第八章分子结构一
第八章(3):大分子结构

大分子结构解析的难度
首先大分子(生物大分子,主要是蛋白 质)晶体一定没有中心中心对称,相位 不仅仅是正负号那么简单;
原子数目很多,帕特逊函数确定的原子 只能是很少很少的重原子(假如有的话, 很多情况下还没有重原子)。
实验数据误差很大。Fra bibliotek解决大分子结构相位问题的主 要方法
同晶差值傅立叶法 分子置换法 同晶置换法 反常衍射法
Isomorphous Difference Fourier Analysis:同晶差值傅 立叶法
已经知道晶体结构的大部分,需要解析余下 小部分的结构:如测定同晶型单点或多点突 变的蛋白结构(野生型的结构已知),确定 结合上小分子后的结构。
平移解
Sol_TF_1 1 30.56 145.90 162.75 0.600 0.156 0.401 88.37 0.554 0.444 Sol_TF_1 2 30.56 145.90 162.75 0.600 0.156 0.222 45.60 0.600 0.335 Sol_TF_1 3 30.56 145.90 162.75 0.599 0.156 0.201 44.52 0.601 0.332 Sol_TF_1 4 30.56 145.90 162.75 0.600 0.156 0.274 43.94 0.603 0.330 Sol_TF_1 5 30.56 145.90 162.75 0.600 0.156 0.263 42.48 0.605 0.327 Sol_TF_1 6 30.56 145.90 162.75 0.600 0.156 0.237 41.32 0.603 0.327 Sol_TF_1 7 30.56 145.90 162.75 0.600 0.156 0.390 40.43 0.607 0.326 Sol_TF_1 8 30.56 145.90 162.75 0.267 0.489 0.401 38.50 0.604 0.328 Sol_TF_1 9 30.56 145.90 162.75 0.600 0.656 0.401 27.25 0.618 0.296 Sol_TF_1 10 30.56 145.90 162.75 0.929 0.820 0.403 21.96 0.612 0.310
MHC分子讲义

组织相容性抗原
• 指供受者组织细胞表面的特异性抗原,是 决 定组织相容性的物质基础,也称移植抗原 (transplantation antigen)。
• 主要组织相容性抗原: 引起强而迅速排斥反应,在移植排斥反
应中起决定性作用。编码该抗原的基因称 主要组织相容性复合体。
主要组织相容性复合体
指某一染色体上的一群紧密连锁的基因群, 他们所编码的抗原决定了机体的组织相容性, 并与免疫应答和免疫调节有关。
HLA-DRB1*0901: 北方汉族人出现频率: 15.6% HLA-DQB1*0701: 北方汉族人出现频率: 21.9%
随机分配规律,几率: 15.6% x 21.9% =3.4%
实际检测出现的频率: 11.3%
三、HLA多态性的产生及意义
1、 产生: MHC基因突变和自然选择的结果
适者生存: 具较强抗病能力和较低死亡率的个 体的等位基因会有更多 的机会 将等位基因传递 给后代。
• 血清补体成分编码基因 • 抗原加工提呈相关基因 • 非经典的I类基因 • 炎症相关基因
血清补体成分编码基因
(经典的HLA-III类基因)
