植物生理学:第三节 植物对矿质元素的吸收
植物生理学-03-矿质营养

0
为膜原有电化学势;
j
aj为离子的电化学活度;Zj为离子的代数化合价 F为法拉第常数,96.5J/mol;E为电势
膜内外离子 j 分布产生的电化学势分别表示为:
µ µ0
=
j外
j + RT ln a j外 + Z j ⋅ F ⋅ E外 (2)
µ µ0
=
j内
j + RT ln a j内 + Z j ⋅ F ⋅ E内 (3)
What was wrong with him?
矿质营养学说与农业化学的建立
李比希(J. Liebig)1840年伦敦有机化学年会上发表 了“化学在农业和生理学上的应用”的论文,否定了 腐殖质营养学说,提出了矿质营养学说。
腐殖质(humus)是有了植物后才出现在地球上的而不是 植物出现以前。因此土壤中矿物质是一切绿色植物的 唯一养料,厩肥及其它有机肥料对于植物所起的作用, 并不是由于其中所含的有机质,而是由于这些有机质 在分解时所形成的矿物质。
平衡溶液:将某种植物所必需的矿质元素按照一定浓度和适当比例 配制成的,并对该植物生长发育具有良好作用而无毒害的混合溶液, 称为该种植物的平衡溶液。
二、植物细胞吸收矿质元素的机理
两种吸收机理:主动吸收和被动吸收
离子过膜的驱动力: 1、化学势梯度(浓度差):由高到低 2、电势梯度:阳离子被负电荷吸引;阴离子被正电荷 吸引。
植物的必需元素(essential element) 大量元素:C、H、O、N、P、K、S、Ca、Mg、 微量元素:Fe、Mn、Cu、Mo、Zn、B、Cl、Ni、Na
必需的矿质元素:N、P、K、S、Ca、Mg、Fe、Mn、 Cu、Mo、Zn、B、Cl、Ni、Na
植物生理学:第三节 植物对矿质元素的吸收

酸性时,利于难溶金属离子的吸收, 但一些离子(P、K、Ca、Mg等) 易流失; 碱性时,一些金属离子形成难溶盐 See your Text Book
3 菌根 • 非病原性或弱病原性真菌与根的活细胞间形成的表皮或皮层细胞间隙, • 自然界中,97%的植物的根与真菌共生成菌根 • 菌根增加了植物对矿质元素的吸收
• 相互依赖:
矿质须在溶液状态才被吸收,矿质随水分一起进入根部 的质外体中; 根系对矿质的主动吸收使根部的水势降低,有利于水分 进入根部
• 相互独立:
吸收矿质和水分的机理不同; 吸收矿质以耗能的主动吸收为主; 而水分则按水势高低进行被动的运输
2 根系对矿质的吸收具有选择性
• 细胞中不同溶质的载体数量及活性均不同
第三节 植物对矿质元素的吸收
• 通过地上部吸收 • 通过根系吸收
主要途径
• 研究(大麦根)离子吸收 的模式
Plants develop extensive root systems
Wheat
Fibrous Root System
Tap Root System
一、根系吸收溶质的特点
1 根系对矿质与水分吸收的相对性
Total length of roots (m m)
59 70 254 324
二、根系吸收溶质的部位
• 根毛区:表面积大,输 导组织发育完全,易吸 收并转运
• 分生组织:积累大量离 子,因无输导组织,所 吸收的离子不易被转运 出去
甜玉米幼苗的主根
三、根系吸收溶质的过程
• 通过与H+和HCO3-交换吸附方式吸附在根部细胞表面 • 进入根的内部:
• 阴离子被土壤颗粒排斥,存在于土壤溶液中;易被水 淋洗而流失;
02-植物吸收矿质元素机理

(二)杜南平衡
当细胞内某些离子的浓度已经超过外界溶液该离子的浓度时,外界的 离子仍然向细胞移动的现象。
