶解后4℃保存一周;
试剂五:粉剂×1瓶,4℃保存。
临用前加入15mL双蒸水,溶解后4℃保存一周;
试剂六:液体15mL×1瓶,室温保存;
标准品:10μmol/mL标准磷贮备液1mL×1支,4℃保存;
提取液的配制:按提取液一:提取液二=990:10(V:V)的比例配制,根据样本量现配现用,禁止将提取液二一次性全部加入提取液一混匀分装待用。
O:试剂四:试剂五:试剂六=2:1:1:1的比例配制,配好的工作液应为工作液(定磷剂)的配制:按H
2
浅黄色。
若变色则试剂失效,若是蓝色则为磷污染,工作液应现配现用。
注意:配试剂最好用新的烧杯、玻璃棒和玻璃移液器,也可以用一次性塑料器皿,避免磷污染。
操作步骤:
一、粗酶液提取:
组织:按照组织质量(g):提取液体积(mL)为1:5~10的比例(建议称取约0.1g组织,加入1mL提取液)进行冰浴匀浆,然后,8000g,4℃,离心10min,取上清置于冰上待测。
细菌或细胞:收集细菌或细胞到离心管内,离心后弃上清;按照细菌或细胞数量(104个):按照细菌或细胞数量(104个):提取液体积(mL)为500~1000:1的比例(建议500万细胞加入1mL提取液),冰浴超声波破碎细胞(功率300w,超声3秒,间隔7秒,总时间3min);然后8000g,4℃,离心10min,取上清置于冰上待测。
血清(浆):直接测定。
二、测定步骤:
1、分光光度计预热30min以上,调节波长至660nm,蒸馏水调零。
2、将10μmol/mL标准液用蒸馏水稀释为1.25、0.625、0.3125、0.15625、0.078μmol/mL的标准溶液备
用。
3、操作表:(在1.5ml离心管中依次加入下列试剂)
酶促反应:
试剂名称对照管测定管
试剂一(µL)450
试剂二(µL)-250
试剂三(µL)-200
样本(µL)5050 37℃(哺乳动物)或25℃(其它物种)准确反应30min后,沸水浴5min(盖紧,以防止水分蒸发散失),冷却后,10000g,25℃离心5min,取上清。
定磷反应:
试剂名称标准管空白管对照管测定管标准溶液(μL)100---H
O(µL)-100--
2
上清液(µL)-100100
工作液(µL)900900900900混匀,37℃(哺乳动物)或25℃(其它物种)反应30min,冷却至室温,在660nm处,蒸馏水调零,记录各管吸光值A,分别记为A标准管、A空白管、A对照管和A测定管。
ΔA=A测定管-A对照管,ΔA标准=A标准管-A空白管。
标准管、空白管只要做一次即可,每个测定管需要设一个对照管。
三、ACC活性计算
1、标准曲线的绘制:
以各个标准溶液的浓度为x轴,其对应的ΔA标准为y轴,绘制标准曲线,得到标准方程y=kx+b,将ΔA带入方程得到x(μmol/mL)
2、ACC活性的计算:
(1)按蛋白浓度计算
酶活定义:每小时每毫克组织蛋白产生1μmol无机磷的量为一个ACC活力单位
ACC酶活(U/mg prot)=x×V总÷(V样×Cpr)÷T=20x÷Cpr。
(2)按样本质量计算
酶活定义:每小时每g组织产生1μmol无机磷的量为一个ACC活力单位。
ACC酶活(U/g鲜重)=x×V总÷(V样×W÷V样总)÷T=20x÷W。
(3)按照细菌或细胞数量计算
酶活定义:每小时每104个细菌或细胞产生1μmol无机磷的量为一个ACC活力单位。
ACC酶活(U/104cell)=x×V总÷(V样×细胞数量(万个)÷V样总)÷T
=20x÷细胞数量(万个)。
(4)按液体体积计算
酶活定义:每小时每mL液体产生1μmol无机磷的量为一个ACC活力单位。
ACC酶活(U/mL)=x×V总÷V样÷T=20x。
V总:酶促反应总体积,0.5mL;V样:加入样本体积,0.05mL;V样总:加入提取液体积,1mL;T:反应时间,0.5h;Cpr:样本蛋白质浓度,mg/mL,蛋白浓度自行测定;W:样本鲜重,g。
注意事项
1、工作液(定磷剂)应现配现用,正常颜色为浅黄色,如有变色或变蓝则均为失效。
2、此法具有微量、灵敏、快速的特点。
所以对测定所用试管或EP管等试验器材均要求严格无磷。
3、显色结束后应立即检测。
4、当ΔA大于1时,建议将样本用提取液稀释后再进行测定。