编码的产物均参与免疫应答和非特异性免疫调节 包括: A.补体成分编码基因:C2、C4A、C4B、BF
B.TNF基因家族(TNF) 热休克蛋白基因家族(HSP70)
识别、递呈原外源性抗原 受体:CD4分子 识别、调节Th细胞功能
二、HLA分子结构及其 与抗原肽的相互作用
• HLA分子结构 • HLA分子与抗原肽的相互作用 • HLA分子功能特点
HLA分子结构
HLA-I类分子: α链 ---- I类基因编码 β2-微球蛋白-第15对染色体基因编码 α1、α2 组成抗原结合槽
18 分子结构2

§3 共 价 键 的价 键 理 论
四、共价键的键型
第 八 章
共价键的键型有两种键,键
两个原子都含有成单的 s和px, py,pz电子,当它们沿x轴 接近时,能形成共价键的原子轨道有:s-s、px-s py-py px-px pz-pz
分 键: 沿键轴方向,以“头碰头”的方式发生轨道重叠, 子 轨道重叠部分是沿着键轴呈圆柱形的对称分布。 结 特点:键的键能大,稳定性高。 构
107°18′
101.9pm
N H H H
6
§1 化 学 键 参 数 四、键的极性
第 八 章 分
非极性键:在单质分子中两个原子之间形成的化 学键,由于原子核正电荷重心与负电 荷重心重合,叫做非极性键。
极性键:不同原子间形成的化学键,由于原子的电 子 负性不同,成键原子的电荷分布不对称,电 结 负性较大的原子带部分负电荷,电负性较 构 小的原子带部分正电荷,正负电荷重心不 重合,形成极性键。
第 八 章 分 子 结 构
16
§2 离 子 键
⑸、9~17电子构型:最外层为9~17个电子,ns2 np6 ndx(x=1~9) 如Cr3+、Fe2+
第 八 章 分 子 结 构
说明:
一般在离子的电荷和半径大致相同的条件下,不同构型 的正离子对同种负离子的结合力的大小可有如下经验规律: 8 电子构 型的离子
24
§3 共 价 键 的价 键 理 论
共价键的实质:成键原子的原子轨道发生了重叠。
第 八 章 分 子 结 构
二、价键理论
基本要点:
1、电子配对原理 两个原子接近时,自旋方向相反的成单电子可 以互相配对,形成共价键。
例如:
25
§3 共 价 键 的价 键 理 论
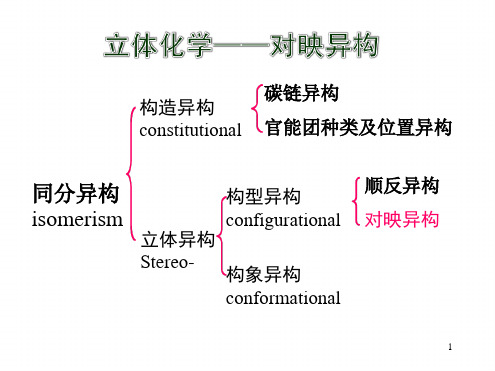
第八章 立体化学对映异构
? : 旋光度;c: 溶液浓度 (g/mL);ρ: 纯液体密度(g/mL) ;l: 管长(dm)
? ? 通常要标出测定时的温度和偏振光的波长:
?
t
?
溶剂对比旋光度也有影响,要注明所用溶剂。
例: 在20℃时,以钠光灯为光源测得葡萄糖水溶液的比旋光度为
右旋52.5°,记为:
??
?20 D
?
? 52.5?(水)
返回
26
R-S 标记法
? 根据手性碳原子所连接的四个基团在空间的排列来标记: (1)先把手性碳原子所连接的四个基团设为: a、b、c、d,并
将它们按次序规则排队。 (2)若a,b,c,d 四个基团的顺序是 a(较优基团)>b>c>d ,将该手
性碳原子在空间作如下安排:
? 把排在最后的基团 d放在离观察者 最远的位置,然后按 先后次序观察其他三个基团。 ? 即从最先的a开始看,经过b,再到c。 ? 若方向是 顺时针 的,则该手性碳原子的构型标记为 “R”;若为逆时针,则标记为“S”
-31.3°(乙酸乙酯 )
+31.3°(乙酸乙酯
外消旋体153
)
(III) (2R,3S)-(-) 167
-9.4°(水)
(IV) (2S,3R)- 167
+9.4°(水)
?非对映(+体) 可用一般的物理方法分离.