杜南平衡是科学家杜南(F.G.Donnan)提出的一种说明离子积累现象 的特殊平衡。其基本观点是:
3.5 植物吸收矿质元素的机理
植物细胞吸收矿质元素的方式有三种:主动吸收、被动 吸收和胞饮作用。其中以前两种方式较为普遍。
主动吸收:
需要利用代谢提供的能量, 逆电化学势梯度吸收物质的 过 程。故又称代谢性吸收。
被动吸收:
不需要代谢提供能量,顺电化学势梯度吸收物质的过程 。 故又被称为非代谢性吸收。
上述学说是高度假定的,但它概括了研究离子吸收的主要实验事实、 含义和结论:
1.能量驱动的离子吸收是有选择性的; 2.能量可能成ATP形式; 3.离子运输可以逆浓度梯度发生; 4.离子吸收与酶促过程有关;
其实验证据:
遇到的困饱和动力学 2. 离子竞争现象 3. 解偶联剂抑制离子的吸收 4. 人工模拟及现代膜结构理论
活化载体
离子 载体-离子
复合物
P AC
磷酸 激酶
IC CIC P
ATP
ADP
线 粒 体
复合体运转至膜内侧,在磷酸 酯酶作用下释放出的磷酸基, 未活化载体
磷酸
酯酶 Pi
使载体失去对离子的亲和力,
从而将离子释放到膜内。
外
内
膜
扩散方式
A
i
i
i
C
i
能量
B
i
i
能量
D
i
i
矿质营养植物对矿质元素的吸收(共63张PPT)

离子通道运输:ion channel transport
一个开放
的离子通
道每秒可
运输107 –
108个离子, 比载体蛋
白的运输
快1000倍。
②载体蛋白:又称载体、透过酶、运输酶,包括单向运
输载体、同向运输载体、反向运输载体√
细胞质膜上的蛋白,可选择性地与质膜一侧的分子 或离子结合,形成载体-离子复合物,通过载体蛋 白构象的变化,把分子或离子运送到质膜的另一侧。
(1) 溶液培养法(水培法): 在含有全部或部分营养元素的溶液中栽培植物 的方法。
(2) 砂基培养法(砂培法): 在洗净的石英砂或玻璃球中,加入含有全部 或部分营养元素的溶液来栽培植物的方法。
20世纪70年代后还发展了气培法和营养膜法
2.植物必需元素的种类√ 根据试验结果,按照上述原则,确定植物的 必需元素有17种。
如:海洋植物+纯NaCl 不久就死去 海生植物+海水(NaCl含量很高) 生活的很好
离子拮抗:√
细胞通过膜的内折
从外界直接摄取物质进
入细胞的过程。
细胞内吞作用
是非选择性吸收,在吸收水的同时,把水分中的 物质如各种盐类和大分子物质甚至病毒一起吸收进来。
番茄和南瓜的花粉母细胞、蓖麻和松的根尖细胞
都有胞饮现象。
第三节 植物对矿质元素的吸收
根系吸收矿质元素的特点 根系吸收矿质元素的部位 根系吸收矿质元素的过程 影响根系吸收矿质营养的土壤因素 植物地上部分对矿质元素的吸收
积累现象
活细胞从周围环境吸收必需元素,最终使其在细胞内的 浓度远高于细胞外的浓度,这种现象称为积累。
内部浓度与外部浓度之比称为积累比。
如通常植株中K+浓度约为25mmol/L-1,而土壤中溶解
第三节 植物对矿质元素的吸收及运输
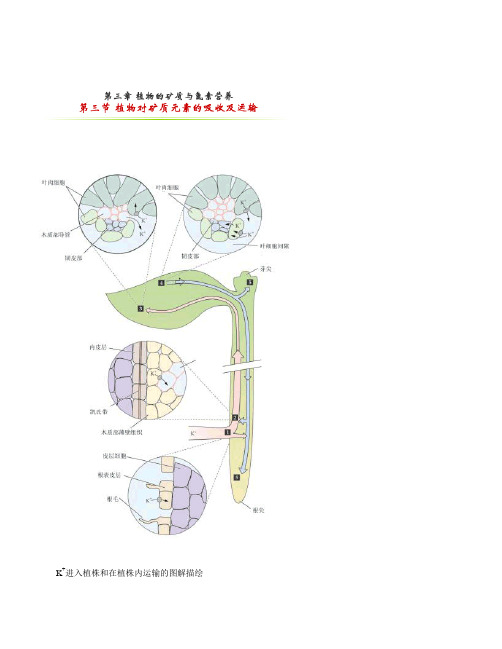
K+进入植株和在植株内运输的图解描绘K+在木质部内的运输(红箭头)和在韧皮部中的运输(蓝箭头)。
数字代表K+长距离运输途径中重要的运输位点。
5个中的4个数字标示的位置,夸大描绘了K+在细胞水平的运输(1)K+被跨根细胞质膜吸收(纵切观察)。
(2)K+通过跨木质部细胞膜输出。
运输死的厚壁木质部导管(横切面观察)。
—、植物吸收矿质元素的特点(一)根系吸收矿质与吸收水分不成比例矿质元素必须溶解在水中,才能被植物吸收。
过去认为植物吸收矿质是被水分带入植物体的。
按照这种见解,水分和盐分进入植物体的数量,应该是成正比例的。
但后来的大量研究证明,植物吸水和吸收盐分的数量会因植物和环境条件的不同而变化很大。
有人用大麦作试验,通过光照来控制蒸腾,然后测定溶液中矿质元素的变化。
结果发现,光下比暗中的蒸腾失水大2.5倍左右,但矿质吸收并不与水分吸收成比例(表3-6)。
如磷酸根和钾离子在光下比暗中的吸收速度快,而其它无机盐,如Ca、Mg、SO2-4、NO-3等,在光下反而吸收少。
总之,植物对水分和矿质的吸收是既相互关联,又相互独立。
前者,表现为盐分一定要溶于水中,才能被根系吸收,并随水流进入根部的质外体。
而矿质的吸收,降低了细胞的渗透勢,促进了植物的吸水。
后者,表现在两者的吸收比例不同,吸收机理不同:水分吸收主要是以蒸腾作用引起的被动吸水为主,而矿质吸收则是以消耗代谢能的主动吸收为主。
另外两者的分配方向不同,水分主要分配到叶片,而矿质主要分配到当时的生长中心。
(二)根系对离子吸收具有选择性离子的选择吸收(selective absorption)是指植物对同一溶液中不同离子或同一盐的阳离子和阴离子吸收的比例不同的现象。
例如供给NaNO3,植物对其阴离子(NO-3)的吸收大于阳离子(Na+)。
由于植物细胞内总的正负电荷数必须保持平衡,因此就必须有OH-或HCO3-排出细胞。
植物在选择性吸收NO-3时,环境中会积累Na+,同时也积累了OH-或HCO-3,从而使介质pH值升高。
植物生理学03矿质营养

图3-5 植株缺磷症状
4、钾(K)
吸收形式:K+。 作用:
是酶的活化剂。 促进碳水化合物的合成和运输。
增加原生质的水合程度,提高细胞的保水能力,增强 抗旱性。 缺素症:叶色缺绿变黄,逐渐坏死。茎秆柔弱
易倒伏(图3-6、3-7)。 K的移动性大,缺乏时,症状在老叶出现。
图3-6 番茄缺钾初期叶缘失绿
图3-4 缺硫植株中上部叶色淡
3、磷(P)
吸收形式:H2PO4-。 作用:
磷是核酸、磷脂、辅酶和ATP的组成成分; 磷在碳水化合物代谢中起着重要作用; 磷对氮代谢也有影响。 缺素症:缺素症与N相似,生长缓慢,植株矮小, 叶片暗绿,有些植物呈紫色或红色。 P的移动性大,缺乏时,症状首先在老叶出现(图35)。
图3-9 缺钙幼叶叶尖变褐枯死
6、镁(Mg)
吸收形式:Mg2+。 作用:
是叶绿素的组成成分。 是酶的活化剂。
缺素症:缺绿病,严重时形成褐斑坏死。 Mg的移动性大,缺乏时,症状首先 在老叶出现(图3-10)。
图3-10 植株缺镁症状
7、铁(Fe)
作用: 是酶的辅助因子。 是叶绿素合成过程所必需的。
岩石圈的元素组成包括O、Si、Al、Fe、Ca、 Na、K、Mg等,其中O和Si的含量最丰富,约占岩 石圈元素总量的75%。
二、植物体内的元素
植物体
干物质(5-90%) 有机物(90%)
水分(10-95%) 无机物(10%)
105℃ 烘干
植物
干物质
600℃ 灰分
构成灰分的元素称为灰分元素(灰分中的元素直 接或间接地来自土壤矿质,故又称矿质元素)。
必需元素是指维持植物正常生理活动所必需的元素。
判断必需矿质元素的原则:
植物对矿质元素的吸收
• 影响根系吸收矿质元素的因素(条件)
植物地上部分对矿质元素的吸收 矿质元素在植物体内的利用
一、根系对矿质元素的吸收
(一)根系吸收的主要区域
植物吸收的部位:根、茎、叶、果 根系是陆生植物吸收水和无机盐的主要器官 陆生植物有庞大的根系