37
例:酒石酸的立体异构体
COOH
COOH
O HO C
HH *C C* OH OH
COOH
OH
乳酸(2-羟基丙酸)
H3C
C H
COOH
手性分子
乳酸的分子模型图
两个乳酸模型不能叠合
《医学遗传学》第八章 生化遗传病

三、珠蛋白生成障碍性贫血
珠蛋白生成障碍性贫血是由于珠蛋白多肽链完全不能合成或合成不足所致。成人血红蛋白(HbA)由两条α链和两条β链组成。珠蛋白生成障碍性贫血中凡由于α链合成减少或不能合成者称为α珠蛋白生成障碍性贫血,由于β链合成减少或不能合成者称为β珠蛋白生成障碍性贫血。
α或β珠蛋白链合成减少或甚至完全缺如涉及α或β珠蛋白基因的种种突变,可分为非缺失型(包括微缺失型)和缺失型两大类。非缺失型珠蛋白生成障碍性贫血涉及从5’转录控制信号,外显子密码,内含子(间隔顺序)拼接信号和共有序列(consensus sequence),外显子和内含子潜在的拼接部位,终止密码和3'多聚腺苷化信号等处的碱基取代、缺失、插入、移码等,导致转录受阻或转录产物异常,使RNA加工拼接或翻译受阻,RNA不稳定或翻译后异常肽链不稳定,最终导致患者α或β珠蛋白链减少(α+或β+珠蛋白生成障碍性贫血)或完全缺如(ao或β0珠蛋白生成障碍性贫血)。缺失型珠蛋白生成障碍性贫血涉及α或β珠蛋白基因簇较大范围的缺失,包括涉及α或β珠蛋白基因簇5'上游60kb α或β位点控制区缺失。大部全缺失发生在α或β位点控制区。
4.Hb Bristol不稳定血红蛋白病 本症亦为常染色体显性遗传,是由于β链第67位缬氨酸被天冬氨酸所取代,导致血红蛋白分子不稳定。这种不稳定的血红蛋白易在细胞中发生变性沉淀而形成Heinz小体,因而造成溶血性贫血。本症主要临床症状是先天性溶血性贫血,黄疸和脾肿大,因而亦称先天性Heinz小体溶血性贫血(congenital Heinz body hemolytic anemia)。已知的不稳定血红蛋白有90余种。
(一)α珠蛋白生成障蛋白基因主要突变类型和主要缺失类型见图7―9。
人体第16号染色体短臂上有2个连锁的α珠蛋白基因。如果16号染色体上的2个α基因均因突变或缺失而丧失了功能,我们把这种单倍型称为α地1(α-thal1)。如果16号染色体上的2个α基因只有1个因突变或缺失而丧失了功能,这种单倍型称为α地2(α-thal2)。α珠蛋白生成障碍性贫血杂合子也有一定的临床表现,故本症属常染色体显性遗传。因为患者丧失功能的α基因的数目不同,α珠蛋白生成障碍性贫血有以下4种临床类型(图7―10)。
第八章 醛和酮(一)醛和酮的命名(二)醛和酮的结构(三)醛和

N
六亚甲基四胺(乌咯托品)
(己)与Wittig试剂加成
Ph3P + CH3CH2Br C6H6
PhLi
Ph3PCH2CH3 Br
Ph3P=CHCH3 + C6H6+LiBr
Ph3P CHCH3
O CH3 C CH3 + Ph3P=CHCH3
O PPh3 CH3 C CHCH3
CH3
O PPh3 CH3 C CHCH3 0oC CH3 C CHCH3 + Ph3P O
C
O
92%
(4)羧酸衍生物的还原
COCl
LiAl(OBu-t)3H OCH3 乙醚,-78 oC
CH3
H+/H2O
CHO
OCH3 CH3
60%
CH3(CH2)10
COOC2H5
Al(Bu-n)2H 己烷,-78 oC
H+/H2O CH3(CH2)10 CHO 88%
(5)芳烃的氧化
V2O5 CH3 + O2(air) 350-360oC
CH CCOOH CH3
(丙)Mannich反应
O CCH3 HCHO
HN(CH3)2 HCl
O CCH2CH2N(CH3)2
H3C
CO +HCl AlCl3_ CuCl, 20oC
H3C
CHO
(四)醛和酮的物理性质
沸点:介于烃、醚与醇、酚之间。
CH3CH2CH2CH3 CH3OCH2CH3 CH3CH2CHO CH3COCH3 CH3CH2CH2OH
沸点/ oC -0.5
8
49