吸收矿质元素的主要部位:根毛区
一、根系对矿质元素的吸收
离子在根中经质外体途径和共质体途径到达输导组织的模型
①共质体运输;②质外体运输;→表示主动运输,
表示再吸收
二、植物吸收矿质元素的特点
(一)根系吸收矿质与吸收水分的相对性 1. 相互关联
矿质需要溶解于水才能被根系吸收 2. 相互独立
两者的吸收机理不同,吸水是因蒸腾拉力引 起的被动过程;而吸盐是消耗代谢能量的主动吸 收过程,有饱和效应。
(一)根系吸收的主要区域
植物吸收的部位:根、茎、叶、果 根系是陆生植物吸收水和无机盐的
主要器官 陆生植物有庞大的根系
吸收矿质元素的主要部位:根毛区
(二)根系吸收养分的主要形态
气态:CO2、O2、SO2、水汽等 气气态态::CCOO2、2通、OO过22、、扩SSOO22、散、水水进汽汽入等等,植,通通物过过扩体扩散或进散入进有植入叶物植体片物内的体或内由气或叶孔片由直的叶气片接孔的进直气接入孔进直入接植进
开始随浓度的提
25
K 吸收速度(μmol/g.h)
高而迅速增加,
20
然后缓慢增加,
以后稳定在一定
15
的速率。如果继
10
续提高养分浓度
5
,养分吸收的速
率会出现“迅速 增加—缓慢增加
0.10
0.20 10
25
50
植物生理学-大纲
第一节植物对水分的需要第二节植物细胞对水分的吸收第二章植物的矿质营养第二节植物细胞对矿质元素的吸收第三节植物体对矿质元素的吸收第四节无机养料的同化第五节矿质元素在植物体内的运输第六节合理施肥的生理基础第三章植物的光合作用第一节光合作用的重要性第二节叶绿体及叶绿体色素第三节光合作用的机理第四节光呼吸第五节光合作用的进化第六节影响光合作用的因素第七节植物对光能的利用第四章植物的呼吸作用第一节呼吸作用的概念及其生理意义第二节植物的呼吸代谢途径第三节生物氧化第四节呼吸过程中能量的贮存和利用第五节呼吸作用的调节和控制第六节影响呼吸作用的因素第七节呼吸作用与农业生产第五章植物体内有机物质的运输第一节植物体内的信息传递第二节高等植物的运输第三节有机物运输的机理第四节同化物的分配及影响因素第六章植物生长物质第一节生长素类第二节赤霉素类第三节细胞分裂素类第四节脱落酸第五节乙烯第六节油菜素内酯第七节生长抑制物质第八节多胺和钙调素第七章光形态建成第一节光敏色素的发现和分布第二节光敏色素的化学性质及光化学转换第三节光敏色素的生理作用第四节光敏色素的作用机理第五节蓝光反应第八章植物的生长生理第一节种子的萌发第二节细胞的生长和分化(略讲)第三节植物的生长第四节植物的运动第九章植物的生殖生理第一节幼年期第二节春化作用第三节光周期第四节花器官形成的生理第五节受精生理第十章植物的成熟和衰老生理第一节种子成熟时的生理生化变化第二节果实成熟时的生理生化变化第三节种子和延存器官的休眠第四节植物的衰老第五节植物器官的脱落。
植物生理学2矿质营养
第二节 植物细胞对矿质元素的吸收※
1、离子的选择性积累
玉米根对离子的选择性吸收
离子 胞外浓度 胞内浓度 积累率
(mmol/L) (mmol/L) (Ci/Co)
K+ 0.14
160 1142
Na+ 0.51
0.6 1.18
NO3- 0.13 SO42- 0.61
38
292
14
23
Байду номын сангаас
第二节 植物细胞对矿质元素的吸收※
向通道 ➢ 应用膜-片钳技术,在质膜、液泡膜上已发现有K+、
Cl-、Ca2+等离子通道 • 离子带电性和水合规模影响其通透性; • 离子通道可由电势或外界刺激(光照、激素、化
学物质等)激活。
社引膜
:
,2009.