56
97
CH2CH3
CHO
第八章 核酸的结构

第八章核酸的结构主要内容一.核苷酸二.核酸的共价结构三.DNA的高级结构四.RNA的高级结构核糖常见的核苷酸及其缩写符号多磷酸核苷酸3`,5`-环化腺苷酸z5′-磷酸端(常用5′-P表示);3′-羟基端(常用3′-OH表示)z多聚核苷酸链具有方向性,当表示一个多聚核苷酸链时,必须注明它的方向是5′→3′或是3′→5′。
5′P dA P dC P dG P dT OH3′5′P A P C P G P U OH3′或5′ACGTGCGT 3′5′ACGUAUGU 3′DNA RNA2.DNA的一级结构DNA的一级结构是由数量巨大的四种核苷酸连接起来的直线或环线多聚体。
包含着生物体的遗传信息。
基因组计划人类基因组中仅有1.1-1.4%编码蛋白质。
2.RNA的一级结构RNA的种类多,结构也不一样。
tRNA含有较多的稀有碱基,3′-端为CCA,5′-端多数为pG,也有的为pC。
一级结构中有一些保守序列,与其特殊结构与功能有关。
真核生物rRNA的甲基化修饰核苷比原核生物多。
原核生物的mRNA是多顺反子mRNA:一条mRNA上有多个编码区、5′-端、3′-端和各编码区间的非编码区。
真核生物的mRNA为单顺反子,5′-端有帽子结构,3′-端有poly(A)尾巴。
帽子结构有助于核糖体对mRNA的识别和结合,使翻译正确起始。
尾巴结构与mRNA的运输与寿命有关。
1953年Watson和Crick提出了著名的DNA双螺旋结构模型。
1962年,沃森与克里克,偕同威尔金斯共享诺贝尔生理或医学奖。
1976年沃森担任美国冷泉港实验室主任。
沃森使冷泉港实验室成为世界上最好的实验室之一。
他还是人类基因组计划的倡导者,1988年至1993年曾担任人类基因组计划的主持人。
DNA双螺旋内容:的多核苷酸链围绕同一中心轴盘绕而成的右手双螺旋(2)碱基处于螺旋内侧,而磷酸及戊糖位于外侧。
碱基的平面与螺旋轴相垂直,糖平面与碱基平面几乎成直角。
第8章 分子的拆分

H O O
H
H
H
H
2
H CO2Me FGI CO2Me
CO 2Me CO Me
FGI HO HO H
O O H HH H
H
+
CO2Me
CO2Me
H CO2Me OsO
4
H KMnO4 HO HO CO2Me
O
TM
+ H+
H
H
3. β-羟基羰基化合物 β-羟基羰基化合物可用羟醛缩合反应来制备。
OH O O H + O
1)
LiAlH4 Ac2O, R2NH NH3(l)
2) 3)
TM
Et H
6. 利用分子的对称性
eg. 设 计
eg. 设 计 分 析 TM
eg. 设 计
HO
Et H HEt
Et H H Et HCH Et Cl
合 成 路 线 OH 的
合 成 路 线 OH 的
合 成 路 线 OH 的
HO
HO
HO
HO
O O O O O O OEt OEt , O CN ,
1. 相同酯间的缩合
O O CH3C CH2C OC2H5 O CH3C OCH2CH3
δ +
O
+
CH2
C OCH2CH3
2.酯分子内缩合---Dieckmann 酯缩合
合 成 2 CH 3O CH 3O
Fe CH CH CH 3 茴 香 脑 ( 大 豆 茴 香 油 为 原 料 ) 2 CH 3O CH Et Fe HCl △ 2 CH 3O CH Et CH CH CH 3 △ Cl CH CH CH 3 茴 香 脑 ( 大 豆 茴 香 油 为 原 料 ) Cl 2 CH Fe O CH Et 3 △ Cl CH CH 2 CH 3O Et Et OCH 3 HI TM CH CH
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
如 Fe2+,Mn2+ 等(称为不饱和电子构型)
Na ( s ) Cl2 ( g ) NaCl ( s ) H 410.9kJ mol1 2
10
(二) 离子键的本质和特点 无方向性:离子是带电体,电荷分布是球形对称, 对各个方向的吸引力是一样的。 ●Na+ 无饱和性:空间许可的话,一个离子可以同时和几 ●Cl个电荷相反的离子相吸引。
荷越高,半径越小。如 S2- > S; Fe3+ < Fe2+ < Fe
5) 一般,正离子半径较小,约在10~170pm之间;
负离子半径较大,约在130~260pm之间。
如第二周期 F- (133pm),而Li+(76pm); 第四周期 Br-(196pm),而 K+(138pm) 。 虽然差了两个周期,但 F- 与K+半径接近。
27
§2 共价键
1916年美国化学家路易斯(G. N. Lewis)为了说 明分子的形成,提出经典的共价键理论,成功地解 释了同原子组成的分子(H2, O2, N2等)及性质相近的 不同原子组成的分子(HCl, H2O等)的形成,初步揭 示了共价键和离子键的区别。 但它不能解释某些分子的中心原子最外层电子数多 于或少于8时,分子仍能稳定存在(BF3, PCl5),以及 共价键的特性(方向性、饱和性),单电子键的存在
17
(三) 离子半径(ionic radius)
离子半径:根据晶体中相邻正负离子的核间距 d
测出,并假设 d = r+ + r1926年, 哥德希密特(Goldschmidt)用光学折射方法测
定, 得到了F- (133pm)和 O2-(132pm) 的半径, 并以此
为基准,结合X射线衍射数据, 得到一系列离子半径。
18
Mg2+的半径 rMg2+ = 210 - 132 = 78pm 1960年, Pauling 以最外层电子到核的距离定义为离 子半径,并利用有效核电荷等关系,用其它方法推算
出rO2-=140pm,以此为基础得到其它离子的半径,求
出一套离子半径数据,称为 Pauling半径。(表8-2)
19
28
(H2+),O2的磁性等。尤其不能阐明为何“共用电子”
基本要求
1. 熟悉离子键的形成与特征。 2. 熟悉现代价键理论。掌握共价键的特征、键型、键的 极性和分子的极性。 3. 熟悉杂化原子轨道,了解其应用。掌握sp型杂化轨道 的类型及空间分布图形。 4. 熟悉分子轨道理论,了解与现代价键理论的区别。掌 握第二周期同核双原子分子的分子轨道能级和电子排 布。了解异核双原子分子的分子轨道组成及大 键。 5. 掌握价层电子对互斥模型。能够利用该模型推测主族 元ABm型分子或离子的空间构型。
23
1)极化力:指离子产生电场强度的大小。
①正离子半径越小,电荷越多,极化能力越强;
Li+ > Na+ > K+ > Rb+ > Cs+
②对不同电子层结构的正离子来说,在半径和电荷
相近时,其极化能力(18或18+2电子构型及He型
离子,如Ag+, Pb2+, Li+) > (9~17电子构型的离子,
如Fe2+, Ni2+, Cr3+) > (8电子层Na+, Mg2+, Ca2+)
③复杂负离子的极化作用通常较小,但 SO42- 和 PO43- 有一定的极化作用。
24
2)变形性:离子在电场作用下电子云发生变形的难易。 ①离子半径越大,变形性越大: Li+ < Na+ < K+ < Rb+ < Cs+; F- < Cl- < Br- < I-
16
(二) 离子的电子构型(electronic configuration of ions) 离子的电子构型就是指离子的电子层结构。
简单负离子的外电子层都是稳定的稀有气体结构的8电子构型, 而正离子电子构型包括: ①2电子构型—离子最外层电子是s2,如 Li+, Be2+ 等
②8电子构型—离子最外层电子是s2p6, 如 Na+, Al3+ 等
13
Born和Haber设计了一个热力学循环过程, 从已知的 热力学数据出发, 计算晶格能。
1 M (s) X 2 ( g ) 2
S 1 D 2
f Hm
MX ( s )
U
M (g) X (g)
S:固态金属M的升华热 D:气体X2的解离能 I: 气态金属M的电离能
I ( A)
离子周围排列电荷相反离子的数目,主