自片
王钳
.
忠 主 编 《 植 物
技 术 测 定 离
生子
理通
学道
.
》原
中 国 农
理 图
业
出
第一节 植物必需的矿质营养
(一)植物必需元素的标准※(Arnon等,1939) 1、不可缺少性:若缺乏,植物不能完成其生活史; 2、不可替代性:若缺乏,植物表现专一的缺素症; 3、直接功能性:
第一节 植物必需的矿质营养
现已证实植物的必需元素有17种, 必需矿质元素 有14种。
大量元素(一般,≥0.1%植物干重)9种 : C、H、O;N、P、K;Ca、Mg、S;
缺Cu:叶片生长缓慢,幼叶呈蓝绿色,出现枯斑, 栅栏组织退化,易过度蒸腾而萎蔫
Mo
吸收形式: MoO42作用: • 硝酸还原酶(NR)、固氮酶(钼铁蛋白)的组
成成分; • 黄嘌呤脱氢酶及脱落酸合成中的某些氧化酶的
第三节 植物对矿质元素的吸收及运输
K+进入植株和在植株内运输的图解描绘K+在木质部内的运输(红箭头)和在韧皮部中的运输(蓝箭头)。
数字代表K+长距离运输途径中重要的运输位点。
5个中的4个数字标示的位置,夸大描绘了K+在细胞水平的运输(1)K+被跨根细胞质膜吸收(纵切观察)。
(2)K+通过跨木质部细胞膜输出。
运输死的厚壁木质部导管(横切面观察)。
—、植物吸收矿质元素的特点(一)根系吸收矿质与吸收水分不成比例矿质元素必须溶解在水中,才能被植物吸收。
过去认为植物吸收矿质是被水分带入植物体的。
按照这种见解,水分和盐分进入植物体的数量,应该是成正比例的。
但后来的大量研究证明,植物吸水和吸收盐分的数量会因植物和环境条件的不同而变化很大。
有人用大麦作试验,通过光照来控制蒸腾,然后测定溶液中矿质元素的变化。
结果发现,光下比暗中的蒸腾失水大2.5倍左右,但矿质吸收并不与水分吸收成比例(表3-6)。
如磷酸根和钾离子在光下比暗中的吸收速度快,而其它无机盐,如Ca、Mg、SO2-4、NO-3等,在光下反而吸收少。
总之,植物对水分和矿质的吸收是既相互关联,又相互独立。
前者,表现为盐分一定要溶于水中,才能被根系吸收,并随水流进入根部的质外体。
而矿质的吸收,降低了细胞的渗透勢,促进了植物的吸水。
后者,表现在两者的吸收比例不同,吸收机理不同:水分吸收主要是以蒸腾作用引起的被动吸水为主,而矿质吸收则是以消耗代谢能的主动吸收为主。
另外两者的分配方向不同,水分主要分配到叶片,而矿质主要分配到当时的生长中心。
(二)根系对离子吸收具有选择性离子的选择吸收(selective absorption)是指植物对同一溶液中不同离子或同一盐的阳离子和阴离子吸收的比例不同的现象。
例如供给NaNO3,植物对其阴离子(NO-3)的吸收大于阳离子(Na+)。
由于植物细胞内总的正负电荷数必须保持平衡,因此就必须有OH-或HCO3-排出细胞。
植物在选择性吸收NO-3时,环境中会积累Na+,同时也积累了OH-或HCO-3,从而使介质pH值升高。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
• 通过地上部吸收 • 通过根系吸收
主要途径
• 研究(大麦根)离子吸收 的模式
Plants develop extensive root systems
Wheat
Fibrous Root System
Tap Root System
一、根系吸收溶质的特点
1 根系对矿质与水分吸收的相对性
• 表现出对同一种盐的正、负离子的吸收速率不同, 产生: 生理酸性盐:如铵盐[(NH4)2SO4],对NH4+的吸 收多于对阴离子的吸收; 生理碱性盐:如硝酸盐(NaNO3),对NO3-的吸收 多于对阳离子的吸收; 生理中性盐:如NH4NO3,对阴、阳离子的吸收速 率相同。
3 根系对矿质的吸收表现出单盐毒害和离子拮抗
Total length of roots (m m)
59 70 254 324
二、根系吸收溶质的部位
• 根毛区:表面积大,输 导组织发育完全,易吸 收并转运
• 分生组织:积累大量离 子,因无输导组织,所 吸收的离子不易被转运 出去
甜玉米幼苗的主根
三、根系吸收溶质的过程
• 通过与H+和HCO3-交换吸附方式吸附在根部细胞表面 • 进入根的内部:
• 影响矿物质的可与程度:
酸性时,利于难溶金属离子的吸收, 但一些离子(P、K、Ca、Mg等) 易流失; 碱性时,一些金属离子形成难溶盐 See your Text Book
3 菌根 • 非病原性或弱病原性真菌与根的活细胞间形成的
一种互惠共生体
• 真菌的菌丝体一部分侵入根的表皮或皮层细胞间隙, • 自然界中,97%的植物的根与真菌共生成菌根 • 菌根增加了植物对矿质元素的吸收
• 红松的菌根
• 菌根对玉米生长的影响
左:有菌根 右:无菌根
• 菌根增加了植 物的吸收范围
4 土壤中矿质元素的量 营养耗尽区
5 温度 通过呼吸作用影响主动运输
6 通气 O2与CO2交换能力
7 地上部生长情况 提供呼吸所需的碳水化合物
六、地上部对矿质元素的吸收
• 主要从叶片吸收: 叶面营养
• 矿质元素主要由角 质层进入细胞内部
• 吸收难溶盐:
根系分泌CO2、柠檬酸及苹果酸等有机酸,溶解某些难溶矿 物质
• 生长在岩石。
五、影响根系吸收矿质元素的因素
1 土壤结构和矿质元素
• 土壤胶体颗粒表面带“-”电荷,故吸附多种阳离子。 加之土壤颗粒的“表面积/体积”极大,成为一营养库;
• 单盐毒害:培养液中 只有一种金属离子而 对植物起毒害作用, 即使这种元素是植物 所必需的
• 离子拮抗:在单盐培 养液中加入少量的含 其他金属离子的盐, 就能减弱或消除单盐 毒害
小麦根在不同溶液中的生长情况
Solution
NaCl CaCl2 NaCl + CaCl2 NaCl + CaCl2 + KCl
• 阴离子被土壤颗粒排斥,存在于土壤溶液中;易被水 淋洗而流失;
• 土壤的通气状况(气相)影响O2和CO2的交换
• 土壤中的 离子交换
2 土壤pH值
• pH直接影响根系生长:
多数植物适宜微酸性(pH5.5-6.5)土壤; 少数植物适宜微碱性土壤(甘蔗、甜菜、石榴等)
• 影响土壤微生物生长:
微酸性时,真菌生长良好; 酸度高时,根瘤菌死亡; 较碱性时,有害微生物(反消化细菌)生长良好
• 相互依赖:
矿质须在溶液状态才被吸收,矿质随水分一起进入根部 的质外体中; 根系对矿质的主动吸收使根部的水势降低,有利于水分 进入根部
• 相互独立:
吸收矿质和水分的机理不同; 吸收矿质以耗能的主动吸收为主; 而水分则按水势高低进行被动的运输
2 根系对矿质的吸收具有选择性
• 细胞中不同溶质的载体数量及活性均不同
(1〕溶质扩散至质外体,即表观自由空间 (2)由于内皮层上凯氏带的阻隔,溶质进入共质体 • 进入导管,随木质部汁液向地上部输送
• 离子进入根部 的径向途径
四、根系对非溶解态矿质元素的吸收
• 吸收土壤胶体上吸附的矿质元素:
1 离子交换:根释放CO2,形成H2CO3,其中的H+与土壤表面 的阳离子交换; 2 接触交换:土壤颗粒上的阳离子与根表面的H+ 通过离子 振动直接交换