要取决于正负离子的半径比 (r+/r-):比
值越大,周围排列的相反离子的数目越多。 NaCl
作用力实际是静电吸引力。
11
二、晶格能-离子键的强度
离子键的本质是正负离子间的静电吸引力,其 强度与离子的电荷成正比,与离子间的距离的成反 比。
Z Z U r r
离子半径变化的规律: 1) 同周期: 主族元素:从左至右正离子电荷数升高,最高价 离子半径最小,如 Na+ > Mg2+ > Al3+ ; 而负离子半径随电荷增加而增大,如 F- (133pm)< O2-(140pm)。
过渡元素:离子半径变化规律不明显。
2) 同主族:从上到下电子层增加,具有相同电荷数
6
形成过程: 以 NaCl 为例
1) 电子转移形成离子,分别达到 Ne 和 Ar 的稀有气体 原子的结构,形成稳定离子。 Na – e —— Na+ Cl + e —— Cl-
相应电子构型变化 2s22p63s1 2s22p6, 3s23p5 3s23p6
7
2)靠静电吸引, 吸引力和排斥力达到平衡时形成化学键,体 系的势能与核间距之间的关系如图所示: r > r0, 当 r 减小时, 正负离子 体系的势能 V
8
离子键的形成条件
1) 元素的电负性差要比较大
Δχ > 1.7, 发生电子转移, 形成离子键; Δχ < 1.7, 不发生电子转移, 形成共价键。
但离子键和共价键之间, 并非严格截然可以区分的。化
合物中不存在百分之百的离子键, 即使是 NaF 的化学键之
中, 也有共价键的成分, 即除离子间靠静电相互吸引外,尚
21
6) 镧系和锕系收缩:像原子半径一样,相同正价的镧系
和锕系阳离子的半径随原子序数的增加而减小。
同一副族内的元素,离子半径没有简单的变化规律。
一般来讲,离子半径越小,离子间吸引力越大,相应
化合物的熔点就越高。如:
离子化合物
熔点/oC 正离子半径/pm
LiF
1040 60
NaF
995 95
KF
856 133
②负离子电荷越多,变形性越大;正离子电荷越高,
变形性越小: O2- > F- > Na+ > Mg2+
③18电子层和不规则电子层的阳离子,其变形性比
相近半径的稀有气体型离子大很多:18电子构型、
9~17电子构型 > 8电子构型。
如 Ag+ > K+, Hg2+ > Ca2+
25
④一些阴离子的变形性: ClO4- < F- < NO3- < OH- < CN- < Cl- < Br- < I3) 相互极化作用(附加极化)
通常正离子半径小,负离子半径大,所以正离子
极化力大,变形性小;而负离子极化力小,变形 性大。因此,一般只考虑负离子在正离子的电场 下发生的变形,即正离子使负离子极化。
当阳离子也容易变形时(Hg2+, Ag+, Pb2+),往往
会引起两种离子之间相互的附加极化效应:
26
即正离子也可被负离子极化,极化后的正离子反过 来又增强了对负离子的极化作用。从而加大了离子 间的引力。 随着极化作用的增强,负离子电子云明显地向 正离子方向移动,使原子轨道重叠的部分增加,即 离子键向共价键过渡。
1
6. 熟悉范德华力的产生及氢键的形成。掌握分 子间力对物质物理性能的影响。
7. 了解离子极化及其对键型和物质某些性质的 影响。
8. 熟悉金属晶体、离子晶体、原子晶体、分子 晶体四种晶体类型和相关的物理性质。 9. 了解金属晶体的紧密堆积方式。
2
分子是物质能独立存在并保持其化学特性的最
小微粒。物质的化学性质主要决定于分子的性质,
的离子半径增加。
Li+ < Na+ < K+ < Rb+< Cs+ ; F- <Cl- <Br- <I20
3) 周期表相邻两主族对角线上,左上元素和右下元素
的离子半径相似:如 Li+(76pm) 和 Mg2+(72pm),
Na+(102pm) 和 Ca2+(100pm)
4) 同一元素正离子的电荷数增加则半径减小,且正电
靠静电相互吸引, V减
小, 体系稳定。 r = r0, V有极小值, 此时体系 最稳定,表明形成了离 子键。 核间距r